
Abstract
Modeling risks for the forward contamination of planetary surfaces from endemic bioburdens on landed spacecraft requires precise data on the biocidal effects of space factors on microbial survival. Numerous studies have been published over the preceding 60 years on the survival of diverse microorganisms exposed to solar heating, solar ultraviolet (UV) irradiation, vacuum, ionizing radiation, desiccation, and many other planetary surface conditions. These data were generated with diverse protocols that can impair the interpretations of the results due to dynamic experimental errors inherent in all lab protocols. The current study (1) presents data on how metal surfaces can affect spore adhesion, (2) proposes doping and extraction protocols that can achieve very high recovery rates (close to 100%) from aluminum coupons with four Bacillus spp., (3) establishes a timeline in which dried spores on aluminum coupons should be used to minimize aging effects of spore monolayers, (4) confirms that vacuum alone does not dislodge spores dried on aluminum coupons, and (5) establishes that multiple UV irradiation sources yield similar results if properly cross-calibrated. The protocols are given to advance discussions in the planetary protection community on how to standardize lab protocols to align results from diverse labs into a coherent interpretation of how space conditions will degrade microbial survival over time.
Introduction
The search for life on Mars remains a top priority for the Mars Exploration Program (Garvin et al., 2001) and Mars astrobiology (Des Marais et al., 2008; Horneck et al., 2016). To protect this search, the Committee on Space Research (COSPAR) has adopted a series of planetary protection categories and protocols to guide space-faring nations on how to appropriately prepare spacecraft for diverse missions to Mars (see review by Frick et al., 2014). A Category IVa mission to Mars does not include a life-detection payload, and the bioburden on spacecraft surfaces prior to launch is set at <300 spores/m2 and <3 × 105 spores for the entire spacecraft. A Category IVb mission does include a life-detection payload, and the mission engineers can choose either a full-system or a subsystem sterilization process depending on the design of the life-detection payload and sample handling device. The hardware and spacecraft bioburdens for Category IVb missions are <0.03 spores/m2 and <30 spores for the surface bioburden of the life-detection hardware. Category IVc missions are those that will land on, or near, Special Regions on Mars (defined as regions that may support an extant microbiota or be conducive to the proliferation of dispersed Earth microorganisms during the mission; see Kminek et al., 2010; Rummel et al., 2014). Allowed bioburdens for IVc missions are dependent on the expected bioburdens of landed hardware and based on experimental needs; the protocols can be equal to the guidelines given above or be more stringent.
Planetary protection protocols have been developed to sample and monitor the cleanliness of spacecraft during payload processing to meet the above goals. In addition, part of the evaluation process is to model how the launched spacecraft bioburdens will survive the Earth-Mars transit and the surface environment once spacecraft land. Several studies have evaluated the biocidal nature of the martian surface and set limits for temperature, water activity, UV irradiation, ionizing radiation, and salt tolerance for expected microbial survival and growth on Mars (e.g., Schuerger et al., 2003, 2005, 2006; Beaty et al., 2006; Kminek et al., 2010; Rummel et al., 2014; Cockell et al., 2016). In these studies, between 15 and 22 biocidal factors are listed that will work alone or in combination to inactivate landed bioburdens from Earth. However, there remains no established Mars microbial survival model that confirms that Earth microorganisms will be inactivated on Mars over short or long periods of time.
Such a microbial survival model for Mars will require precise inactivation kinetics for numerous biocidal factors when applied alone and in combination to yield overall predictions. The current literature, in general, is consistent in that the inactivation kinetics for specific microbial species when exposed to vacuum, solar UV irradiation, solar heating, or ionizing radiation follow similar decay kinetics for each factor when considered alone (e.g., Moeller et al., 2008, 2012; Wasserman et al., 2012; Zhang et al., 2018). The kinetics for heat and ionizing radiation tend to be linear over wide ranges of the biocidal variables. However, vacuum and UV models tend to exhibit exponential decay kinetics with obvious tailing of survival over time (e.g., Cerf, 1977; Nicholson, 2003; Schuerger et al., 2003, 2006; Schubert and Beaudet, 2011; Vaishampayan et al., 2012). Except for a few papers (e.g., Moeller et al., 2012; Schuerger et al., 2019; Moores and Schuerger, 2020), there is a paucity of multifactorial biocidal experiments or modeling of the above biocidal space conditions.
An important factor in developing microbial survival models is a set of standardized protocols for creating and sampling populations of test microorganisms under simulated space or martian conditions. Creating uniform and smooth monolayers is perceived as crucial to developing accurate inactivation kinetic models for spacecraft targeted for planetary bodies in which life-detection missions might be sent (e.g., Mars, Europa, Enceladus). Recently, several studies have proposed methods for creating uniform bacterial spore monolayers by using the microdrop (i.e., droplet) (Mancinelli and Klovstad, 2000; Schuerger et al., 2003), dry deposition (Edmonds et al., 2009), airbrush (Levy et al., 2011; Raguse et al., 2016), and filtration/transfer (Noell et al., 2013) protocols. A key problem that motivated many of these studies was the difficulty in consistently creating smooth spore monolayers with the microdrop technique in which spore or cell suspensions in carrier fluids are applied to glass or metal sample coupons; artifacts of stacked multilayered spore deposits and the formation of a coffee-ring effect were reported (Yunker et al., 2011; Raguse et al., 2016).
However, of the many spore deposition protocols discussed briefly above, the microdrop technique is the simplest and easiest to perform. The objectives of the current project were to (1) develop protocols for depositing uniform and smooth monolayers when using the microdrop protocol on flat aluminum coupons, (2) develop a reliable extraction protocol that achieves >90% recovery of applied spores, and (3) determine the effects of UV and vacuum on spore survival and recovery from coupons. The current study is the first in a series of papers that will describe and suggest a standard set of protocols relevant to processing hardware and spacecraft prior to launch to planetary surfaces like Mars.
Methods
Microbial protocols
Cultures of Bacillus subtilis HA101, B. subtilis 168, and B. pumilus SAFR-032 were grown in a spore-germination medium and processed as described previously (Schuerger et al., 2003, 2006). Spores of B. atrophaeus ATCC 9372 were purchased from Mesa Labs (Lakewood, CO, USA). All spores were stored at 4°C until required. To characterize the numbers of viable spores recoverable from spacecraft materials, spore-doped coupons were processed in 50 cc polypropylene conical tubes with 20 mL of sterile deionized water (SDIW) as the carrier fluid using a Most Probable Numbers (MPN) assay described by Mancinelli and Klovstad (2000) and Schuerger et al. (2003).
Spores were affixed to uncoated aluminum coupons at the rate of ∼2 × 106 spores per coupon—in 200 μL drops—by pipetting cell suspensions in either SDIW or 40% ethanol (B. atrophaeus only) (as described below) onto individual coupons. Spores in suspensions were allowed to settle overnight onto the upper surfaces of coupons, maintained at 24°C, and then dried under a sterile airflow within a NuAire biosafety cabinet (model NU-440-600, Class 2, Type A2, Plymouth, MN, USA).
Spore imaging protocols
Two forms of direct imaging were used to characterize the quality and uniformity of spore monolayers on spacecraft materials. The spores were either imaged via light microscopy (Olympus S2X-12 Dissecting Scope with an Olympus DP-11 Digital Camera, Olympus Corporation, Center Valley, PA, USA) or with scanning electron microscopy (Hitachi S-4000, Hitachi High-Tech America, Inc., Schaumburg, IL, USA). Spores on aluminum coupons for scanning electron microscope (SEM) imaging were first inactivated by exposing the monolayers to UVC (200–280 nm) irradiation derived from a xenon-arc illuminator (model 6262 UV lamp, Oriel Instruments, Stratford, CA, USA). Spores were exposed to UVC photons in excess of 140 kJ m−2, in which spores would have received at least 4 × the normal biocidal dosage for B. subtilis HA101 spores on aluminum coupons (Schuerger et al., 2006). Liquid fixation of the spores was not possible because such a procedure would have washed spores from the coupons; thus, some morphological artifacts were observed on imaged cells. However, this was deemed to be tolerable because the goal was to examine the spore densities in the monolayers on the spacecraft materials and not to evaluate spore ultrastructure on the coupons.
Spacecraft materials tested
Three types of spacecraft materials were used in the current study. First, uncoated aluminum coupons, measuring 55 × 18 × 0.5 mm, were purchased from Seton, Inc. (Buffalo, NY, USA). Second, the uncoated aluminum coupons were treated with a chemical conversion coating in order to passivate the aluminum. Chemfilm (MIL-DTL-5541; also known as Alodine or Iridite) coatings were applied to uncoated aluminum 6061 coupon surfaces by emersion into chromium oxide solutions as either Class 1A (smoother layer) or Class 3 (rougher layer) coupons. Figure 1 depicts the surfaces of the uncoated aluminum 6061, Class 3, and Class 1A coupons with SEM. The Chemfilm Class 3 coating was the roughest of the three options and was dropped from these experiments due to concerns that spores would be ensnared or trapped under the chromium oxide surface defects and be lost to the assays described below. Individual coupons were considered as separate replicates in all assays.

Spacecraft materials used in the current study were composed of either uncoated aluminum 6061 (
Uncoated aluminum coupons were used to evaluate three spore recovery procedures. Spore monolayers of B. subtilis HA101 were applied to uncoated Al-coupons and dried as described above. Separate coupons with monolayers were processed in sterile 50 cc conical tubes that each contained 20 mL of SDIW. The recovery protocols were as follows: (1) only SDIW was present as the recovery fluid in the 50 cc tubes, and the tubes were vortexed for 2 min; (2) coupons in SDIW were sonicated for 2 min in a water-submersion sonication system set at 35 kHz (model 75D, Aquasonic Ultrasonic Cleaner, VWR International, LLC, Atlanta, GA, USA); or (3) coupons in SDIW were vortexed with 1 g of sterile silica sand per tube for 2 min interspersed with vigorous hand-shaking of tubes every 30 s. Results were compared to T = 0 min samples of spores in cell suspensions not previously dried on coupons.
Spore viability versus spore recovery over time
As before, uncoated Al coupons were used to assess whether spore viability versus spore recovery changed over time. A fresh population of B. subtilis HA101 spores was created within 1 week of the following experiment. Fresh spores were diluted and approximately 2 × 106 spores applied per coupon. Cohorts of the dried monolayers were processed for spore recovery every 7 days for 12 weeks using only the SDIW + silica sand protocol as the recovery process. First, the stock spore suspension from which the coupons were prepared was maintained at 4°C for the duration of the experiment. Every 7 days, the stock suspension was measured for viable spores by the MPN assay. Next, a subset of the dried coupons was processed via the MPN protocol described above to determine the numbers of recovered spores per week over time. All dried monolayers on coupons were created at the same time, and within 2 days of starting the experiment.
Biocidal effects of three UV sources on spore viability
Three UV illuminators—xenon-arc (model 6262, Oriel Instruments), deuterium (D2) (model 68840, Oriel Instruments), and mercury-line (254 nm) (model 50-212-695, Fisher Scientific, Pittsburg, PA, USA)—were used to characterize the biocidal effects of UV irradiation on spore viability. Spectra from all three illuminators were collected with an Optronics OL-754 high-resolution UV-VIS spectrometer (Optronic Laboratories, Orlando, FL, USA) between 200 and 800 nm. Plots of the three UV spectra obtained between 200 and 400 nm were previously published by Schuerger et al. (2003). The OL-754 can integrate the UVC, UVB (280–320 nm), and UVA (320–400 nm) bands separately and provided individual band fluence rates. The illuminators were adjusted so that the UVC fluence rates were equal to 3.25 W m−2 (i.e., 195 J m−2 min−1) at the upper surfaces of spore monolayers. Thus, the fluence rates were approximately equal, and the effects of the spectra could be evaluated for each of the sources. Spore monolayers of B. subtilis HA101 were exposed for 1, 5, 60, 480, or 1440 min and then processed with MPN assays.
Residual spores on coupons after MPN assays
Four Bacillus spp.—B. subtilis HA101, B. subtilis 168, B. atrophaeus ATCC 9372, and B. pumilus SAFR-032—were tested to determine whether all spores were removed from Chemfilm Class 1A aluminum coupons by the SDIW + silica sand MPN protocol. Spores were prepared in 40% filter-sterilized ethanol to the desired concentrations. Monolayers of approximately 2 × 106 spores per coupon were prepared as described above; a cohort of coupons was removed for a pre-MPN SEM imaging pipeline; and a second cohort of coupons was processed through the MPN protocol, air-dried, and placed into a post-MPN SEM imaging pipeline.
Effects of vacuum on spore removal from Al coupons
Two Bacillus spp.—B. subtilis 168 and B. pumilus SAFR-032—were tested to determine whether spores could be dislodged by exposure to low pressures. Spore monolayers were created on Class 1A coupons, as described above, and the coupons were processed with the SDIW + silica sand MPN protocol. Separate cohorts of dried monolayers were placed in either sterile 10 cm diameter glass Petri dishes (horizontal orientation with monolayers in the up direction) or sterile 50 cc conical tubes (vertical orientation). The conical tubes were loosely sealed to permit the equilibration of internal headspace gases at low pressure; glass Petri dishes were naturally vented. Samples were then placed on the upper surface of a cryogenic fluid thermal control plate designed by the author and used in combination with a liquid nitrogen (LN2) thermal platform described by Schuerger et al. (2008).
Once the coupons of both Bacillus spp. were in place, the Planetary Atmospheric Chamber (PAC) was closed, the internal atmosphere pumped down to 7 hPa ±0.2 hPa (Exp-1) or 0.025 hPa ±0.005 hPa (Exp-2), and held at 24°C for 7 days. The PAC internal atmosphere—composed of an Earth-standard atmosphere including 78% pN2, 21% pO2, and the remaining balance of trace gases—was bled into the PAC system by a mass-flow controller set at 25 cc per min.
Following exposure to the 7 or 0.025 hPa pressures for 7 days, the PAC internal atmosphere was repressurized to 1013 hPa (sea level average), and the samples were aseptically returned to a NuAire biosafety cabinet. The coupons were aseptically and gently removed from the glass Petri dishes and 50 cc conical tubes and processed with the SDIW + silica sand MPN protocol. Liquid trypticase soy agar (TSA) at 48°C was added to the glass plates and conical tubes at the rate of 15 mL per device. The 15 mL of liquid TSA in each conical tube was spread out on the inner walls of the tubes by rotating the conical tubes until the TSA hardened. All TSA-treated samples were incubated at 30°C for 5 days. It was assumed that microcolonies of dislodged spores would be clearly visible within the TSA layers in the glass dishes or conical tubes. After 5 days, the glass dishes and tubes were carefully examined for the growth of bacteria entrapped within the TSA. If no colonies were observed, then dispersal of spores was unlikely to have occurred during the 7-day assays.
Statistics
Statistics were conducted with version 9.4 of the PC-based Statistical Analysis System (SAS) (SAS Institute, Inc., Cary, NC, USA). Data were analyzed as either untransformed data or with a 0.25-power transformation to induce homogeneity of treatment variances. However, all data in figures are presented as untransformed values. Results were then subjected to ANOVA followed by protected least-squares mean separation (LSmeans) tests with PROC GLM in SAS (P ≤ 0.05); numbers of replicates are given in figure legends when appropriate.
Results
Rough surface textures, pits, cracks, and defects in spacecraft materials can significantly affect the survival rates of attached microorganisms by allowing spores or cells into UV-protected niches. Three spacecraft materials were tested here to determine the effects of surface qualities on spore attachment and recovery. Of the three materials tested, uncoated aluminum 6061 coupons were the smoothest with the lowest level of observed defects (Fig. 1A). Chemfilm Class 3 coated coupons exhibited the most surface defects (Fig. 1B), and Chemfilm Class 1A (Fig. 1C) exhibited an intermediate level of cracks and surface roughness compared to the other two materials. Chemfilm Class 3 coupons were not tested further in the current study due to their extreme roughness and surface defects.
The protocol used here for creating smooth monolayers on uncoated Al or Class 1A coupons relied upon the settling of spores in the 200 μL droplets of SDIW overnight (approximately 18 h), the attachment of spores to the coupon surfaces while hydrated, and then a gentle drying process of the SDIW in a biosafety cabinet sterile air-flow. In each trial, approximately 80% of the B. subtilis HA101 spores affixed to coupons exhibited smooth monolayers (Fig. 2A, 2B, 2C) that were easily identifiable by visual inspections. In contrast, approximately 20% of the monolayers exhibited severe stacking artifacts in which the spores formed swirls of multilayered spores (Fig. 2D, 2E, 2F).

Light microscope images of high-quality (
In preliminary experiments in which only SDIW was used as an extraction fluid, the numbers of spores recovered were approximately one order of magnitude (1 log) lower than the spore densities applied to aluminum coupons (data not shown). Thus, three extraction protocols were tested to determine whether the 1-log reductions could be improved or eliminated. First, the three extraction protocols were (1) SDIW +1 g sterile silica sand, (2) SDIW + sonication, and (3) SDIW alone as the extraction fluid. The SDIW + silica sand protocol routinely yielded much higher spore numbers than either the SDIW alone or SDIW + sonication extraction protocols (Fig. 3). Moreover, the SDIW + silica sand protocol yielded almost the same numbers of spores as were originally applied to the coupons; Figure 3 shows an approximately 15% reduction between the T = 0 min controls and the SDIW + silica sand extraction protocol. All subsequent experiments used the SDIW + silica sand extraction protocol.
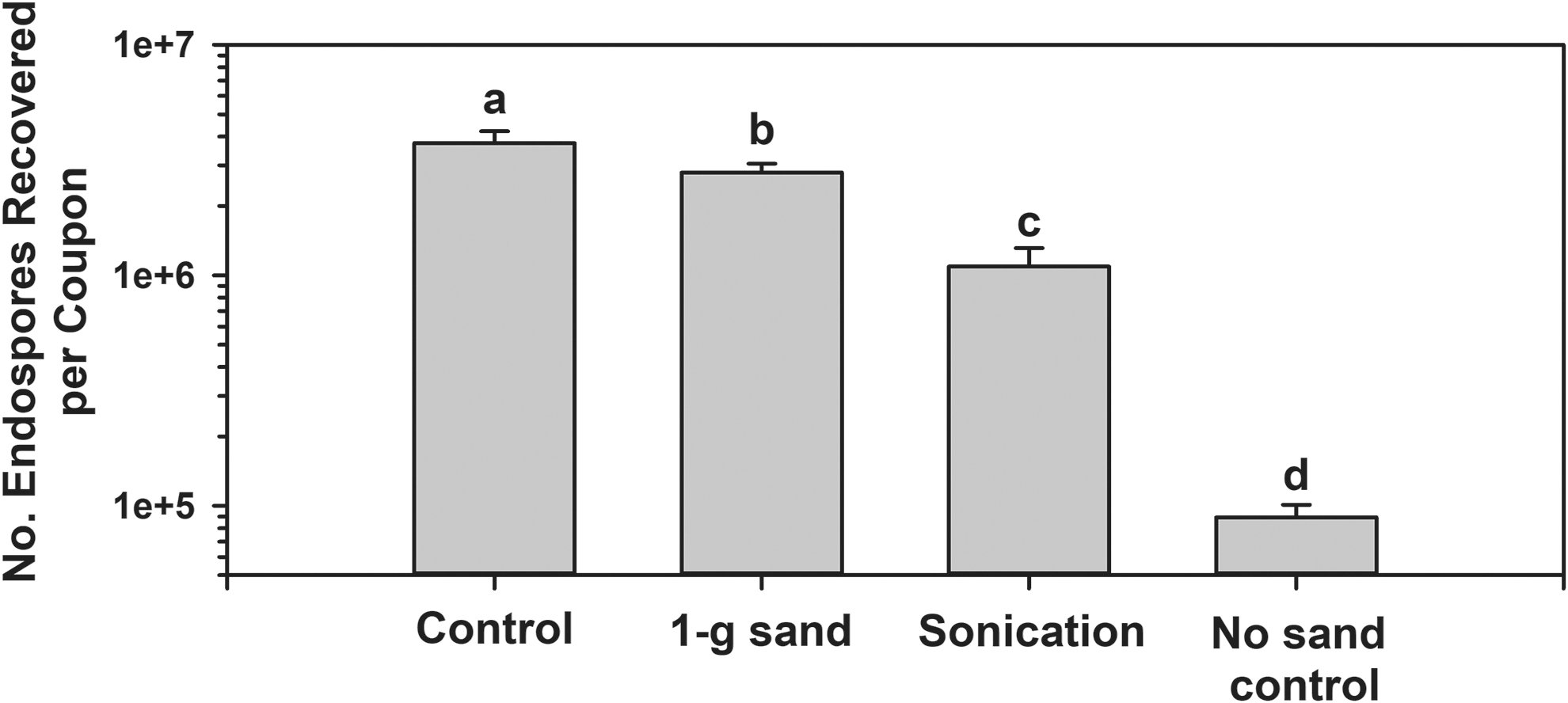
Recovery of Bacillus subtilis HA101 spores from uncoated aluminum 6061 coupons using three methods of extraction. First, the Control data indicates the number of spores in the inoculum suspension of spores at T = 0 min. The extraction protocols were (1) use of 1 g of sterilized silica sand in the extraction tubes, (2) sonication for 2 min, or (3) no sand in the extraction tubes (see text). All extraction tubes were filled with 20 mL of SDIW. Results were analyzed as untransformed data with ANOVA followed by a LSmeans test; treatments followed by diverse letters were significantly different (P ≤ 0.05; n = 12).
To determine whether the age of dried monolayers on coupons could alter the recovery of spores, an experiment was conducted in which the recovery rates from coupons was measured over a 12-week time period. First, fresh spores of B. subtilis HA101 were created 1 week prior to initiating the following experiment. A large population of 100 cohort monolayers on uncoated Al-coupons was created and stored at 24°C until assayed. Spores in SDIW were stored at 4°C until assayed. Every 7 days, samples were withdrawn from the SDIW liquid culture and diluted, and the viable spore numbers were estimated by the MPN assay. In addition, between 4 (weeks 1–7) and 6 (weeks 8–12) coupons were randomly picked from the cohort of 100 coupons and assayed every 7 days to determine whether the recovered numbers decreased over time. Figure 4 demonstrates that the spore numbers in the SDIW culture maintained their viability at approximately 2 × 106 spores per 200 μL over the 12 weeks of the experiment. In contrast, the recovered numbers of spores from pre-dried monolayers decreased by approximately 90% (1-log) over the same 12 weeks. Thus, only monolayers created and held at lab conditions for less than 3 weeks were used for all subsequent experiments.
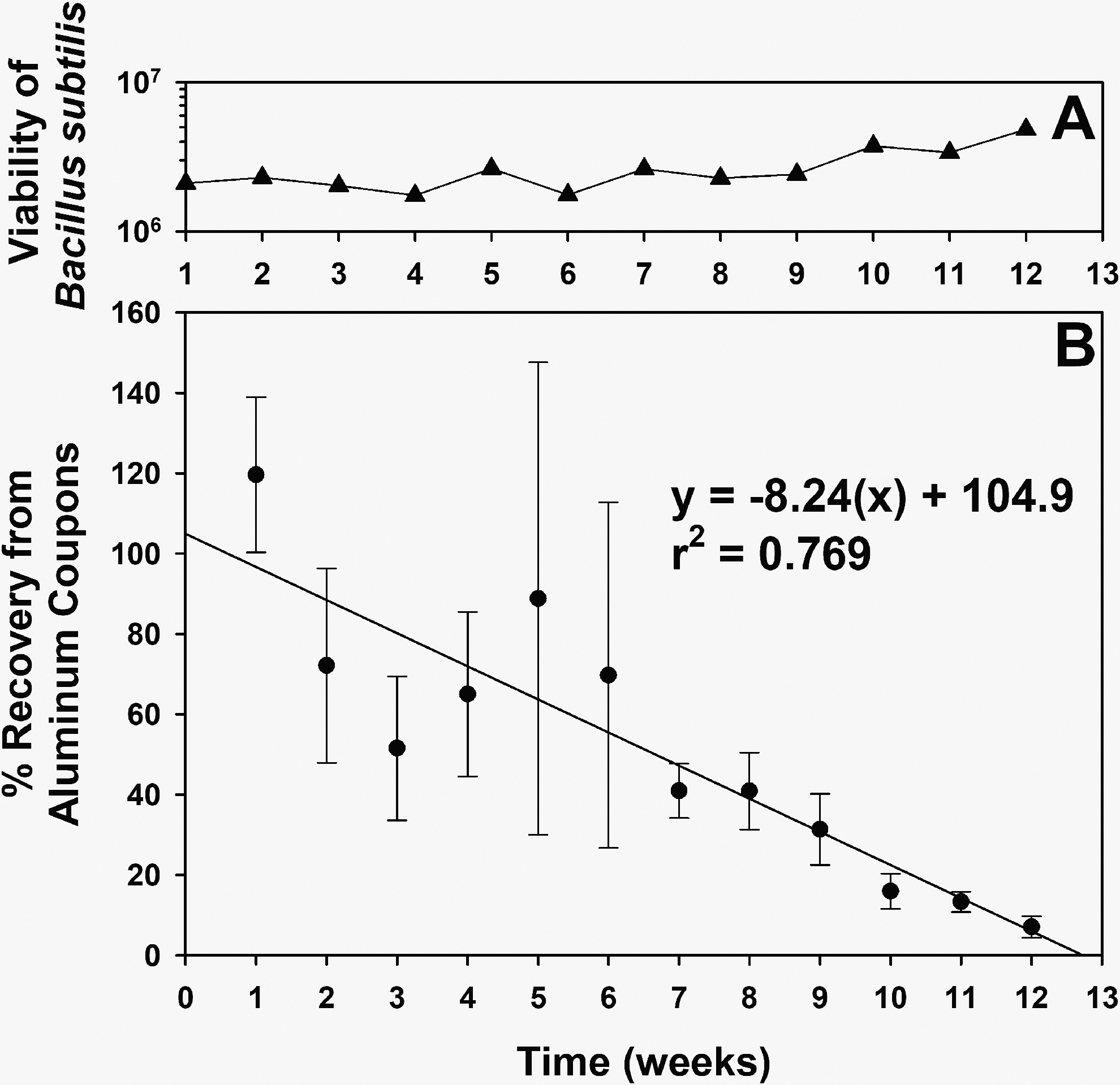
Recovery of Bacillus subtilis HA101 spores from uncoated aluminum 6061 coupons over time. Spores were prepared as described in the text, stored at 4°C in SDIW, or applied to Al-coupons at the rate of approximately 2 × 106 spores per coupon. The stored spores in SDIW were assayed every 7 days to confirm there was no loss of viability over time (
When exposing spores to UV irradiation, researchers typically use one of three UV illuminators: (1) a 254 nm Hg-line lamp, (2) a deuterium (D2) illuminator, or (3) UV photons from a xenon arc lamp. However, no data were found in the astrobiology literature that directly compared the efficacy of these three UV illumination sources. An assay was set up that cross-calibrated these three UV illuminators to expose smooth monolayers of B. subtilis HA101 under the same fluence rate of 3.25 W m−2 (195 J m−2 s−1). Results indicate that there were no significant differences among the three UV sources at any given time-step or in the way the biocidal effects of UV irradiation for each UV source decreased uniformly over time (Fig. 5). Thus, direct comparisons can be made among diverse studies in which one of these three UV sources is used.
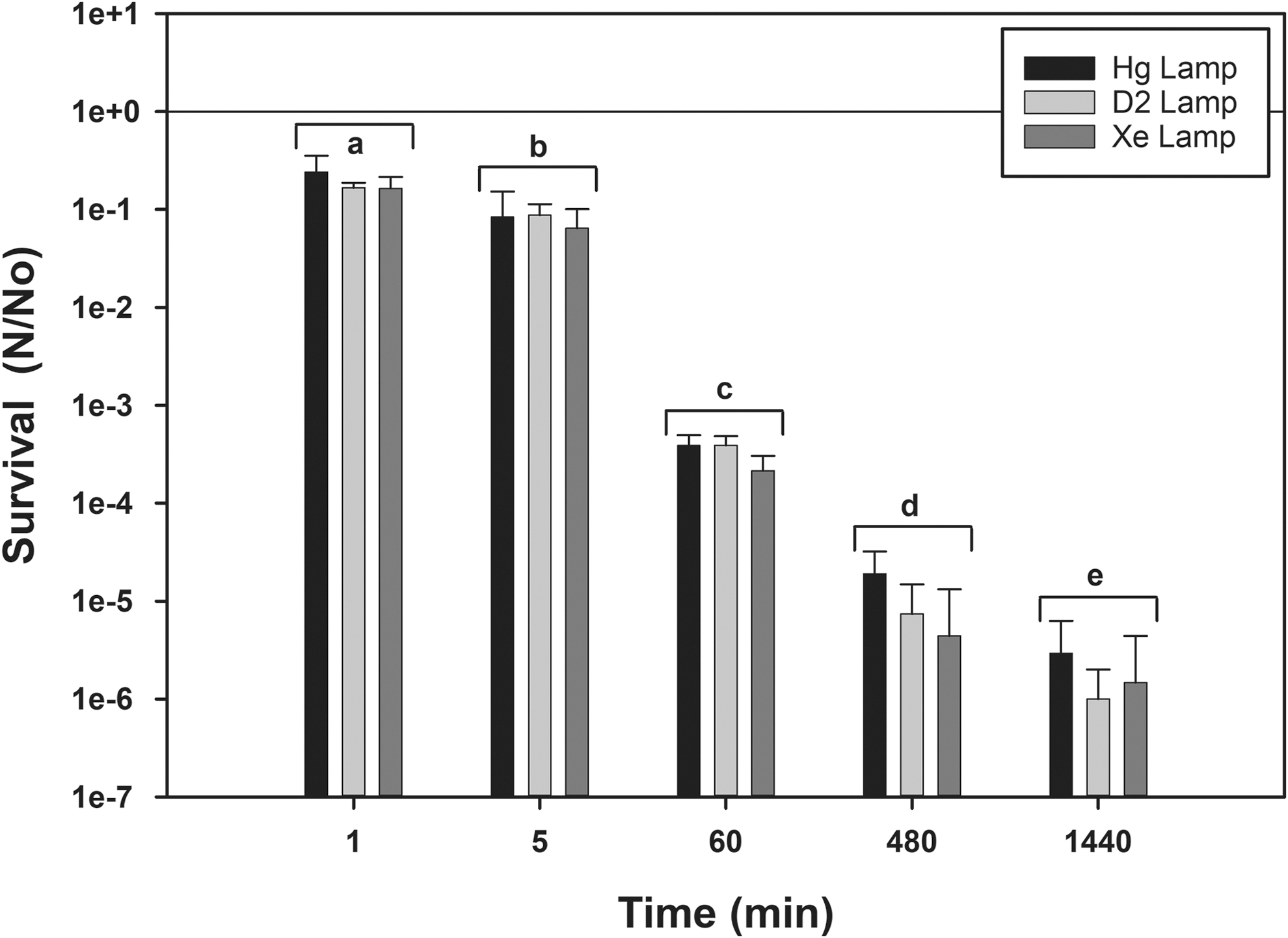
Comparison of the biocidal effects of three UV sources on the survival of Bacillus subtilis HA101 spores. Uncoated aluminum 6061 coupons were doped with 2 × 106 spores per coupon and exposed to UV photons from mercury-line (254 nm), deuterium, or xenon-arc lamps (see text) cross-calibrated to each deliver 3.25 W m−2 of UVC (200–280 nm) illumination. Data were 0.25-power transformed prior to analysis with ANOVA and protected LSmeans tests (P ≤ 0.05; n = 4). There were no differences among the results for each time-step, indicating that each UV source produced the same biocidal effect on the spore monolayers when calibrated to similar fluence rates. However, all UV lamp sources yielded significantly different results over time for the UV exposures; diverse letters indicate significant differences for the clustered results for the three UV lamps.
The experiments described above were expanded to include a total of four Bacillus spp.—B. subtilis HA101, B. subtilis 168, B. atrophaeus ATCC 9372, and B. pumilus SAFR-032—to determine whether the protocol used for B. subtilis HA101 can be extended to other Bacillus species. An experiment to compare the quality of the spore monolayers with SEM imaging for pre-MPN and post-MPN assays was designed in which the coupons extracted with SDIW + silica sand were air-dried and imaged with SEM. Figure 6 depicts the general responses of all four Bacillus spp. to the extraction protocol. First, in most regions of interest (ROIs) imaged by SEM, B. subtilis HA101 and B. atrophaeus 9372 exhibited no spores (data not shown); however, a few ROIs for both species exhibited a few single-digit numbers of spores (e.g., Fig. 6B, 6D). Second, most ROIs of coupons with B. subtilis 168 showed a few single-digit spores to a few small clusters of spores on MPN-extracted coupons (Fig. 6F). Third, ROIs on coupons doped with B. pumilus SAFR-032 spores exhibited small to medium sized clusters of spores (often in depressions), which suggests that the spores of SAFR-032 were affixed to the Class 1A coupons slightly more aggressively than the other three species (Fig. 6H).

Scanning electron microscope images of spores from four Bacillus spp. applied to Chemfilm (Class 1A) treated aluminum coupons processed with MPN assays. For most of the coupon surfaces viewed for B. subtilis HA101 and B. atrophaeus 9372, no spores were observed on the Chemfilm-treated coupons after the MPN assays; however, shown here are examples of when a few spores were observed. In contrast, most viewed SEM regions of interest (ROIs) for B. subtilis 168 and B. pumilus SAFR-032 contained spores. In particular, some of the viewed ROIs for B. pumilus SAFR-032 contained dozens of spores within the 70 × 50 μm ROIs. Magnifications (e.g., × 2.00k) and micrometers (e.g., right side of banners) are given for each image.
When coupons of each Bacillus spp. were processed with the SDIW + silica sand extraction MPN protocol, the quantitative data supported the conclusions outlined above. For example, spores of B. subtilis HA101 and B. atrophaeus 9372 (∼100% of the applied spores were recovered) followed B. subtilis 168 (∼85%), and B. pumilus SAFR-032 (∼60%) were recovered from dried doped-coupons when assayed with the silica-sand MPN protocol (Fig. 7). Furthermore, the quantitative extractions indicated that the lowest number of recovered spores was observed with Class 1A coupons doped with B. pumilus SAFR-032, but still within 1-log of the number of spores initially deposited. Results in Figs. 6 and 7 support the conclusion that spores of different Bacillus spp. can attach to aluminum coupons at slightly different rates; thus, slight differences in recovery rates should be factored into population dynamics on the effects of biocidal factors on spore survival.

Recovery of spores from four Bacillus spp. on Chemfilm-coated (Class 1A) aluminum coupons. The bacteria were Bacillus subtilis HA101, B. subtilis 168, B. atrophaeus 9372, and B. pumilus SAFR-032. Spores were prepared in 40% ethanol and applied to Chemfilm-treated coupons at the rate of approximately 2 × 106 spores per 200 μL microdrop per coupon. The SDIW + silica-sand MPN assay was used to recover spores from coupons. Untransformed data were analyzed with ANOVA and protected LSmeans tests, and similar letters indicated that treatments were not significantly different (P ≤ 0.05; n = 6). Spores of B. subtilis HA101 and B. atrophaeus 9372 were recovered from aluminum coupons at similar rates as were deposited, indicating that nearly 100% of the spores were recovered with the silica-sand extraction process. In contrast, the other two Bacillus spp. exhibited slight reductions on recovered spores, indicating that some spores remained on the coupons after processing. The dashed line indicates the T = 0 control prior to applying spores to coupons.
And lastly, the results of the vacuum-dispersal assays indicate that extremely low levels of dispersed spores occurred at 7 or 0.025 hPa. First, of the numerous replicates of each Bacillus spp. (2 species) in each sample holding device (2 devices), only a single colony of B. subtilis 168 and three colonies of B. pumilus SAFR-032 were observed in 50 cc conical tubes for the 54 coupons assayed (data not shown). Second, when MPN assays were compared among the two Bacillus spp. and two sample holding devices, there were no differences observed among the various treatments (i.e., overall average for 54 coupons was 3.52 × 106 spores per coupon).
Diverse materials have been used to assay the survival of spore-forming and non-spore-forming microorganisms for their resiliency to endure the harsh biocidal conditions of interplanetary space (e.g., glass, aluminum, steel, graphite; Schuerger et al., 2005, 2006; Horneck et al., 2012; Vaishampayan et al., 2012; Wassmann et al., 2012; Panitz et al., 2015; to mention a few). This is both a boon and a problem to modeling microbial survival in outer space. First, the diversity of materials tested provides data on microbial survival on a wide range of species and spacecraft components. In contrast, results among the numerous studies can be difficult to interpret due to wide-ranging survival and recovery rates of the species tested. For example, are the quantitative differences in survival data due to the effects of the space conditions on the microbes directly, on how the microbes attach to or interact with surfaces, or on assay protocols used in the numerous studies? The motivation for the research presented here was to begin a discussion in the planetary protection community on how to standardize protocols to mitigate against interpretive difficulties observed in microbial survival data under various biocidal factors encountered in space.
First, of the numerous materials used on Mars spacecraft, only a few have been imaged with SEM in relevant microbial studies; thus, the general qualities of the surfaces are unknown. For example, uncoated aluminum coupons and Class 1A Chemfilm-treated aluminum coupons were found to be smooth enough to create monolayers of Bacillus spp. spores (Fig. 1A and 1C, respectively). In contrast, the Class 3 Chemfilm-treated aluminum coupons exhibited significant levels of surface defects in which spores could become entrapped and might easily become shielded from UV irradiation (Fig. 1B). Glass and quartz discs are usually the smoothest coupons to use (Edmonds et al., 2009; Levy et al., 2011; Yunker et al., 2011) but may have problems with surface tension with droplets of spore/cell suspensions. All other materials (e.g., aluminum, steel, painted surfaces, anodized, or Chemfilm-treated coupons) typically exhibit surface textures and defects that are likely to impact survival rates of microorganisms (see Schuerger et al., 2005).
Second, a key goal for preparing spores or vegetative cells (henceforth, spores/cells) for the biocidal assays is to create uniform monolayers to avoid the effects of stacking spores/cells in aggregates. Such stacking may not interfere in the biocidal effects of ionizing radiation or heating due to these factors being able to penetrate multiple layers of spores/cells. However, multiple layers of stacked spores have been shown to be capable of attenuating UV irradiation (e.g., Raguse et al., 2016).
The easiest protocol to use to create monolayers of spores is known as the droplet or microdrop technique, in which known densities of spores/cells are applied in small amounts of carrier fluids on the sample coupons (Schuerger et al., 2003, 2005, 2006; Edmonds et al., 2009; Yunker et al., 2011). However, two drawbacks of the droplet technique are (1) consistency and (2) the creation of a coffee-ring effect at the edges of droplets that may create stacked spores/cells (see Levy et al., 2011; Yunker et al., 2011). In response to this problem, airbrush or aerosol spray-deposition devices (Levy et al., 2011; Raguse et al., 2016) and a droplet/filtration/transfer protocol to apply spores to glass coupons (Noell et al., 2013) have been developed. The drawbacks of the airbrush, aerosol, or droplet/filtration/transfer protocols are that they are significantly more complicated than the droplet technique. In addition, for each of the techniques listed above, there are both deposition and recovery efficiencies that must be characterized.
In the current study, a simple droplet technique was used in which smooth monolayers of spores were achieved with four Bacillus spp. (Figs. 2 and 6). The simple protocol was to apply spore suspensions to coupons, allow the spores to settle to the surfaces of the coupons overnight (i.e., monolayers were created in late afternoon and dried early the next morning; approximately 18 h), and then evaporate the carrier fluid—under a sterile air-flow—in an operating biosafety flow hood for 2–4 h the next day. In general, 80% of the created coupons exhibited smooth monolayers of spores with very minor coffee-ring effects (Figs. 2A–2C, 6A, 6C, 6E, and 6G). In contrast, approximately 20% of the created spore layers exhibited multilayered swirls and defects (Fig. 2D–2F) that rendered them unusable for UV irradiation assays but not for assays with ionizing radiation or heat-sterilization protocols. The defective spore layers were attributed to placing the coupons overnight on lab benches that exhibited slight vibrations due to adjacent equipment. These results are very similar to those of Mancinelli and Klovstad (2000) and Schuerger et al. (2003, 2005, 2006) but differ greatly from those of Edmonds et al. (2009) and Raguse et al. (2016), who reported significantly greater clumping in the droplet technique compared to their wet/dry aerosol deposition protocols.
Following the creation of smooth monolayers of spores/cells, how long can they be held in storage before using them in spacecraft survival experiments? In preliminary experiments, spore recovery from doped coupons seemed to decrease with increased time in storage. An experiment was designed to characterize the recovery of spores over 12 weeks and compare it to spore viability in SDIW. Results indicate that spore recovery of B. subtilis HA101 appeared to drop approximately 90% (1 log) over 12 weeks when doped coupons were stored at lab conditions of normal pressure (1013 hPa) and 22–24°C (Fig. 4). Initially, spore recovery was variable during the first 4 weeks (i.e., large standard errors of the means in Fig. 4B) but then grouped around a linear model of declining recovery. In contrast, spore viability from SDIW stocks showed no decline over the same time period (Fig. 4A). No similar data were found in the literature. This experiment highlights the potential for spores to either die over time or become more adhered to spacecraft surfaces than observed with fresh spores and fresh monolayers. It is recommended that spore monolayers should be used no later than 3 weeks after creation to avoid a possible artifact (i.e., experimental error) of declining recovery over time.
Once smooth and uniform monolayers are exposed to experimental space-biocidal factors, extracting the spores/cells efficiently from coupons is essential. Here, the extraction efficiencies of three protocols were compared to T = 0 min controls (Fig. 3). The starting concentrations of spores were approximately 2 × 106 spores per 200 μL droplet per coupon. The monolayers were not exposed to any biocidal factors but were instead assayed with the MPN protocol 24 h after they were dried. Results indicate that adding 1 g of heat-sterilized silica sand to each 50 cc polypropylene tube enhanced the extractions approximately 1 order of magnitude (i.e., henceforth 1 log). In general, using SDIW alone as the extraction fluid resulted in approximately 0.5 to 1 log reduction in recovered spores, which is consistent with the studies by Shuster et al. (2019) and Grand et al. (2011). Furthermore, of the four Bacillus spp. tested for extraction efficiencies from Class 1A Chemfilm–treated aluminum coupons, the most efficient extraction of spores from coupons was noted for B. subtilis HA101 and B. atrophaeus 9372 (∼100% of the applied spores were recovered) followed by B. subtilis 168 (∼85%), and B. pumilus SAFR-032 (∼60%) (Fig. 7). Thus, it appears that divergent Bacillus spp. will exhibit diverse recovery efficiencies, and as such, care must be taken to properly characterize the extraction efficiencies to calibrate the protocol used for any given study. Not accounting for the “stickiness” of the spores/cells might artificially bias results to lower recovery rates that might be interpreted as loss of viability.
Scanning electron microscope images of spore monolayers for the four Bacillus spp. confirmed that smooth monolayers are possible with the droplet protocol (Fig. 6A, 6C, 6E, 6G) and that the order of recovery listed above was most likely due to spores of specific Bacillus spp. remaining on coupons after MPN extractions (e.g., compare pre-MPN to post-MPN SEM images for each species in Fig. 6). For example, B. subtilis HA101 and B. atrophaeus 9372 consistently exhibited approximately 100% recovery in MPN assays (Fig. 7) and exhibited few, if any, spores in most ROIs observed during post-MPN SEM imaging (Fig. 6). In contrast, B. pumilus SAFR-032 exhibited the most imaged spores in post-MPN ROIs (Fig. 6H) and had the lowest number of spores recovered in MPN extractions (Fig. 7). These data support the conclusion that spore stickiness to coupons had a significant effect on accurately recovering spores from monolayers of the four Bacillus spp. tested. Furthermore, because B. subtilis HA101 was consistently recovered at close to 100% efficiency of the applied densities, previous studies with the SDIW +1 g silica sand MPN protocol (e.g., Schuerger et al., 2003, 2005, 2006) can now be considered more reliable in their results compared to studies that used only a SDIW extraction protocol (e.g., Shuster et al., 2019).
The high efficiencies for recovering spores from metal coupons reported here (90–100%) are significantly higher than other protocols like swab, vacuum, and wipe assays (5–40%; e.g., Brown et al., 2007; Estill et al., 2009; Probst et al., 2010, 2011). The primary difference appears to be that the MPN + silica-sand protocol described here was devised to assay doped coupons of various materials used in spore-survival experiments, while the lower efficiency assays listed above were measuring the efficiencies of protocols for sampling spacecraft surfaces. Here, the doped coupons were placed in 50 cc tubes with SDIW and an abrasive, creating ideal conditions to remove spores at close to 100% efficiency. The swab, vacuum, and wipe assays, in contrast, were devised to sample complex surfaces that are much larger than the coupons used here, and in which the sampling area might be on a structural component or payload on an assembled spacecraft. The recovery efficiencies for spore survival experiments (MPN used here) give insights into how biocidal factors degrade and kill bacterial spores, while spacecraft recovery assays (e.g., Brown et al., 2007; Estill et al., 2009; Probst et al., 2010, 2011) give insights on the bioburdens on spacecraft prior to launch. Both approaches are critical to calibrate and standardize in order to predict microbial survival over time on planetary and interplanetary spacecraft.
Given (1) a consistent monolayer preparation protocol, (2) constraining the time between creating and using monolayers to no longer than 3 weeks, and (3) developing an efficient extraction protocol by adding 1 g of sterile silica sand to the MPN assay tubes, a question arose on whether three different UV illumination sources would affect spore survival. Deuterium, 254 nm Hg line, and xenon arc lamps were used to expose smooth monolayers of B. subtilis HA101 to increasing time-steps up to 1440 min (24 h) (Fig. 5). All three UV sources were carefully cross-calibrated to deliver 3.25 W m−2 (195 J m−2 min−1) at the surfaces of monolayers on uncoated aluminum coupons. Results indicate that there were no significant differences among the three UV sources at each time-step (P > 0.05) but that spore survival rates for the UV sources decreased the same amount over time (P ≤ 0.05). Thus, there were no apparent advantages of using one UV light source over the others provided that the fluence rates at the reactive surfaces were carefully calibrated.
The effectiveness and lack of specific effects for the three UV sources allow the direct comparison of survival rates of bacterial species across multiple studies. For example, Mancinelli and Klovstad (2000) and Schuerger et al. (2003, 2005, 2006) used deuterium and xenon arc UV lamps, respectively, for their studies on spore survival of B. subtilis HA101. Results from both studies are in close agreement. In another example, numerous studies have been conducted on the biocidal effects of UV irradiation on a diversity of Bacillus spp. using all three UV lamp types (e.g., Mancinelli and Klovstad, 2000; Nicholson et al., 2000; Setlow, 2001; La Duc et al., 2003; Nicholson and Galeano, 2003; Schuerger et al., 2003, 2005, 2006), the results of which are very consistent among those studies that revealed a general trend of 1-log reductions (LD90) in survival following 20–75 s of UV exposure at the same dosage of ∼200–400 J/m2, regardless of the UV source used.
And lastly, in the experiments on the effects of high vacuum on spore removal from aluminum coupons, results indicate that few if any spores of three Bacillus spp. were dislodged and dispersed by vacuum exposures down to 0.025 hPa. The vacuum assay for the removal of spores was motivated by numerous questions posed by reviewers and conference participants over the last 7–10 years (Schuerger, unpublished). The data presented here need to be confirmed with other species and for longer time-steps than 7 days, but for now suggest that low-pressure experiments with spores applied to aluminum surfaces will not result in the removal of spores from coupons by vacuum. This observation supports the conclusion that loss of spore viability is likely due to the biocidal nature of test parameters and not an artifact of vacuum exposure.
Summary
Developing models for microbial survival in space (e.g., Dillon et al., 1973; Schuerger et al., 2019; Moores and Schuerger, 2020) will depend on microbial survival data that can be directly compared. If experimental protocols and results are too divergent, developing such models becomes significantly more difficult and may require creating new internally consistent data sets instead of using published studies. The experiments presented here are offered to promote conversations in the planetary protection community to develop a standard set of assay protocols that can be used in divergent labs to generate directly comparable results on spore survival against space biocidal factors.
The data presented here demonstrate the following: Smooth monolayers are possible on aluminum coupons with the droplet technique. Adding 1 g of sterile silica sand to individual 50 cc assay tubes enhances spore recovery by approximately 1 log. Fresh spore monolayers are important to achieve high recovery rates from coupons. Vacuum exposure does not dislodge spores from aluminum coupons. If the above procedures are followed, UV inactivation kinetics for diverse species are directly comparable among many studies because there is apparently no effect on spore survival by divergent UV lamp sources.
Future research is encouraged to extend these assays to a wider range of spore-forming and non-spore-forming bacteria, fungi, and archaea. If the results presented here are confirmed with other species, it may be possible to adopt a standardized set of microbial assay protocols for spore survival under key biocidal factors that will allow direct lab-to-lab comparisons. Moreover, such a set of standardized microbial assay protocols for spacecraft will enhance the development and verification of microbial survival models for interplanetary spacecraft.
Conflict of Interest
The author has no financial, commercial, or other relationship conflict of interest with any organization listed in the text.
Footnotes
Author Contributions
ACS envisioned, performed the experiments, wrote, and edited the paper as presented above.
Funding
Funding support for the creation of this work was provided by the National Aeronautics and Space Administration (NASA), Planetary Protection Office, NASA Headquarters, Washington, DC, USA (grants #NNX12AJ84G; 80NSSC17K0263; 80NSSC20K0745) and the Jet Propulsion Laboratory, Pasadena, California, USA (grant #JPL1643079).
Data Management Plan
All raw data for figures are presented in a single Excel file labeled Table S1 in the Supplementary section. In addition, raw data from the current study are available in a University of Florida Institutional Repository (UFIR) for ACS at the link
Supplementary Material
Supplementary Table S1
Abbreviations Used
Associate Editor: Christopher McKay
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.