
Abstract
Sustainable agriculture in microgravity is integral to future long-term human space exploration. To ensure the efficient and sustainable cultivation of plants in space, a contingency plan to monitor plant health and mitigate plant diseases is necessary. Yet, neither methods nor tools currently exist to evaluate the plant microbial interactions or to diagnose potential plant diseases in space-based bioregenerative life support systems. In this study, we show how the MinION sequencing platform can be used to diagnose the opportunistic pathogen Fusarium oxysporum sensu lato, a fungal infection on Zinnia hybrida (zinnia) plants that were grown on the International Space Station (ISS) in 2015–2016. Genomic DNA from the infected plant material (root and leaf tissues) retrieved from the ISS were extracted and sequenced. In addition, pure cultures of Burkholderia contaminans, F. oxysporum sensu lato, and Fusarium sporotrichioides were used as controls to test the specificity of the bioinformatics pipeline developed. The results show that the MinION platform can be used to accurately differentiate between fusaria species and strengthens the case for using the platform as a rapid plant disease diagnostic tool in space.
1. Introduction
Crop production in the spaceflight environment is a complex process as it requires the use of controlled environment agricultural practices under novel environmental stimuli, such as microgravity conditions, elevated radiation levels, and other less-characterized stresses associated with the spaceflight environment. Yet, growing crops in the spaceflight environment to supplement essential nutrients to the astronaut's diet, aid atmospheric regeneration, and purify water is expected to have a profound impact on future human space missions. To ensure that successful cultivation of crops on long-duration crewed missions in space can be accomplished, a contingency plan to rapidly diagnose the onset of any plant disease and mitigate the risk of it spreading needs to be developed. Unfortunately, such protocols do not exist for current planned space missions.
In 2015, a naturally occurring plant disease outbreak was encountered onboard the International Space Station (ISS). Zinnia hybrida plants grown in a plant-growth system called Veggie between November 2015 and February 2016 (Massa et al., 2017) were subjected to an opportunistic fungal infection (Fig. 1). By December 22, 2015, three of six zinnia plants developed severe signs (e.g., aerial mycelium) and symptoms (e.g., chlorosis, necrosis, and wilt) of disease (Schuerger et al., 2021). The causal agent of the disease was not immediately diagnosed on-orbit due to the lack of any diagnostic tools onboard the ISS. Infected and symptomatic tissues were collected, frozen at −80°C, and returned to Earth for analysis beginning on May 18, 2016. The complete diagnostic procedure required ∼9 months to complete, and the causal agent of disease was identified as the opportunistic pathogen, Fusarium oxysporum sensu lato (Schuerger et al., 2021).

Fusarium oxysporum sensu lato signs (aerial mycelium; arrows) and symptoms (e.g., chlorosis, necrosis, and wilt) on Zinnia hybrida plants grown in the Veggie VEG-01C experiment onboard the ISS (Schuerger et al., 2021).
As we embark on long-duration space missions, the reliance on Earth-based diagnostic capabilities for plant diseases will diminish. Instead, new methods of diagnosing plant diseases in space will need to be developed so that informed agricultural practices can be used to maintain a sustainable cultivation of crops in space. The MinION sequencing platform has recently been shown to reliably and accurately diagnose plant diseases (Bronzato Badial et al., 2018; Filloux et al., 2018; Chalupowicz et al., 2019; Xu et al., 2021). In this study, we show preliminary results on how the MinION platform can be used to accurately diagnose a spaceflight-specific plant pathogen that was previously characterized by Schuerger et al. (2021).
2. Methods
2.1. Sample collection and isolates
Zinnia hybrida cv., “Profusion” plants were grown on the ISS in the Veggie hardware (VEG-01C) between November 2015 and February 2016 (Massa et al., 2017). Thirty-six days after the initiation of plant growth—on December 22, 2015—aerial mycelium was observed on the leaves and stems of zinnia plants (Fig. 1). Diagnosis of the causal agent revealed that F. oxysporum sensu lato acted as an opportunistic pathogen on the severely high-water stressed plants (Schuerger et al., 2021). The infected zinnia leaves grown onboard the ISS were stored continuously at −80°C on-orbit, during re-entry, and until returned to the National Aeronautics and Space Administration (NASA) Kennedy Space Center, Florida, laboratories on May 18, 2016. The initial diagnostic procedures lasted ∼1 month, after which all remaining tissues were again archived at −80°C until used in this study that began on November 1, 2020.
The same set of infected zinnia plant tissues previously characterized by Schuerger et al. (2021) (i.e., 202 mg of root tissue and 814 mg of leaf tissue) were designated as ISS root and ISS leaf samples. In addition, pieces of infected zinnia leaves and roots were placed on potato dextrose agar (PDA) (Fisher Scientific, Pittsburg, PA) and incubated at room temperature (∼24°C) for 2 days. Plant material along with any bacterial or fungal growth on these plated tissues were collected and designated as PDA-root or PDA-leaf samples, respectively. Moreover, pure cultures of Burkholderia contaminans, F. oxysporum sensu lato, and Fusarium sporotrichioides grown on trypticase soy agar (Becton Dickenson, Franklin Lakes, NJ) and PDA, respectively, for bacteria and fungi, were used as controls in this study.
2.2. DNA extraction
Genomic DNA was extracted with the MoBIO PowerSoil Kit (Qiagen, Valencia, CA) according to the manufacturer's protocol. The DNA was then purified with the Zymo DNA clean and concentration kit according to the manufacturer's protocol (Zymo Research, Irvine, CA). The quantity of DNA was assessed with a Qubit 2.0 fluorometer (Thermo Fisher Scientific, Waltham, MA) and quality was assessed with a 2100 Bioanalyzer (Agilent Technologies, Inc., Santa Clara, CA).
2.3. Nanopore DNA-PCR library preparation
The DNA-PCR library preparation was performed by using the Rapid PCR Barcoding Kit (SQK-RPB004) following the manufacturer's instructions (Oxford Nanopore Technologies, Oxford, UK). In brief, 1–5 ng of purified genomic DNA were combined with the fragmentation mix and incubated in a thermal cycler at 30°C for 1 min and then 80°C for 1 min. Amplifications of the tagmented DNA fragments were performed in PCR reactions by using the Nanopore rapid barcoding primers (SQK-RPB004) and the LongAmp Taq 2x Master Mix (New England Biolabs, Ipswich, MA) with the following cycling conditions: an initial denaturing step at 95°C for 3 min, followed by 14 cycles of 15 s at 95°C, 15 s at 56°C, and 6 min at 65°C, a final extension step at 65°C for 6 min, and a hold at 4°C.
The PCR products were then purified and separated by using 30 μL of Agencourt AMPure XP beads (Beckman Coulter, Indianapolis, IN). A total of 50–100 fmoles of barcoded genomic DNA were pooled and loaded onto a FLO-MIN106D R9 flow cell. Barcodes (BCs) 1–2 and 4–6 were run together as a single library. The flow cell was washed and stored at 4°C for several weeks before a second library comprising BCs 8–9 was run on the same flow cell. Sequencing was performed on a MinION device for a total of 19 h for each library.
BCs used in this study correspond to the following samples: BC01—PDA Root, BC02—PDA Leaf, BC04—B. contaminans, BC05—F. oxysporum sensu lato, and BC06—F. sporotrichioides, BC08—ISS Root, BC09—ISS Leaf.
2.4. Bioinformatics and data analysis
Sequencing data were based-called in real time with the MinKNOW software v3.3.2 through the MinIT device v19.05.2. All FASTQ files containing sequencing data were analyzed with Nanoplot v1.0.0 (De Coster et al., 2018). BCs were demultiplexed and adapters were trimmed with Porechop v0.2.4 (Wick et al., 2017). Reads were filtered for a minimum read length of 100 bp and minimum Q score of 7 by using Filtlong v0.2.0 (Wick, 2020).
Taxonomic classification of genomic DNA reads was performed with Kraken2 v2.0.8b (Wood et al., 2019) using a modified Kraken2-microbial database (
3. Results
In this study, we investigated the effectiveness of the MinION sequencing platform in identifying F. oxysporum sensu lato as the causative agent in infected zinnia plants grown on the ISS. The frozen infected tissue samples obtained from the ISS (Schuerger et al., 2021) were directly used for DNA extractions and small fragments (2–3 mm length) of infected zinnia root and leaf tissues were also incubated on PDA before genomic DNA extraction. The decision to incubate plant tissue on PDA plates was to obtain fresh microbial DNA as the original ISS zinnia tissues were frozen for nearly 4 years before this study, and the ability to extract high-quality genomic DNA for MinION sequencing was in question.
In addition, pure cultures of the bacterium—B. contaminans—and the fungi—F. oxysporum sensu lato and F. sporotrichioides—were used to test how well the pipeline could differentiate between the kingdoms and species (Fig. 2A). The DNA extraction results showed that it was possible to obtain high-quality genomic DNA directly from <1 g of infected plant tissue (Supplementary Table S1).

Samples and bioinformatics pipeline used to validate the plant disease diagnosis capability of the MinION sequencing platform.
The sequencing statistics (Supplementary Table S2) showed that the mean read lengths for both libraries were on average 3488 bp and the mean Q scores were >8. Sequencing data were analyzed through the bioinformatics pipeline (Fig. 2B) and reads from each Kraken report (Supplementary Tables S3–S9) were sorted based on the highest percentage of fragments covered by the clade and the rank code (S) for species. The top 13 species with the highest percentage, and >200 reads, were selected from each sample and visualized as a bubble plot (Fig. 3; Supplementary Table S10).
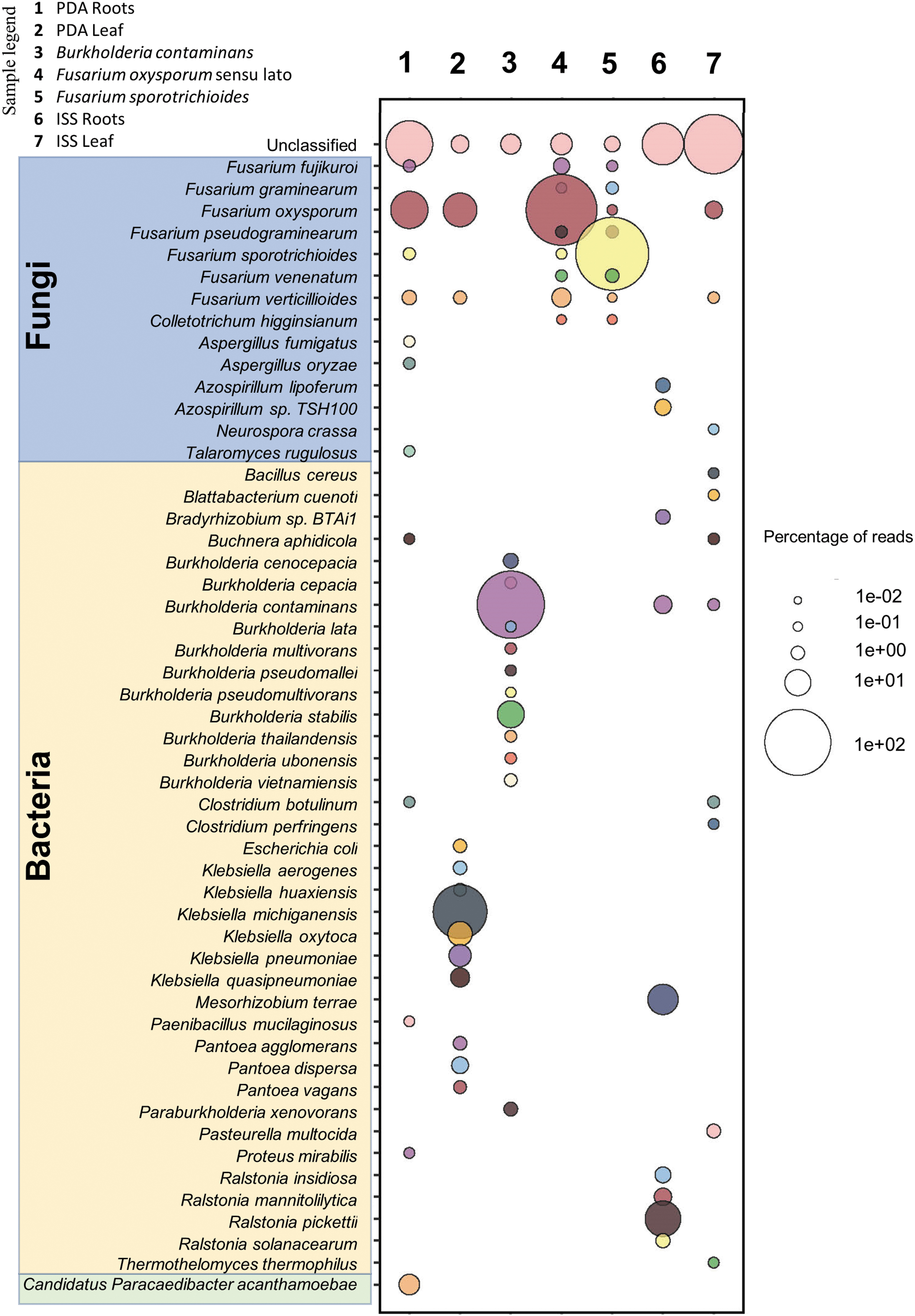
Taxonomy diversity of genomic DNA reads in plant samples and pure cultures analyzed by Kraken2. Percentage of reads from the top 13 species with >200 reads were selected from each sample and visualized a bubble plot. The size of the bubble indicates the percentage of reads assigned to each species. The corresponding samples for each barcode are as follows: (1) PDA roots, (2) PDA leaves, (3) Burkholderia contaminans, (4) Fusarium oxysporum sensu lato, (5) Fusarium sporotrichioides, (6) ISS roots, and (7) ISS leaves. Species highlighted in blue, yellow, and green belong to the following categories fungal, bacterial, and others, respectively.
The data show that the pipeline clearly distinguished between kingdoms and species. In each of the pure cultures (B. contaminans, F. oxysporum sensu lato, and F. sporotrichioides) >75% of all reads sequenced were mapped to the exact species used (Fig. 3). Interestingly, the ISS leaf tissue samples showed that a small number of active pores in the MinION flow cell—toward the end of its sequencing capability—generated <200,000 reads for each barcoded sample; but were enough to diagnose the causative agent of disease. F. oxysporum sensu lato was the leading species sequenced in the ISS leaf sample. Although F. oxysporum sensu lato was the leading species sequenced in the PDA root samples, and the second most sequenced species in the PDA leaf samples, it was not detected in the ISS root sample.
4. Discussion
MinION uses shot-gun sequencing technology that requires no prior knowledge or prediction about the possible causative agent, detection of any microbial organisms, and is limited to the known microbial sequences in the NCBI database. Currently, commercially available portable rapid plant disease diagnostic tools are based on immunodetection methods, PCR-based assays, and spectrometry-based protocols that require some prior prediction of the microbial organisms that are being tested to design antibodies or PCR primers for the analyses (Fang and Ramasamy, 2015; Martinelli et al., 2015).
The MinION sequencer has been successfully utilized onboard the ISS for 16S microbial analyses (Castro-Wallace et al., 2017; Stahl-Rommel et al., 2021). In this article, we show the feasibility of using the MinION for a different application—a space-based plant pathogen detection tool. Although several aspects of the protocol (i.e., purifications and library preparations) still require further development, automation, and testing for flight readiness; we have shown that the MinION platform used for microbial sequencing on the ISS can be applied to rapidly diagnose plant diseases in future space-based bioregenerative life support systems habitats or small plant-growth modules. Furthermore, had such a MinION-based protocol been established and available for the Veggie VEG-01C experiment, it is likely that the causal agent of the infected and symptomatic Z. hybrida plants could have been diagnosed within 48 h instead of the 9 months required by Schuerger et al. (2021).
The manual DNA extraction method of infected plant tissue showed that <1 g of material can be used to diagnose the causative agent. It is expected that the amount of diseased plant tissues required from fresh samples would be much less for diagnostic purposes (possibly similar to the amount of fresh weight used in the PDA samples, which was ∼50 mg) if the tissues were harvested directly from infected plants on-orbit. The sequencing data obtained from this study also highlight the benefits of using fresh samples for diagnosis. In Fig. 3, the titer of F. oxysporum sensu lato DNA in 4-year-old frozen roots was much lower than the reinvigorated fungal mycelium of the PDA culture. This observation is consistent with the earlier attempts to diagnose the zinnia plant disease by Schuerger et al. (2021) and supports the need to perform such diagnosis on-site.
Thus, these findings strengthen the rationale that the MinION sequencing platform—which is currently onboard the ISS—could also be utilized as a plant disease diagnostic tool. In addition, and to streamline the process, further development of the technology and protocols for processing the samples are necessary so that the entire procedure will require low demand for crew time. Nonetheless, it is imperative to advance the ability to investigate plant microbiomes and diagnose plant diseases on orbit to improve future space food production.
Footnotes
Authors' Contributions
N.J.S. performed the experimental work, data analyses, and led the article development. A.C.S. assisted in the conceptualization of the research and helped write the article.
Data Management Plan
The raw and processed MinION Oxford Nanopore sequencing data reported in this article have been deposited with NASA's GeneLab (GLDS-358). Original, high-resolution, and uncropped images used in this article will be posted in the University of Florida Institutional Repository (UFIR) at the website
Author Disclosure Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding Information
This study was supported in part by NASA Space Biology Postdoctoral Fellowship (80NSSC19K0520) awarded to N.J.S.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
Supplementary Table S7
Supplementary Table S8
Supplementary Table S9
Supplementary Table S10
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.