
Abstract
Subglacial environments on Earth offer important analogs to Ocean World targets in our solar system. These unique microbial ecosystems remain understudied due to the challenges of access through thick glacial ice (tens to hundreds of meters). Additionally, sub-ice collections must be conducted in a clean manner to ensure sample integrity for downstream microbiological and geochemical analyses. We describe the field-based cleaning of a melt probe that was used to collect brine samples from within a glacier conduit at Blood Falls, Antarctica, for geomicrobiological studies. We used a thermoelectric melting probe called the IceMole that was designed to be minimally invasive in that the logistical requirements in support of drilling operations were small and the probe could be cleaned, even in a remote field setting, so as to minimize potential contamination. In our study, the exterior bioburden on the IceMole was reduced to levels measured in most clean rooms, and below that of the ice surrounding our sampling target. Potential microbial contaminants were identified during the cleaning process; however, very few were detected in the final englacial sample collected with the IceMole and were present in extremely low abundances (∼0.063% of 16S rRNA gene amplicon sequences). This cleaning protocol can help minimize contamination when working in remote field locations, support microbiological sampling of terrestrial subglacial environments using melting probes, and help inform planetary protection challenges for Ocean World analog mission concepts.
Introduction
Drilling through glaciers and ice sheets for instrument deployment and sample collection presents significant engineering challenges (Talalay, 2016). An added difficulty is maintaining a clean drilling environment in order to ensure the integrity of materials collected for subsequent microbiological and geochemical analyses. Contaminated cleaning solutions, human handling, the field camp environment (i.e., dust or soil) or nontarget glacier ice that the drill must navigate through (see Schuler et al., 2023) can all introduce contamination. Thus, any drilling fluids or equipment utilized for subglacial access must be cleaned and verified, which can be burdensome in a remote field setting.
Given the pristine nature of subglacial aquatic environments, operational protocols that preserve these ecosystems is paramount. As such, subglacial ecosystems are protected under the Protocol on Environmental Protection within the Antarctic Treaty (Siegert and Kennicutt, 2018), and research activities involving access beneath the Antarctic Ice Sheet must demonstrate that impact to the natural ecosystem is minimized. A Code of Conduct (CoC) for the Exploration and Research of Subglacial Aquatic Environments was established to provide specific guidelines for the scientific ice drilling community (Doran and Vincent, 2011). This CoC set forth minimum standards to prevent contamination with recommendations relevant to this study presented here including the need to reduce the number of cells on instrumentation that enters subglacial environments (Recommendation 7) and researching and developing methods that reduce and monitor contamination (Recommendation 13).
Glacial systems are also unique analog sites that can inform operations on ice-covered extraterrestrial targets; thus, the guiding principles of the Committee on Space Research's (COSPAR) Planetary Protection Policy can also inform subglacial exploration. The goal of Planetary Protection (Cheney et al., 2020; COSPAR, 2021) is to prevent biological or organic contamination of natural celestial environments. Measures are taken to reduce bioburden and organic contaminants on spacecraft (forward contamination) in order to maintain the integrity of astrobiological investigations. When samples or spacecraft are returning to Earth, measures are taken to prevent the introduction of harmful extraterrestrial materials (backward contamination). Planetary protection is conducted by national space agencies in accordance with Article IX of the 1967 Outer Space Treaty and policies maintained by the Committee on Space Research (COSPAR, 2021). The US (NASA) and European (ESA) space agencies have detailed instrument preparation protocols for the exploration of icy bodies in our solar system (NPR 8715.24 [NASA Procedural Requirements, 2021]; NASA-HDBK-6022 [NASA, 2010]; ECSS-Q-ST-70-55C [ECSS-Q-ST-70-55C Working Group, 2008]).
The standard for bioburden reduction since the Viking Project (Shirey et al., 2017), which is approved by both NASA and ESA (ECSS-Q-ST-70-57C Working Group, 2013; Rummel and Pugel, 2019), is dry heat microbial reduction. In this method, hardware is heated under set conditions of temperature, humidity, and time. Other methods include physical removal of bioburden with solvents or foams and exposure of instruments to ultraviolet or gamma radiation, gas plasmas or vapor hydrogen peroxide (Rummel and Pugel, 2019). These protocols are typically performed in settings that control for contaminating particles (such as International Organization for Standards [ISO] certified cleanrooms). Regardless of the method employed, protocols must meet NASA's decontamination requirements. The NASA Standard Assay for measuring microbial contamination on spacecraft is designed to detect endospore-forming heterotrophic microorganisms (La Duc et al., 2004). Briefly, an area (25 cm2) of instrument surface is swabbed, placed into 10 mL of sterile phosphate buffer, and sonicated for 2 min (NASA Procedural Requirements, 2021). An aliquot of the buffer is directly plated onto TSA agar, while another aliquot is first heat-shocked (80°C for 15 min) and then plated and incubated at 32°C for 3–7 days, after which colony-forming units (CFUs) are enumerated. NASA and ESA have developed standards based on the probability that a contaminant microbial passenger might harm an intended astrobiological target and hinder life-detection missions. For Ocean Worlds, NASA aims for 4-log bioburden reduction on space hardware (NASA Procedural Requirements, 2021). While currently the only measure of spacecraft cleanliness is based on the abundance of spores on surfaces (NASA Procedural Requirements, 2021), another example benchmark of cleanliness, proposed by Benardini and Venkateswaran (2016), is the total concentration of adenosine triphosphate (ATP), an important biomolecule for all life, on the instrument. This study concluded that to achieve an acceptable level of cleanliness, a clean surface should contain less than 0.023 pmol per 25 cm2 of ATP.
At the time the Antarctic Treaty Subglacial Access CoC guidelines were developed, no subglacial environments in Antarctica had been directly accessed. Recently, the US Antarctic Program utilized hot water drilling approaches to sample subglacial lakes (Whillans and Mercer Subglacial Lakes) along the Siple/Gould Coast (Tulaczyk et al., 2014; Priscu et al., 2021). These projects generated large access holes (∼60 cm in diameter), down to a depth of ∼800 m for the multiple deployment of diverse instruments (Tulaczyk et al., 2014). Water used for drilling was first passed through a series of filters (2.0 and 0.2 μm in pore size) and a UV sterilization system (Priscu et al., 2013). The surfaces of down-hole instruments were spray-saturated with 3% H2O2 before deployment (Christner et al., 2014), although subsequent analyses of cleanliness were only conducted on drilling fluids (Michaud et al., 2020), which indicated these fluids met the standards of the CoC. Any disturbance due to accessing Whillans Subglacial Lake was considered “transient” as hydrological studies indicated the lake drains directly into the ocean on subdecadal time scales (Carter et al., 2013) and therefore was not subject to the same review as isolated lake ecosystems.
Melt probes offer an efficient, environmentally sensitive, and potentially cleaner means to access subglacial environments, in that they can be specifically designed for cleaning ease, utilize less fuel, and have a significantly smaller logistical footprint (∼1 ton of equipment transported in a single helicopter sling load) than other access approaches such as hot water drilling (for example, the Whillans Subglacial Lake project required 12 tractors pulling 26 sleds for drilling and sampling equipment; Rack, 2016). Another advantage is that melt probes can be operated in a (semi-) autonomous way, which qualifies them for use in future Solar System exploration missions. The IceMole is a novel thermoelectric melt probe developed under funding from the German Federal Ministry for Economic Affairs and Energy (BMWi) with project management under the German Space Agency (DLR) (Dachwald et al., 2014; Kowalski et al., 2016). It was designed for cleanability and minimal disturbance when accessing sample targets within and below ice. Our cleaning protocol for the IceMole was developed in collaboration with engineers to establish compatibility with materials, melt probe operations and integrity of sample collection. A key science objective for the IceMole was to navigate within a glacier and collect samples from a discrete interface within the ice. Blood Falls, Antarctica was identified as an ideal site for testing targeted collection of englacial fluid samples using the IceMole.
Blood Falls (Fig. 1A) is a distinct feature that forms at the end of an englacial conduit or conduits through which subglacial brine episodically discharges (Badgeley et al., 2017). It is a surface manifestation of an extensive subsurface aquifer below Taylor Glacier (Mikucki et al., 2015), thought to have been isolated for perhaps a million years (Elston and Bressler, 1981). Direct discharge collected at Blood Falls indicates it originates from a source that is geochemically and biologically distinct from the surrounding glacial ice (Mikucki and Priscu, 2007; Mikucki et al., 2009; Campen et al., 2019; Lyons et al., 2019). When subsurface brine is discharged to the surface at Blood Falls, it is rapidly altered by oxygenation, exposure to ultraviolet radiation, and/or mineral precipitation (Campen et al., 2019; Sklute et al., 2022); however, samples collected during active discharge events appear geochemically and biologically stable over multiple years (Campen et al., 2019; Lyons et al., 2019). Discharge events remain unpredictable; thus, for ecological studies, we endeavored to capture brine within the glacier, prior to discharge, in order to confirm the characteristics of the subsurface end-member brine. In a planetary context, targeting and sampling the conduit connecting the Blood Falls feature with its subsurface aquifer source offers a unique Ocean World analog. For example, melt probes could be used for targeted sampling of saline water bodies that may form at relatively shallow depths (∼1–5 km) within the thick ice shell, such as those predicted below lenticulae, the dark, dome-like, elliptically shaped features observed at the Europan surface (Chivers et al., 2021).

Blood Falls in the McMurdo Dry Valleys, Antarctica.
Due to the unique nature of the Blood Falls ecosystem, it receives additional protection beyond the CoC recommendations for subglacial sampling described above and is designated as an Antarctic Specially Protected Area (ASPA; Blood Falls is designated ASPA#172). ASPAs are features identified as having outstanding environmental, scientific, historic, aesthetic, or wilderness values (Siegert and Kennicutt, 2018). A permit is required to enter Blood Falls, and the entirety of the subglacial area that terminates in Taylor Valley is protected. Thus, all operations within the Blood Falls ASPA must be conducted with environmental sensitivity. This has some analogy to the COSPAR Planetary Protection Policy that defines Mars Special Regions as regions where terrestrial organisms may be able to replicate or have a high potential to harbor Martian life (COSPAR, 2021). Missions that may have direct contact with Special Regions or induce them through interaction of instrumentation with the planet (i.e., heating of permafrost, for example) must meet the most stringent planetary protection guidelines (Rummel et al., 2014; Kminek and Rummel, 2015; Rettberg et al., 2016; COSPAR, 2021).
We posit that the combination of special protections afforded to Blood Falls, its distinct geological and glaciological context, and the unique low-biomass ecosystem it contains makes our IceMole sampling expedition a relevant analog study of a life-detection mission concept in potential icy “Special Regions.” Environmental stewardship, planetary protection, and sample integrity requirements for geomicrobiology are synergistic goals; thus, we adopted procedures from both subglacial and space exploration and adapted them to operations in a remote field scenario. We developed a simple, field-executable approach for addressing clean access requirements which minimizes microbial contamination on sampling equipment. An array of biomolecules were measured in samples collected from the IceMole probe before and after cleaning for comparison with our target sample, the englacial brine collected at Blood Falls.
Description of the IceMole
The IceMole is a thermoelectric melting probe (Dachwald et al., 2014, Kowalski et al., 2016) that can melt its way through glacier ice. The probe is ∼2 m long with a square cross-section of 0.15 by 0.15 m for a total surface area of 1.245 m2. The IceMole has a heated melting head that integrates a rotating ice screw oriented in the drilling direction to maintain a predefined contact force. This unique attribute enables melting vertically into the ice, similar to conventional melting probes, as well as melting at an angle that deviates from strict vertical downward motion. The head of the IceMole is made from a Cu-Ni alloy and has a maximum heating power of 2.88 kW, laid out to maintain a forward melting velocity of 1 m/h in -17°C ice, such as expected for Taylor Glacier. The heating power of the IceMole head is distributed among 16 heating cartridges (0.2 and 0.16 kW) and thus can be differentially regulated in order to control melting velocity during operations. The IceMole body is made from Al and integrates powerful side wall heating systems (2 kW each) that can be used to avoid freezing stall and enable movement of the probe along a tractrix-type trajectory. The IceMole trajectory, including information on depth below the ice surface and attitude, can be tracked through a sophisticated onboard localization solution (Clemens and Schill, 2016; Kowalski et al., 2016).
The IceMole includes an internal and external sampling system. The internal system routes meltwater through a hollow proboscis within the ice screw; from there, water can optionally be pumped to the surface via the IceMole meltwater tubing or collected in a sample bag within the IceMole. The internal sample bag (as described in Kowalski et al., 2016) did not function during our field campaign at Blood Falls and is not reported on further here. The external sampling system consists of a borehole pump attached to the rear of the IceMole. Samples collected when using this pump passed through the IceMole meltwater system tubing and the pump mechanism.
Science operations—field camps
Camps were established on Canada Glacier in November 2013 and on Taylor Glacier above Blood Falls in November 2014. Field cleaning of the IceMole was first tested on the Canada Glacier, Antarctica, in 2013 (Supplementary Fig. S1). Canada Glacier is within the McMurdo Dry Valleys but outside the Blood Falls protected ASPA (Fig. 1). After demonstrating our field-based cleaning approach in 2013, the IceMole was deployed to Taylor Glacier the following season (2014–2015; Fig. 1), where it was used to collect samples of brine from the englacial conduit that feeds Blood Falls. The Taylor Glacier Camp (Fig. 1B) was within the MDV Antarctic Specially Managed Area (ASMA) as well as the Blood Falls ASPA (#172). Glacier camps were a ∼30–60 min hike from dwelling tents (Supplementary Fig. S1).
At both science operation camps, a clean tent (i.e., an 8' × 6' PolarHaven with a mounted wooden floor and a deployment hole) was established for cleaning and drill deployment operations only; a separate utility tent for mechanical operations was erected to compartmentalize procedures (Supplementary Fig. S1; Fig. 2). The clean tent was divided into two sections with tarps and clean plastic sheeting. The division limited airflow and created an entry room for changing footwear and dressing in full Tyvek suits prior to entering the second half of the tent, which was used exclusively for instrument cleaning and contamination testing. Once cleaning was complete, the IceMole was bagged in sterile flat wrap (FLEXPAK Nylon Cleanroom bags, KNF Corp., USA), and plastic coverings on the floor and benches were replaced. All scientists in the cleaning tent wore full Tyvek suits, booties, and facemasks (Fig. 2A). Laboratory exam gloves were worn at all times and changed regularly when handling supplies and equipment. Sterile nitrile gloves (Kimberly Clark Corp., USA) were worn when manipulating the probe during and after cleaning.

IceMole field-based cleaning operations. The exterior of the IceMole was cleaned in a PolarHaven constructed on the glacier. Inside the PolarHaven, (
Prior to the 2014 deployment at Blood Falls, the interior of the IceMole sampling tubing was flushed with 10% H2O2 and shipped to McMurdo Station, Antarctica. After arrival in Crary Labs (McMurdo Station), ultrapure water from the station's MD Millipore Milli-Q Ultrapure water system (MQ ultrapure water) was pumped through all internal and external sampling tubing. Following flushing of 2 × the volume of the IceMole system, aliquots (∼10 mL) of ultrapure water were collected into pre-combusted glass vials for ATP analysis or sterile tubes for evaluation of total colony forming units (CFUs). Larger volumes (∼1 L) were concentrated through 0.2 μm Supor filters (Pall, Corp., USA), stored at -80°C, and shipped on dry ice to the University of Tennessee, Knoxville (UTK) for nucleic acid extraction and analysis. Samples for direct counts of DNA-containing cells were preserved with 4% (w/v) buffered paraformaldehyde. Water chemistry from this test (i.e., major ions) is reported elsewhere (German et al., 2021).
Science operations—sampling of glacial melt and englacial brine at Blood Falls
On November 29, 2014, the IceMole began melting a channel near Blood Falls, Taylor Glacier (GPS coordinates: 77.72334°S, 162.26805°N, altitude: 42 m). Melt from drilling was continuously pumped out of the channel using the borehole pump. The cleaned pump tubing terminated in the clean tent where subsamples were collected from discrete depths (1 and 11 m) into sterilized vessels. On November 30, a sharp rise in conductivity values was detected in samples pumped to the surface, which indicated penetration into the subglacial brine conduit. At that point in time, the IceMole was at a depth of -15.2 m from the glacier surface, having melted a 16.6 m long channel. Once the probe penetrated the pressurized conduit, brine was pumped through 2 × the volume of the tubing before englacial brine collection. We estimate a volume of ∼100–150 L of brine was collected (Fig. 3) for microbial diversity (Campen et al., 2019) and geochemical analyses (Lyons et al., 2019). During sampling, the IceMole moved forward until reaching a final depth of -16.1 m at a channel length of 17.7 m. Thus, the IceMole entered the brine conduit exposing a maximum length of 1.1 m of its surfaces into the brine.

Schematic of IceMole conduit brine collection showing ATP concentrations estimated for various elements of drilling operations. An idealized representation of the extensive subsurface aquifer detected by airborne geophysical measurements (Mikucki et al., 2015; Foley et al., 2016) and radar evidence for basal crevassing at the base of the Blood Falls conduit (Badgeley et al., 2017). The connectivity between the aquifer and crevasse is not fully understood and depiction of features in this figure are not to scale.
Before each deployment, the IceMole was cleaned within the clean tents following a 10-step procedure (detailed in Table 1). Briefly, all exposed surfaces were spray-saturated with MQ ultrapure water (18.2 MΩ-cm) and then scrubbed with a 1% solution of Triton-X 100 (laboratory grade, Sigma-Aldrich Corp., USA) using sterile wipes and sterile swabs to remove any particulates (dust, sediments, etc.); then it was again rinsed with ultrapure water. Next, all surfaces were spray-saturated with 5% H2O2 (prepared with ultrapure water from the Crary Labs Milli-Q system) and allowed to sit for 15 min to destroy microbial cells. Previously, Leimena et al. (2010) showed that 5% and higher concentrations of H2O2 are effective at decimating microbial DNA and other cellular macromolecules at low temperatures (0–5°C). Surfaces were then wiped with sterile, DNA-free wipes (ITW Texwipe, USA) to remove any residual liquid. Removal (wiping off) of the H2O2 was in such a manner that no surface of the DNA-free wipe was used more than once. Although H2O2 breaks down to O2 and H2O (a first-order reaction where 2H2O2 → 2H2O + O2 with a reaction rate close to 1.4 × 10−4 s−1 at 5°C), residual H2O2 can corrode surfaces if bagged while still wet. Additionally, H2O2 may interfere with some of our bioburden assays, and wiping would remove any final debris or remaining cellular material. The cleaned and wiped IceMole was bagged in sterile flat wrap (KNF Flexpak, KNF Corp., USA) with ends folded over and tied. The bag was removed as the IceMole descended into the ice such that surfaces were not exposed following cleaning (Fig. 2B). Bagging the IceMole after cleaning maintained sterile conditions until drilling operations and allowed for handling and maneuvering the drill without introducing contamination.
Summary of Major Steps in Field-Based Cleaning Protocol for the IceMole
Summary of Major Steps in Field-Based Cleaning Protocol for the IceMole
Surfaces were swabbed with individually wrapped sterile CLASSIQ Swabs (COPAN part No. 164KS01) before and after cleaning the IceMole to assess the effectiveness of our procedure. Autoclaved square cut-outs (25 cm2) made of polysiloxane or polytetrafluoroethylene (Teflon) were placed over sections of the probe so that a distinct measured 25 cm2 area of the IceMole surface was swabbed (Fig. 2A). For each measurement (Supplementary Fig. S2), the swab was moistened with a prepared solution (either sterile DNA/RNA-free water [Fisherbrand, ThermoFisher Scientific, USA; referred to as FB ultrapure water] or autoclaved and 0.2 μm filtered phosphate-buffered saline [PBS] solution). Excess liquid was expunged by pressing the swab against the sterile interior tube wall; then the swab was applied to the surface and the 25 cm2 area was thoroughly covered while rotating the swab (according to the methods detailed in NASA-HDBK-6022 [NASA, 2010]; Supplementary Fig. S2). The tip of the swab was then snipped into its tube with flame-sterilized scissors. Swabs collected for direct counts of DNA-containing cells and CFUs on agar plates were placed in sterile falcon tubes containing 5 mL of autoclaved, 0.2 μm filtered PBS. Samples for direct counts were fixed with paraformaldehyde (4% final concentration). Swabs collected for ATP and Limulus amebocyte lysate (LAL) measurements were placed in ATP-free Biopur microcentrifuge tubes (Eppendorf, AG, Germany) containing 1 mL FB ultrapure water. Ambient air samples were collected by waving swabs in the air of the clean tent for ∼1 min. Replicate swabs of the IceMole surface were collected prior to cleaning (Step #1), prior to bagging with sterile flat wrap (Step #9), and immediately before deployment as the sterile bagging was progressively cut away (during Step #10). Controls were collected from the air and reagents throughout cleaning to determine background levels of biomolecules in the clean tent and in any lab spaces used for the subsequent processing of samples.
Assessment of bioburden on the IceMole
Tubes containing solution (PBS or FB ultrapure water) and swab tips were vortexed for 25 s before removing aliquots for analyses. Samples were collected from the surficial glacier ice surrounding the Blood Falls englacial conduit for background measurements using sterilized instruments for chipping and then melted in sterile vessels at 4°C. Bioburden measurements included lipopolysaccharides (LPS), total ATP, nucleic acids, direct cell counts and viable (colony) counts.
The Limulus amebocyte lysate (LAL) assay
The LAL assay is a rapid test for the presence of bacterial endotoxin (aka LPS), which is a distinctive component of Gram-negative bacterial membranes (Eigenbrode et al., 2009). Endotoxins are a group of lipid and carbohydrate complexes that specifically coagulate LAL derived from horseshoe crabs. The turbidity created from this reaction can be measured as absorbance at 405 nm. Four aliquots of 25 μL were taken from swab collections, vortexed in FB ultrapure water, and analyzed for lipopolysaccharides with the handheld, single-use, disposable, lab-on-a-chip system LOCAD-PTS (Charles River Laboratories, USA) with LAL assay cartridges. Results were measured and reported as endotoxin units (EU); the cartridges used in this study had a 0.01–1.0 EU per mL detection limit. LAL-test results can be obtained within 15–30 min, which would provide a rapid assessment of our cleaning rigor prior to deployment.
Adenosine triphosphate (ATP)
Adenosine triphosphate is an important biosignature molecule used by all cells to transport and convert energy for metabolism. We used the luminescent ATP Kit 266-311 (BioThema Co., Sweden) to quantify total (both intra- and extracellular) ATP concentration. This kit has a detection limit of 10−17 mol and employs firefly luciferase, which catalyzes a light-producing bioluminescent reaction. The luminosity, which is directly proportional to the concentration of luciferase (and ATP) in the sample, was measured with a GloMax 20/20 Luminometer (Promega Corp., USA). The GloMax 20/20 device has low power requirements (0.5A @ 100–240V) and can be run on a small solar battery pack in the field.
DNA containing cells (direct counts)
Sample aliquots (4 mL) were incubated with the nucleic acid stain SYBR Gold (Thermo Fisher Scientific, USA) for 15 min, as described in Mikucki and Priscu (2007), and collected on a 25 mm, 0.2 μm pore size black polycarbonate filter for enumeration using an Axio Imager M2 epifluorescence microscope (Carl Zeiss, Germany) at 100 × magnification.
Assessment of colony-forming units (CFUs)
Aliquots (100 μL) of PBS from vortexed swab samples were plated onto Bacto tryptic soy broth (TSB) agar plates (VWR International, USA) and incubated at 4°C and ambient temperature (20–25°C) for 1 month. While this approach would capture all viable heterotrophs, it does not test for the proportion of these viable cells capable of forming endospores because we did not subsequently perform a heat-shock (80°C for 15 min) step. Additional growth experiments were conducted at UTK including TSB plates and TSA molten agar plates incubated at 25°C (per NASA protocols; NASA-HDBK-6022 [NASA, 2010]). Aliquots from swab samples were also incubated on Marine agar ATCC 2216 (a heterotrophic marine salts medium) and R2A agar ATCC 2258 (a low-nutrient heterotrophic medium used regularly for oligotrophic environments) at 4°C to test for growth under more ecologically relevant conditions. Marine agar has been shown to enrich organisms from Blood Falls discharge (Chua et al., 2018), and microbes from the surrounding ice have been successfully grown on R2A (Mikucki et al., 2004). Aliquots (4 mL) of IMflowthru and conduit brine were filtered onto 0.2 μm gridded sterile cellulose nitrate membrane filters (Sartorius) and incubated on R2A and Marine agar plates.
Nucleic acid analyses of potential microbial contaminants
Samples for DNA analyses were obtained by swiping a full side of the IceMole with a 47 mm Supor filter (Pall Corp., USA) collected before and after cleaning. Filters were stored at -80°C for shipment on dry ice to UTK. MQ Ultrapure water (∼1 L) from the water system and water that was first pumped through the IceMole (IMflowthru) and its sampling tubing was also collected on filters and stored at -80°C for shipment on dry ice to UTK. Nucleic acids were extracted from filters with the PowerSoil DNA extraction kit (MP Bio) according to the manufacturer's protocol. DNA extracts were PCR-amplified using the broad-range bacterial primers (8F and 1391R) or universal primers (515F and 1195R) for 16S rRNA gene analyses using PCR conditions described by Purcell et al. (2014).
PCR products were purified with the Wizard PCR clean-up kit (Promega Corp., USA). A clone library was constructed for PCR products from filtered samples using the PCR-4 TOPO vector (Promega Corp., USA). Vectors were then transformed into competent E. coli cells (One Shot TOP10 Chemically Competent; Invitrogen, USA) and selected for on LB agar (ATCC 1065) with kanamycin (50 μg per mL). Plasmid DNA was isolated and digested using RsaI enzyme for restriction length polymorphism analyses to select unique clones for further sequencing. Colonies from the MQ ultrapure water test (the only samples besides the target conduit brine where CFUs were observed) were prepared for sequencing by picking isolated colonies with a sterile tip for “colony pick” PCR using conditions described by Purcell et al. (2014). Both cloned and isolate 16S rRNA genes were sequenced (∼600 bp) at the UTK Division of Biology sequencing facility using Sanger sequencing.
Sequences were quality checked and manually aligned with BioEdit Software (version 7.2); a consensus sequence was generated and queried against the NCBI database, using blastn (Altschul et al., 1990) to determine the identity of the clones and isolates. Sequences were deposited in Genbank (accession numbers OR123785-OR123790). A BLAST database was generated using ncbi-blast+ toolkit that included all end-member brine paired-end amplicon sequences reported by Campen et al. (2019) which contained four brine libraries generated from three samples of active discharge collected by hand in 2004 and the conduit sample collected with the IceMole in 2014. Possible contaminant sequences detected in this study were queried against the end-member brine database. Amplicon sequences with 95% or greater identity to any clones or isolates from this study were identified as possible contaminating operational taxonomic units.
Results
Biomolecule monitoring
Results are reported per 25 cm2 of the IceMole surface or per liter for the MQ ultrapure water test. Where applicable, we identify whether our measurements address NASA Procedural Requirements or Antarctic Treaty CoC recommendations. Collectively, our decontamination protocol reduced the concentration of biomolecules on the exterior of the IceMole by all metrics measured (Table 2).
Summary of Biomolecule Data Collected on the IceMole Surface, Milli-Q (MQ) Ultrapure Water Pumped through the IceMole and Various Controls
Summary of Biomolecule Data Collected on the IceMole Surface, Milli-Q (MQ) Ultrapure Water Pumped through the IceMole and Various Controls
Proposed NASA standard for ATP (Benardini and Venkateswaran, 2016).
LAL value corresponding to low risk (after Morris et al., 2010).
bd = below detection; nd = not determined.
The disposable LOCAD PTS system is currently used by NASA for microbial monitoring on the International Space Station (Maule et al., 2009) and to assess planetary protection goals in analog environments (Eigenbrode et al., 2009) because it provides rapid results with a portable device. However, we had several challenges with this assay: (1) the LAL test cartridges were cost-prohibitive (when compared to the ATP assay), which limited our ability to run a high number of replicates; (2) the assay appeared sensitive to H2O2 (data not shown), which hindered replicates of LAL analysis; and (3) when we returned from the field, Charles River issued a recall of our particular lot of chips. Thus, we are not highly confident with our few results reported here. LAL concentrations were obtained in 2013 prior to cleaning and averaged 1.023 ± SD 0.030 EU per 25 cm2. Background controls were more variable (0.455 and 0.260 EU). Two clean measurements were obtained, one following cleaning (0.245 EU per 25 cm2) and one while the sterile bag was being removed from the IceMole immediately before deployment (0.20 EU per 25 cm2). These concentrations correspond to a low risk level designation for spacecraft, 0.13–0.25 EU per 25 cm2 (Morris et al., 2010). The LAL assay was not pursued during the 2014 season due to reliability concerns and costs.
Our approach to cleaning significantly reduced ATP burden on the IceMole surfaces (2013 p = 0.02, and 2014 p = 0.00005) with a percent reduction of 91.3% in 2013 (log reduction = 1.06) and 99.3% in 2014 (log reduction = 2.14 log). Prior to cleaning, the concentration of ATP on the IceMole surface was variable and likely reflects the degree of handling. In 2013, the IceMole arrived in McMurdo Station pre-cleaned from FH Aachen, and handling by personnel prior to deployment was minimal. Total ATP concentrations on the surfaces prior to cleaning in 2013 were low (0.15 ± SD 0.11 pmol ATP per 25 cm2) and variable (Fig. 4). Following cleaning, total ATP concentrations were reduced by an order of magnitude (0.016 ± SD 0.003 pmol ATP per 25 cm2) and were less variable. Measurements taken as the sterile bag was removed for deployment on Canada Glacier remained low (0.015 ± SD 0.004 pmol ATP per 25 cm2), indicating that sterile bagging of the instrument was effective in maintaining cleanliness during drilling preparations. In 2014, however, the IceMole was handled in the Crary Lab for mechanical repairs and our MQ ultrapure water test prior to field cleaning which may have contributed to the higher concentrations of ATP on the pre-cleaned surface when compared to 2013 (5.73 ± SD 2.08 pmol ATP per 25 cm2). Following cleaning in 2014, the IceMole had an average concentration of 0.042 ± SD 0.022 pmol ATP per 25 cm2. IceMole surfaces are not directly comparable to air background samples; however, they can provide some measure of the tent conditions. Canada Glacier tents contained 0.019 ± SD 0.013 pmol ATP per swab while Taylor Glacier tents contained 0.061 ± SD 0.010 pmol ATP per swab. The camp and environment surrounding Taylor Glacier were considerably dustier during our fieldwork. FB ultrapure water blanks measured 0.0140 ± SD 0.0002 pmol for an equivalent volume used for tent swabs (5 mls). Total ATP concentrations were low in the MQ ultrapure water (2.60 ± SD 0.03 pmol per L) and in aliquots of IMflowthru (3.95 ± SD 0.60 pmol per L). The glacier ice above the Blood Falls conduit contained 9.49 ± SD 3.9 (n = 10) pmol ATP per L, while concentrations in brine from the conduit target were 160.3 ± SD 64.6 (n = 6) pmol ATP per L.
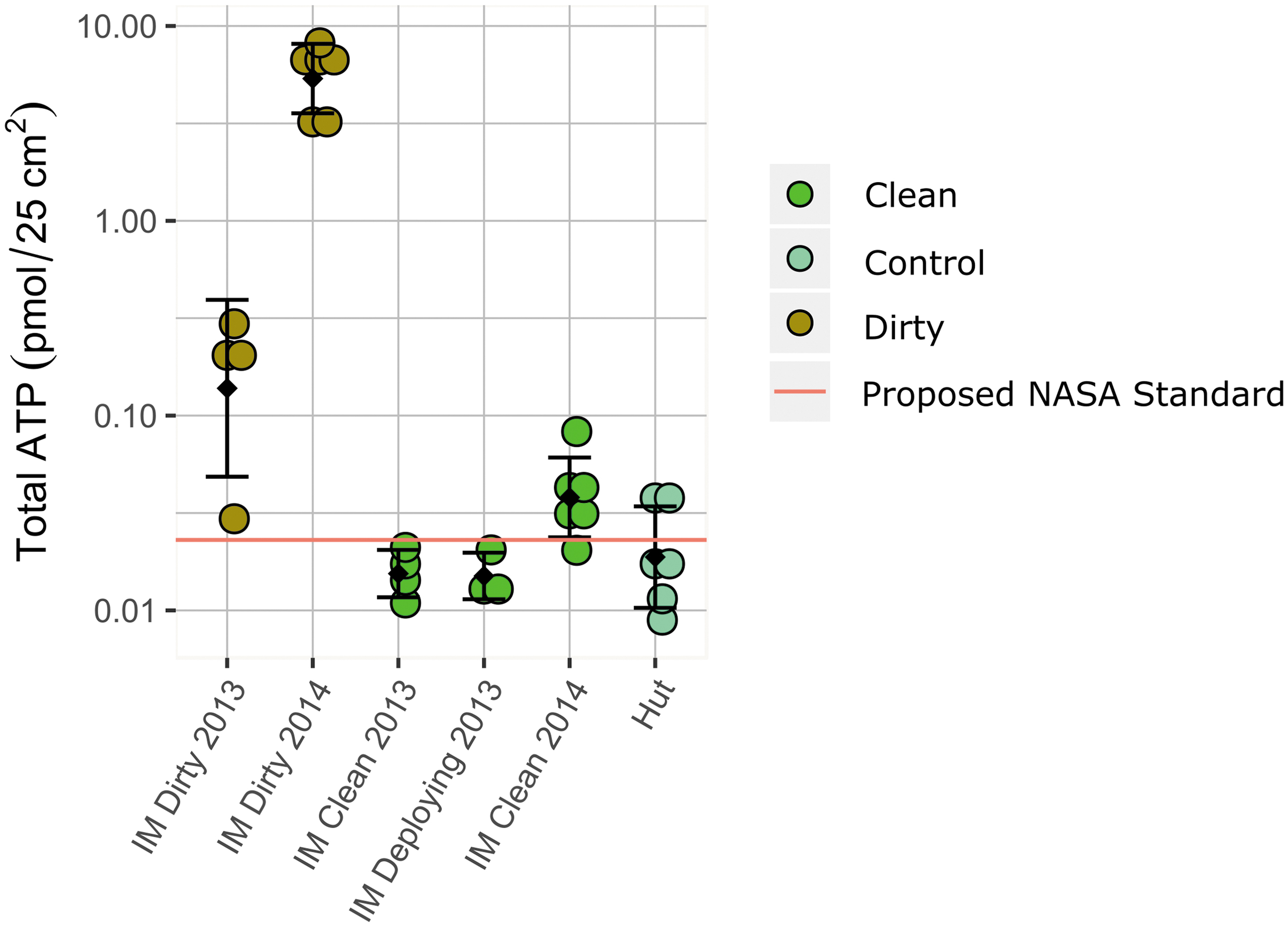
Total ATP concentrations measured on IceMole surfaces. Measurements of total ATP on the “dirty IceMole” before cleaning (brown circles), following cleaning (light green circles). Controls represent Fisherbrand (FB) ultrapure water reagents and air swabs within the tents where measurements were taken (teal circles). The red line represents the proposed NASA standard for spacecraft surfaces of 0.023 pmol per 25 cm2 (Benardini and Venkateswaran, 2016).
Prior to cleaning, direct counts of DNA-containing cells were 1.16 × 104 cells per 25 cm2 on the IceMole surface. Following cleaning, slides prepared with aliquots from surface-collected swabs were not discernable from our blanks, including swab samples collected from the IceMole surface as the sterile bag was removed for deployment. Cell numbers in the MQ ultrapure water samples flushed through the IceMole were at the limit of detection and highly variable (8.4 × 102 cells per mL ± SD 6.2 × 102 cells per mL; n = 5). The average number of cells in samples collected from Taylor Glacier ice overlying the conduit contained 3.29 × 103 ± SD 1.9 × 103 per mL (n = 5) with greater concentrations observed near the surface, possibly due to the presence of cryoconite holes (Foreman et al., 2007). These values are similar to previously published estimates for Taylor Glacier ice (∼2.3 × 103 cells per mL; Mikucki and Priscu, 2007). Glacier melt collected during the descent of IceMole from 1 and ∼11 m depth samples contained ∼5.3 × 103 ± 2.2 × 102 cells per mL (n = 2) and ∼1.2 × 103 ± 7.0 × 102 cells per mL (n = 3) respectively. Brine samples collected with the IceMole in 2014 were 4.88 × 103 ± SD 8.0 × 102 cells per mL (n = 4); previously reported values for cells in the end-member brine range between 1 × 104 and 7.6 × 105 cells per mL (Mikucki and Priscu, 2007). Thus, the conduit brine numbers could be an underestimate, as many cells appeared ultrasmall (<1 μm; data not shown), which may be a stress phenotype (Kuhn et al., 2014).
None of the IceMole surface swabs yielded microbial colonies on agar plates, either before or after cleaning, following one month of incubation. Three distinct morphologies were identified in the MQ ultrapure water (IMflowthru) incubations on R2A; however, total CFUs were highly variable and too low to reliably count (i.e., less than 10 CFUs). Blank control plates did not yield colonies. Only one colony formed on Marine agar plates incubated with IMflowthru.
A single light pink colony was cultivated on R2A agar inoculated with MQ ultrapure water from Crary Lab (i.e., water collected directly from the MQ ultrapure water system and not pumped through the IceMole). The partial 16S rRNA gene sequence of this isolate had 100% identity to Methylobacterium brachiatum strain B0021 (Kato et al., 2008). Colonies observed on R2A plates incubated with IMflowthru (MQ ultrapure water passed through the IceMole) yielded three unique phylotypes (Supplementary Table S1) including the same Methylobacterium sp. identified from the MQ ultrapure water system, an isolate with 98% identity to Rhodopseudomonas boonkerdii strain NBRC 106595 a metal-tolerant member of the Bradyrhizobiaceae (Noisangiam et al., 2011), and an isolate with 100% identity to Sphingomonas zea strain JM-791 (Kämpfer et al., 2015). The Sphingomonas strain was also the sole isolate from the MQ ultrapure water test (IMflowthru) to grow on Marine agar.
DNA was not detected, and no PCR products were obtained from the IceMole surface following cleaning. Measurable DNA was not obtained from MQ ultrapure water alone, and attempts to amplify PCR products were unsuccessful. PCR-amplified DNA extracts of the IMflowthru yielded six unique clones (Supplementary Table S1) based on RFLP analyses. Clone (IMclone-1) had 100% sequence identity to M. brachiatum and 100% sequence identity to our isolates from the MQ ultrapure water system and IMflowthru. The five other phylotypes included IMclone-2 with 99% identity to Sphingobium limneticum strain 301 (Chen et al., 2013), IMclone-3 with 99% identity to Pseudomonas azotoformans strain DSM:18862 (Iizuka and Komagata, 1963), and IMcone-4 with 99.5% sequence identity to Stenotrophomans maltophilia strain NCTC10257. Two fungi were also detected with high identity to Naganisha uzbekistanesis strain CB 968 (100%) and Filobasidium magnum strain JCM 9038 (99%).
We subsequently blasted the few clones and cultures identified during the MQ ultrapure water test (Supplementary Table S1) against the end-member brine paired-end sequences from Campen et al. (2019) to assess possible contamination in our final sample collected with the IceMole. Our search yielded a total of 33 possible contaminating amplicon sequences with 95% sequence identity or greater (in the ∼250 bp V4 region) out of 49,147 total sequences, which combined comprised 0.067% of the total sequences. Nine potential contaminating sequences out of 14,234 (or 0.063%) were detected in brine samples collected with the IceMole in 2014. These abundances are below most filtering thresholds (Karstens et al., 2019). Of these nine sequences from the IceMole collected sample, only 5 shared 100% sequence identity (252 bp) with two possible contaminants. Four amplicon sequences were identical to the Methylobacterium sp. IMclone-1; IMisolate-1), which was also detected in the MQ ultrapure water system. One amplicon sequence was identical to IMclone-2 (related to Sphingobium sp.). Other potential contaminating sequences were found exclusively in the 2004 hand-collected samples. The fungal contaminants did not yield amplicon matches in any of the brine samples.
Discussion
Monitoring the concentration of contaminating biomolecules relative to a desired sample target is crucial for ecological studies in general, but specifically when addressing Antarctic Treaty CoC for exploring Antarctic subglacial aquatic environments. As detailed in Section 4 on drilling and entry, Recommendation 4.3 states, “As a provisional guideline for general cleanliness, [drilling equipment that will enter the subglacial aquatic environment] should not contain more microbes than are present in an equivalent volume of the ice that is being drilled through to reach the subglacial environment.” Using these guidelines, we can compare bioburden measured on the IceMole to the volume of ice the melt probe passed through before entry into the subglacial target.
Our most complete data set was total ATP concentrations, and we use these numbers to discuss possible contamination scenarios (ATP values in Table 2; schematic Fig. 3). As the IceMole descended through the glacier, all meltwater was pumped out of the borehole to the surface to prevent mixing of glacial melt with conduit brine. Subsampling of the meltwater enabled discrete measurements during drilling, corresponding to various depths in the glacier (1 and 11 m depths; Table 2). These subsamples showed little variation to that of surface-collected glacier ice samples. Thus, we combined all borehole melt and glacier ice measurements for an average concentration of 9.5 pmol ATP per L in the glacial ice overlying the conduit (Table 2). We estimate that passage of the IceMole through the 17.7 m long borehole generated ∼398 L of meltwater which, based on our measurements, would contain an average of 3780 pmol of ATP.
Following penetration of the IceMole into the conduit, brine was pumped for twice the volume of the tubing and discarded prior to collecting end-member brine samples. Once direct sampling of brine began, greater than 100 L was acquired from the conduit via pumping: there was no evidence that we pumped the conduit dry, and geophysical surveys suggest the volume of the aquifer below the Taylor Glacier is at least 1.8 × 1011 L (Mikucki et al., 2015). ATP in the sampled brine was 160.30 ± 64.5 pmol per L ATP; assuming the aquifer is a homogeneous mixture, we estimate the aquifer would contain ∼28.9 mol total ATP. If we assume the 100 L we collected was the total volume of brine in the conduit, as a conservative estimate, then the end-member brine within the conduit would contain >16,000 pmol of ATP.
Glacier melt generated by the IceMole was pumped from the borehole to the surface, and the subsurface brine target is presumed to be under pressure; therefore, it is unlikely that any melt from the overlying glacier ice (or its ATP contents) entered the Blood Falls conduit during sampling operations. Thus, we only consider potential contamination from the surfaces of the IceMole probe. Based on a surface area of the IceMole of 1.245 m2 and the average concentration of ATP on the cleaned probe (∼0.042 pmol per 25 cm2), we calculate the IceMole had ∼21 pmol of ATP on its exterior when it was deployed into the Taylor Glacier and melted into the Blood Falls conduit. If the entirety of the IceMole entered the conduit, it could have delivered 21 pmol of foreign ATP into the brine, which is 2 orders of magnitude less ATP than was detected in the volume of overlying ice melted with the IceMole and 3 orders of magnitude less than what was in the englacial brine. Because possible contaminating concentration of ATP on the IceMole was less than the overlying ice that the probe traversed, we conclude that, using this biomolecule as a tracer, the Antarctic Treaty CoC standards for subglacial access into a subglacial aquatic environment were met.
Microbial cells were not measurable on the cleaned IceMole; therefore, estimating possible introduction of cells into the conduit from the melt probe is not possible (Table 2). Cells were detected in IMflowthru during the MQ ultrapure water test, and it is possible the IceMole carried some level of cells within its tubing. However, the IceMole sampling tubing was pumped dry prior to deployment, and all meltwater was pumped to the surface. The CoC recommends that “the number of microbial cells contained in or on the volume of any material or instruments added to or placed in [subglacial environments] should not exceed the minimum concentration of microbes in the basal glacial ice being passed through” and given the limited available data on basal ice, should not “exceed a concentration of 102 cells per ml.” By our measurement, cells in the overlying ice were an order of magnitude higher than the CoC recommendations, and cells on the IceMole were undetectable; we therefore find that the instrument used to sample the englacial conduit contained less than the number of cells in the overlying half meter of ice.
Missions that aim to investigate Martian special regions, even if they do not include life-detection experiments, must meet a surface bioburden level of ≤30 spores, as measured by the NASA standard assay (COSPAR, 2021). Because the NASA standard assay only counts spores that are cultivable, aerobic, resistant to heat shock, and grow at 30°C in a nutrient-rich medium, this approach would not detect the vast majority of unculturable organisms (Crawford, 2005). Here we assayed for aerobic, heterotrophic growth in the absence of a heat shock, an approach which should be inclusive of aerobic heterotrophic spores in addition to non-spore-formers. All possible contaminants detected in this study were aerobic heterotrophs; however, none of our samples from the cleaned IceMole surface yielded microbial colonies following one month of incubation, thus achieving “less than 30 cells” cultured from the probe. However, despite low or nil growth on heterotrophic plates, average ATP concentrations were slightly above the proposed NASA standards on the cleaned probe in 2014. Spores contain significantly lower concentrations of ATP than vegetative cells (Fajardo-Cavazos et al., 2008), and our measurements were of total (intercellular and extracellular ATP); thus, it is hard to conclude the source of this ATP. Still, this finding indicates that cells or their biomolecules may have persisted on the IceMole surface, albeit at very low levels. Heterotrophic plates and molecular sequencing identified contaminants in the MQ ultrapure water test and included a Stenophomonas and Sphingomonas spp., known spore-formers; however, we were unable to detect these sequences in our IceMole-collected sample.
Phylogenetic identification of possible contaminants can help inform downstream analyses of diversity as well as potential harm. A microbe that can proliferate in a target environment is of greater concern, even if it is present on an instrument in low numbers. None of the identified contaminant colonies or DNA sequences from the IceMole surface or MQ ultrapure water test were detected on plates inoculated with Blood Falls conduit brine collected with the IceMole (full data not shown; Chua et al., 2018). Very few sequences related to the Methylobacterium and Sphingobium spp. were detected in our final IceMole sample. All other possible contaminants were only detected in samples from 2004, which were not collected with the IceMole, indicating these contaminants likely originate in the Crary Lab and/or standard sampling supplies.
The Methylobacterium genus has been implicated as a DNA contaminant in nucleic acid extraction kits (Salter et al., 2014); however, it is also a common environmental microbe. Kit contamination seems unlikely in our study as we did not amplify product in our extraction blanks and the Methylobacterium sp. we detected was viable and cultured directly from the MQ ultrapure water system. This may be the case for the Sphingobium sp. as well, but we were not able to cultivate this organism from the MQ ultrapure water alone. Our analyses are not necessarily comprehensive of all possible sources of contamination; however, using several methods, including molecular techniques, we attempted to identify potential microbial passengers in order to provide a more comprehensive assessment of sample integrity. While there is no current standard to control for contaminants in 16S rRNA gene studies, identifying taxa detected in controls and comparing them to the results from biological samples is a recommendation under discussion (Eisenhofer et al., 2019).
Conclusions
Melt probes offer an ideal tool for minimally invasive, clean sampling of subglacial environments; their logistical footprint is significantly smaller than that of hot water drills, providing a more mobile platform for sub-ice exploration projects. Melt probes have also been identified as important tools for the exploration and sampling of Ocean Worlds such as Europa and Enceladus (Phillips et al., 2021). The use of novel engineering tools often requires additional manipulation in the field before deployment; therefore, effective, low-impact means to clean on site are critical for success. Advancing this technology will require that analog mission operations consider planetary protection requirements.
We demonstrated a simple field-based decontamination protocol for melt probes that can be used when collecting samples in remote, glaciated regions. These cleaning methods can also be applied to other tools used for microbiological sample collections in polar regions, such as borehole-deployed instruments or ice core collection tubes. The IceMole surface was effectively cleaned by using a low concentration of a common laboratory detergent (Triton X-100) and 5% H2O2, eliminating the need for cleaners that may alter the natural geochemistry (such as acids or other solvents). Measurements used to ensure cleanliness on site included rapid result assays such as total ATP concentrations on surfaces, in the air, and in reagents. Downstream measurements performed after sampling, such as cultivation and nucleic acid analyses, allowed for more precise identification and tracking of sources of potential contaminants, as potential contaminant “passengers” can originate from multiple sources throughout the workflow of an expedition. In addition to identifying possible contaminants resistant to cultivation, molecular approaches can provide insight into potential sources of contamination. Here we identified a possible contaminant originating from the laboratory ultrapure water system that was later detected in both samples collected with the IceMole and in archived samples, albeit in extremely low abundances. Combined, these data allowed us to confirm integrity of sequencing, cultivation, and other geomicrobiological results from englacial brine samples.
Our results indicate that we achieved the standards set forth by the Antarctic Treaty CoC for access into a subglacial aquatic environment. While we did not measure the presence of spores, we did test for heterotrophic microbial growth using NASA-recommended approaches, followed by phylogenetic identification to assess our ability to meet cleanliness targets, which has important implications for both terrestrial and planetary science. The migration of subsurface aquifer brine to accessible depths within the Taylor Glacier (within tens of meters) provides a rare opportunity to study the ecology of Antarctic groundwater. Because subglacial discharge events are unpredictable and surface discharge rapidly mixes with glacial melt, drilling into the englacial conduit offers an important alternative for sample acquisition. The end-member brine contains a relatively low number of DNA-containing cells (∼104–105 cells per mL) of low diversity (∼477.0 ± 191.2 taxonomic units) (Campen et al., 2019); thus, accounting for contamination is critical. Identifying specific contamination types and their sources can help interpret results and allows for continued protocol improvement. Our data suggest that possible contamination of the sample collected with the IceMole was extremely low. Collectively these studies support the validity of conducting microbiological sample collection with melt probes.
In a planetary context, technical readiness level studies and mission concept operation demonstrations will require sample acquisition at analog field sites. Our field-based cleaning protocol can assess NASA-proposed minimal requirements of total residual ATP on space hardware. Investigations into the persistence of hardy organisms including, but not limited to, spore-formers that survive the cleaning process would further elucidate potential contamination and inform future subglacial and astrobiological missions to icy moons, including a Europa Lander Mission (Lorenz et al., 2011; Hand et al., 2017). Understanding how melt probes interact with the locally heated environment during descent would enable accounting for in situ contamination from glacial ice as well as the possibility of dragging ice-immured microbes into protected ecosystems, such as subglacial aquatic systems.
Footnotes
Acknowledgments
Field logistics was provided by the Antarctic Support Contractor for the United States Antarctic Program. We are particularly grateful to Geordan McQuiston and the McMurdo Station “Carp Shop” for their efforts in designing and constructing our Science operation camps. Feedback from anonymous reviewers improved this manuscript.
Author Contribution
JM, JK, ST, BD, and BL designed the MIDGE project; JM, ID, and MT designed the cleaning plan. JM led the field team; JM, RD, AP, MC, and DG collected samples and ran analyses. JM, CS, MT contributed to data analyses, figure development, and writing of this manuscript. All authors provided critical feedback and helped shape the research, analysis, and manuscript. This work was made possible by collaboration of all members of the MIDGE and EnEx Science Team (
Funding Statement
This work was supported by the National Science Foundation Office of Polar Programs ANT-1144178 and ANT-1344348 (JAM).
Competing Interest Statement
The authors declare no conflict of interest.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
Abbreviations Used
Associate Editor: Christopher McKay
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.