
Abstract
The concept of habitability is now widely used to describe zones in a solar system in which planets with liquid water can sustain life. Because habitability does not explicitly incorporate the origin of life, this article proposes a new word—urability—which refers to the conditions that allow life to begin. The utility of the word is tested by applying it to combinations of multiple geophysical and geochemical factors that support plausible localized zones that are conducive to the chemical reactions and molecular assembly processes required for the origin of life. The concept of urable worlds, planetary bodies that can sustain an arising of life, is considered for bodies in our own solar system and exoplanets beyond.
1. Introduction
Although much has been discovered regarding the habitability of Mars, an equally important consideration is whether the environments on Mars were just as conducive to the abiotic origin of life itself. Could life have arisen on Mars?
This excerpt from a review by Clark et al. (2021) introduced a separation of the current habitability of Mars from a capacity in its ancient past to host an origin of life. The concept of a planetary habitable zone was first defined by Kasting et al. (1993). Although there are multiple aspects of habitability, the primary property of the habitable zone is a region around a star in which conditions are such that liquid water can be present on a planet for extended periods of time.
Habitability also implies that if a sample of terrestrial soil containing thousands of species of bacteria were to be placed experimentally in the liquid water of a habitable planet, some of the bacteria might survive and even thrive in the alien conditions due to their observed ability to survive in extreme conditions on Earth.
Here, we will propose a new term to describe the possibility that life could not only thrive on habitable planetary bodies but also emerge there. The word that seems appropriate is urability, which incorporates the combining term ur- meaning primitive, original, or earliest. The related adjectives are urable and inurable.
2. Processes and Factors Underlying Urable Worlds
Before we describe the properties of a urable planet that is capable of supporting an origin of life, we will frame a fundamental question: What is a minimal set of physical and chemical conditions, energy sources, and processes required for the emergence of life on a habitable planet? To answer this question, we will focus on the conditions and constraints that were likely to have played a role in the origin of life on Earth. However, our present understanding of prebiotic conditions represents only a subset of the possible conditions that can lead to “urable zones” just as our understanding of habitability represents a subset of possible “habitable zones” in solar systems.
The meaning of urability does not reflect an exact set of conditions, but rather the broader concept that there exists a set of conditions, which, in combination, create a window of opportunity for life to emerge.
What we will present next is a set of geophysical and geochemical factors that characterize locales where prebiotic conditions and processes comprise a urable zone that is conducive to the origin of life. Urable zones are related to the Goldilocks zones that characterize the orbits of habitable worlds (Rampino and Caldeira, 1994) but describe more localized settings on a potentially habitable planetary body where life can emerge. The urability of a given setting is a combination of the individual factors that influence it. As depicted in Fig. 1, a design for a urability graph is proposed in which each factor can be represented as a strip with a range of values along its length.
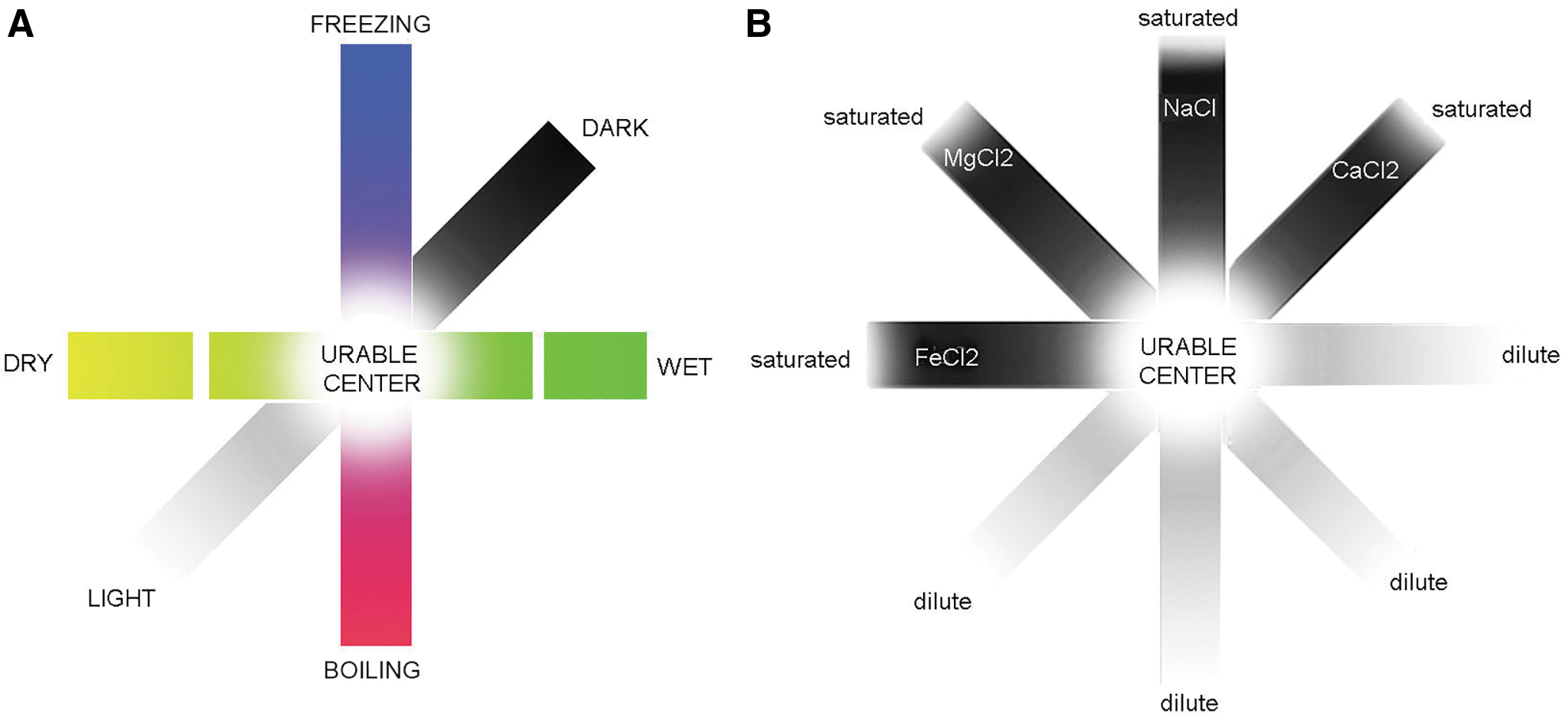
Plausible prebiotic chemical reactions that might be limited on the extreme ends of each factor could become much more likely where the factors overlap to form urable centers. In the first example in the figure, such a center emerges within the fortuitous overlapping of a bounded range of temperature, light energy required to drive photochemical reactions, and water as a solvent. The second example shows how the concentrations of ionic solutes also overlap to form a urable center. We have used just a few factors in the illustration but have identified 28 that constitute a full complement of urable conditions: 12 geophysical, 14 geochemical, and 2 combinatorial.
The size and persistence of urable centers would be highly sensitive to variable conditions that affect each contributing factor. For example, in the early stages of life's origin, a continuous supply of energy and organic compounds would be required. If this supply is interrupted, fragile prebiotic chemical networks would not persist but return to equilibrium. For urable parameters such as mineral inventories, sustained urable centers become more complex as chemical networks are distributed within multiple localized environments.
Urable centers and the corresponding zones in the landscape they can operate within will again be modified when the interaction of energy and organic compounds allows more complex structures called protocells to assemble. When protocells become sufficiently robust to survive environmental stresses and distribution, urable zones will grow in size and diversify as protocells adapt to changing conditions. Urability graphs must correspondingly reflect the many factors and variations that influence protocell evolution through multiple transitions as they enter different settings with new resources and stresses.
3. Specific Planetary Factors Related to Urability
What factors of urability might have predicted that life would begin on the early Earth, other planetary bodies in the solar system, or any of the >1011 exoplanets estimated to exist in our galaxy? (Cassan et al., 2012). It is possible that there are fundamental functions and underlying systems that are universal to all forms of life. A subset of these could not only include the requirement for compartments, homochirality, and various functional polymers that are catalysts but also serve to store and transmit information. The main focus of origins of life research so far has been to understand how these structures and functions emerged on the sterile surface of the prebiotic Earth, and perhaps Mars, ∼4 billion years ago.
As we gain knowledge of the enlarging inventory of the worlds in our solar system and exoplanets beyond and use our growing understanding of how life may have emerged on the early Earth, we can expand our predictions of where life could begin elsewhere. The following list summarizes the parameters related to urability. Some of these can be expressed in relative terms similar to Goldilocks' porridge metaphor of too hot, too cold, or just right, whereas others are simply present or absent.
3.1. Geophysical factors
Size of the planetary body (planet or moon).
Planetary rotation rate and tidal locking.
Presence or absence of a sizeable moon.
Planetary core composition and temperature, and subsequent effect on volcanism and the formation of subaerial landscapes that can interact with the atmosphere and surface liquids.
Hydrological cycles of evaporation and precipitation.
Light energy (infrared—visible—ultraviolet wavelengths).
Magnetosphere providing protection of the atmosphere from solar wind.
Planet-star distance.
Levels of stellar activity.
Tectonic activity.
Crustal mineral inventories: mafic rock versus felsic crust.
Presence or absence of hydrothermal subaerial pools or submarine vents that allow the assembly and distribution of molecular systems.
3.2. Chemical factors
Anoxic atmosphere perhaps with admixtures of reactive gases such as HCN (hydrogen cyanide) and HCHO (formaldehyde).
Liquid water within temperature ranges conducive to sustained prebiotic reactions.
Ionic concentrations ranging from fresh water to salty seas.
Availability of trace element co-factors required for catalytic activity.
Acidity or alkalinity of aqueous solutions.
A continuous source of key organic compounds made available by local synthesis or exogenous delivery.
Synthesis or delivery of specific compounds that are capable of serving as monomers, including amino acids, nucleobases, monosaccharides, and phosphate.
Amphiphilic compounds available for assembly into vesicular boundary membranes.
Sources of energy to drive reactions in a timely manner: chemical energy, redox potentials, light energy, wet-dry cycles, and chemiosmotic energy.
Processes that concentrate dilute solutions of reactants sufficiently to react.
Conditions that capture energy to enable polymerization reactions such that populations of polymers of a sufficient length emerge to support catalytic and information storage functions.
Mixtures of organic compounds capable of being incorporated into systems related to autocatalysis and primitive metabolism.
Selective processes that lead toward homochirality.
Encapsulation processes to enclose sets of polymers and other molecules into populations of protocells.
3.3. Combinatorial factors
Cycling of sets of encapsulated polymers through dynamic environmental stresses to drive the first steps of evolution by combinatorial selection.
Environments capable of supporting selective processes and widespread distribution of self-assembled protocells. Both the environments and protocells must be stable long enough to support the evolutionary transition to living microbial communities.
In Section 4 of this article, we will briefly summarize the relevance and significance of primary urability factors related to the assembly of protocells. Although multiple definitions of protocells have been proposed in the literature, in general they are considered to be a stage on the path toward a living system. Rather than focusing on the properties of individual protocells, it is probably better to think of them in terms of compartmentalized populations of polymers and other components in which (at least initially) each protocell's components are different from all the rest.
During cycles of selection, most will be inert and their components will be recycled, but a few will survive because by chance they have properties and active functions that allow them to withstand environmental stresses. Protocell populations would, therefore, be capable of undergoing selection and evolution toward primitive forms of life.
In Section 5, we will apply the urability concept to two sites proposed to be conducive to the origin of life, and in Section 6 we will consider how the concept of urability can be applied to planetary bodies in the solar system and beyond.
4. From Chemical Evolution to Protocells
4.1. When is a habitable world inurable, and how can it become urable?
A useful perspective addressing the earlier question can be developed through a series of thought experiments. Suppose that we discover a planet in the habitable zone that has an ocean and volcanic land masses, but that is sterile. Now we add a few drops of a mixed culture of terrestrial microorganisms to a small tidal pool. In a few weeks, the pool teems with their descendants and it is clear that we have “infected” a planet that by definition is habitable. However, it may not be urable because life has not originated there. Something is missing from the environment.
We can also perform a laboratory demonstration of habitability in which amino acids, nucleotides, membrane-forming amphiphilic compounds, and trace elements required by life are dissolved in a 1-L flask of water. If the solution is sterilized, filtered, and sealed, it can be stored indefinitely because it is not urable. However, it passes the test of habitability because if a drop of mixed soil bacteria is added, a few days later it will be teeming with microorganisms.
Now let us consider a planetary test of urability. Imagine an Earth-sized planet in orbit around a red dwarf star. The light is in infrared wavelengths longer than 1 μm, just enough to warm the atmosphere but insufficient to drive photochemical reactions. The planet is gravitationally locked, does not rotate, and lacks a moon and tidal effects. It has no land masses but is covered by a salty global ocean several kilometers deep that contains millimolar concentrations of amino acids, nucleobases, simple carbohydrates, phosphate, and other biologically relevant solutes. There is also a mixture of oily hydrocarbons and long chain monocarboxylic acids present as an oil slick on the surface (Lasagah et al., 1971; Nilson, 2002).
The organic compounds were supplied during accretion by the infall of interplanetary dust particles with smaller amounts delivered by carbonaceous meteorites and comets. The atmosphere is a mixture of nitrogen and carbon dioxide, and the planet's deep interior is warmed by radioactive decay but only to 300K so that there is no magma and hence a lack of volcanic or hydrothermal activity.
Is this a urable planet? Could life begin in its ocean? The answer, based on what we know about terrestrial life, is no. There is no energy source available to drive chemical reactions because the global ocean is composed of soluble compounds and salts present as a dilute solution at thermodynamic equilibrium. Now we come to the second step in the thought experiment. What parameters in the above list can be added to make the planet urable? Let us first consider the list of geophysical parameters. For instance, liquid water is a property of habitable planets, so the planet must have sufficient gravity to maintain liquid water for several billion years on its surface.
Earth and Mars are contrasting examples in the solar system. Although early Mars had a geomagnetic field and atmosphere sufficient to support liquid water at the surface, the size and gravity of Mars were insufficient to maintain the original water for more than a few 100 million years, perhaps longer (Carr and Head, 2010), whereafter the atmosphere escaped and the pressure-temperature regime at the surface only permitted transient liquid water. In contrast, Earth has maintained an ocean for more than 4 billion years, largely due to increased gravity related to its mass combined with a metallic core that produces a geodynamo and drives a magnetosphere. It is reasonable to conclude that Earth-sized planets can be urable for extended periods whereas Mars-sized planets may only be urable for a limited period of time.
Instead of being gravitationally locked, we could add rotation to the planet in the thought experiment but it is not apparent why this might significantly enhance urability. Adding a moon to a rotating planet will produce a modest tidal bulge in a global sea but would not add a source of chemical energy. Finally, we can bring the wavelength of light down from the infrared into the visible range. One might imagine that this would provide an energy source to drive photochemical reactions but in the absence of pigment molecules that can capture and transduce the light energy, photosynthesis or other light energy harvesting processes would not begin.
We have now exhausted all but one of the geophysical factors, which is to add sufficient quantities of long-lived radioactive isotopes such as thorium 232 and uranium 235 and 238 to the planet's core so that the temperature increases to that of the Earth's core and mantle, around 6000K (Nimmo et al., 2020). At this temperature, the silicate minerals of the mantle surrounding the core are melted and molten plumes begin to rise and break through the crust, producing volcanic islands resembling Iceland and Hawaii.
Volcanism is a significant source of energy in settings for origin-of-life processes that can drive organic compounds away from equilibrium and build molecular complexity. Complexity also implies diversity in molecular structures, and in a dynamic volcanic landscape environment, can be the stage on which chemical selection processes occur; a key process to facilitate an origin of life and is thus listed as a combinatorial urable factor above. Urability strongly depends on one or more sources of chemical energy, and volcanism facilitates this as well as creating a urable landscape that is both chemically complex and diverse.
There is one geochemical factor that can also provide chemical energy. If the submarine crust happens to have extensive deposits of olivine, which seems likely, exposure to seawater allows a chemical reaction called serpentinization. This reaction produces strongly alkaline solutions enriched in dissolved hydrogen gas as well as columns of calcium carbonate minerals such as those observed at Lost City (Kelley et al., 2001). However, although such systems exist for a limited time away from equilibrium, to serve as a factor of urability there must be a way for the energy be captured by the synthesis of more complex organic compounds such as polymers, a possibility that will be discussed in Section 5.
4.2. Properties of liquid water related to urability
We can use the variable properties of liquid water on Earth as a guide to its physical and chemical parameters contributing to urability. Temperature, of course, immediately comes to mind. Water is a liquid near 0°C and hydrothermal water associated with volcanic activity can reach boiling temperatures near 100°C, depending on altitude and atmospheric pressure. Various species of microbial life have adapted to thrive throughout the range at which water is a liquid, so temperature is not a major limitation of habitability.
However, extreme temperatures can markedly affect urability. Temperatures near 0°C may not provide sufficient thermal energy to overcome the activation energy barrier and allow chemical reactions to occur, whereas temperatures near boiling can strongly affect self-assembly processes such as membrane stability and hydrogen bonding required for the functions related to the double helix of DNA and folding of RNA into functional structures such as ribozymes. It is reasonable to assume that temperatures in the mid-range are optimal for an origin of life on Earth or elsewhere.
Another property of water is related to the pH of solutions. The acidity or alkalinity of water has two important aspects. Extremely low or high pH catalyze the hydrolysis of the covalent ester and peptide bonds that link monomers into biopolymers, so it seems unlikely that life could begin in extremely alkaline or acidic water. A more conducive pH to support life's emergence is in the range from moderately acidic (pH 3) to slightly alkaline (pH 8) (Deamer et al., 2019). Second, pH strongly affects the chemical properties of monomers with titratable moieties such as the carboxylate and amine groups of amino acids and the phosphate groups of nucleotides.
The presence or absence of a negative or positive charge on a potential monomer dramatically affects its ability to engage in polymerization reactions. It also affects various biophysical self-assembly processes, such as the ability of amphiphilic compounds to form membranes and the hydrogen bonding that stabilizes protein folding and base pairing of nucleic acids.
We also need to consider the soluble salts of Na, K, Mg, Ca, and Fe, such as chlorides and sulfates, as well as phosphates that are essential to bioenergetic metabolism. In terrestrial life, all these ionic species are essential, but there are limitations imposed by physical and chemical effects. For instance, in the slightly alkaline pH of today's ocean (pH 8.1), calcium strongly binds to phosphate groups in solution and precipitates as the insoluble mineral called apatite, thereby removing phosphate as a soluble species.
The high concentration of sodium chloride in terrestrial seawater also causes osmotic effects that reduce the stability of membranous compartments (Black et al., 2013; Milshteyn et al., 2018). Life has evolved mechanisms in the form of ion transport enzymes to adapt to salty seawater, but primitive life would not have such complex systems. For this reason, we and others have argued that a freshwater environment would be more conducive to urability and the origin of cellular life on the early Earth (Mulkidjanian et al., 2012; Damer and Deamer, 2015, 2020; Van Kranendonk et al., 2021).
4.3. Sources of organic compounds
It is given that a source of organic compounds is essential for life to emerge on a habitable planet. One possible source is the chemical synthesis of organic compounds in the atmosphere, as originally proposed by Miller and Urey (see McCollom, 2013 for review) and more recently by Pearce et al. (2020). An alternative is the synthesis at hydrothermal vents suggested by Martin and Russell (2007), Lane and Martin (2012), and Russell (2021). The delivery of reduced organic compounds by extraterrestrial infall during late accretion is another possibility.
Chyba and Sagan (1992) and Flynn et al. (2004) reviewed such sources and concluded that organics delivered by the continuous infall of interplanetary dust particles would be by far the major source 4 billion years ago, delivering 108 kg/year with much less delivered by carbonaceous meteorites and comets. Most of the organic compounds would fall into the ocean and become too dilute to take part in chemical evolution. However, assuming that volcanic land masses resembling Hawaii and Iceland had emerged from the global ocean, the organic compounds would accumulate on mineral surfaces and then be further concentrated when precipitation flushed them into small pools.
Pearce et al. (2017) used a computational approach to estimate how long a delivered molecule such as adenine would be available in such pools. This is a random process so the distribution would not be uniform, but it seems likely that the organic compounds would accumulate in small pools that were exposed to wet and dry conditions by cycles of evaporation and precipitation.
It has always been assumed that a variety of organic compounds would have been synthesized by Miller-Urey or Fischer-Tropsch reactions (Lazcano and Bada, 2003), but lacking direct evidence confirming that this occurred it remains as an inference from laboratory simulations. On the other hand, extraterrestrial delivery is reasonably certain because it is still happening today, although at much reduced rates. Which source is primary may not be a significant issue.
Any organic compounds that were present in the late Hadean and early Archean Earth, either synthesized or delivered, would concentrate in pools, mix, and undergo further reactions that would drive the mixture of solutes in hydrothermal pools toward a similar composition. Some of the products such as polymers represent increases in chemical complexity that clearly contribute to the urability of that particular pool.
4.4. Amphiphiles and encapsulation
All life today requires compartments, so it seems plausible that the first forms of cellular terrestrial life had boundary structures composed of simple amphiphilic molecules such as fatty acids, which underwent chemical selection to become the highly evolved phospholipid membranes used by modern organisms. There are two possible sources of amphiphiles. Carbonaceous chondrites such as the Murchison meteorite contain amphiphilic compounds, including monocarboxylic acids, which represent half of the soluble organic compounds in the meteorite (Huang et al., 2005). The total amount present is ∼500 ppm relative to the total mass of the meteorite, and the molecules range up to 13 carbons in length.
Monocarboxylic acids having hydrocarbon chains with lengths of 10 or more carbons are long enough to assemble into lipid bilayer membranes at ordinary temperatures (Monnard and Deamer, 2002). Amphiphilic compounds present in the Murchison meteorite can also form membranous vesicles (Deamer and Pashley, 1989).
The second potential source is Fischer-Tropsch synthesis in geothermal conditions, which are known to produce a mixture of monocarboxylic acids and alcohols (Nooner et al., 1976; Simoneit, 2004). It is not clear which might be the main source but the fact that amphiphiles were present in carbonaceous meteorites delivered to the early Earth suggests that they are likely to have been available in the prebiotic environment. Other structures such as mineral pores and coacervates were less likely to serve as compartments, because they lack boundary membranes that can function as permeability barriers to diffusion of ionic solutes.
The fact that all life today is cellular with boundary membranes composed of amphiphilic compounds supports the conclusion that the first forms of primitive life were also cells. It follows that the encapsulation of polymers by membranous compartments was a major urability factor on a habitable planet.
4.5. Sources of energy and concentration processes
Even if multiple organic compounds are present as dilute solutions in liquid water, not much would happen unless sources of energy impinged on the mixture. Only a few primary energy sources would have been available on the prebiotic Earth.
Light energy would be abundant as it is today, but two factors are required for it to become a useful source: light-absorbing molecules capable of capturing the energy and transitioning from a ground state to an excited state, and a mechanism by which the photochemically activated compound can couple the energy of its excited state to the synthesis of stable biologically relevant molecules. Life today depends on chlorophyll as a pigment, and the stable products that store the energy are carbohydrates and hydrocarbon derivatives. However, the question of which systems would be capable of performing similar photochemical reactions in early life remains to be answered.
A second source is chemical energy, with a good example being the pyrophosphate bond in adenosine triphosphate (ATP) that plays an essential role in metabolism and most other cellular functions. Holm and Baltscheffsky (2011) proposed that pyrophosphate itself could have served as an energy currency in early life, but a mechanism by which high energy bonds could be continuously synthesized and coupled to primitive metabolic systems is still an open question. Significant chemical energy is also made available in conditions that mix dissolved gases such as hydrogen and carbon dioxide, where hydrogen provides reducing power that can donate electrons to CO2.
The products can include formic acid and formaldehyde (Herschy et al., 2014), which have been proposed to represent the initial steps of primitive metabolism (Lane and Martin, 2012).
Another version of chemical energy is made available by electron transfer reactions, which are fundamental bioenergetic processes in all life today. Bacteria, mitochondria, and chloroplasts have molecular systems called electron transport chains composed of proteins embedded in membranes. The electron transport chains have two functions. The first is to capture redox energy by transporting electrons from a source to a sink, and the second is to generate proton gradients across membranes.
This brings us to a third energy source called chemiosmosis (Mitchell and Moyle, 1967). Mitchell's aim was to explain how ATP can be synthesized by bacteria, chloroplasts, and mitochondria. He proposed that if an enzyme capable of hydrolyzing ATP was embedded in a membrane and served as a proton pump, the reaction could be driven in reverse if another process produced a proton gradient across the membrane. Instead of hydrolyzing ATP, the enzyme would synthesize ATP from adenosine diphosphate and inorganic phosphate. There was significant initial skepticism about chemiosmosis, but as supporting evidence accumulated the scientific community became convinced that it was correct.
Could chemiosmosis serve as an energy source for the first forms of life? Koch (1985) first suggested this possibility, and Lane and Martin (2012) extended it to the mineral matrices of alkaline hydrothermal vents. In our view, there are two hurdles to overcome. First, there must be a microscopic compartment that is bounded by a lipid bilayer membrane, and the membrane must represent a significant barrier to proton permeation so that it can maintain a proton gradient. There must also be a mechanism by which a redox potential can be coupled to the generation of a proton gradient. Two components of electron transport chains in chloroplasts and mitochondria are quinone compounds called plastoquinone and ubiquinone.
These are small mobile molecules embedded in membranes that can accept electrons and protons on one side of the membrane and transfer them to an acceptor on the other side. When quinones are reduced they must also accept hydrogen ions (protons) from water, but the receptor on the other side of the membrane may not require hydrogen ions so they are released. This process produces a proton gradient across the membrane that has been measured experimentally in bacteria, mitochondria, and chloroplasts.
A recent study (Milshteyn et al., 2019) demonstrated that quinone compounds in the Murchison carbonaceous meteorite are capable of generating proton gradients in a model membrane system. In principle, the gradient could provide a source of chemiosmotic energy, but there is not yet a system equivalent to an ATP synthase that could couple the energy of the gradient to ATP synthesis. Until such a coupling mechanism can be experimentally demonstrated, chemiosmotic energy transduction seems more likely to be a later development associated with the evolutionary emergence of microbial life.
4.6. Wet-dry cycles as an energy source for polymerization
For Earth-like worlds sharing similar prebiotic chemical and physical environments, a minimal requirement for the origin of life is a nonenzymatic process by which catalytic and replicating polymers are synthesized under plausible prebiotic conditions. This brings us to the last source of energy to be described, a source that is the simplest in concept but largely ignored, perhaps because it is not used by life today. We will begin with a fundamental equation of thermodynamics: ΔG = ΔH − TΔS, where ΔG is the Gibbs free energy available to drive a chemical reaction, ΔH is the enthalpy of the reaction, T is the absolute temperature in degrees Kelvin, and ΔS is the change in entropy of the reaction. If ΔG is negative, the reaction will be spontaneous and can proceed from reactants to products. A negative ΔG is, therefore, an essential property of any chemical reaction proposed to be involved in the urability of a habitable planetary body.
The condensation reaction to be described requires monomers such as amino acids or nucleotides to be sufficiently concentrated to polymerize into peptides and oligonucleotides. This process is called a condensation reaction, because it involves the removal of water from between monomers to form peptide bonds or ester bonds. All life today depends on polymers, so it is important to understand how condensation reactions might have synthesized polymers on the prebiotic Earth. The basic reaction is monomers polymers + H2O, which requires ΔG to be negative.
The term ΔH − TΔS must therefore also be negative, and this can occur if either or both ΔH and ΔS can change sufficiently in a favorable direction. When a dilute solution of monomers undergoes evaporation, the monomers become increasingly concentrated, which represents a favorable change in ΔS. As the solution approaches dryness, the activity of water decreases and produces a favorable change in the enthalpy term ΔH. The result is that condensation reactions and polymerization can occur when a dilute solution of monomers undergoes wet-dry cycles at an elevated temperature so that activation energy is available. Under these conditions, polymers can rapidly form in the concentrated film of the dry phase of the cycle (Ross and Deamer, 2016). This principle has been demonstrated in the laboratory by Rajamani et al. (2008), Forsythe et al. (2015), and Rodriguez-Garcia et al. (2015).
4.7. Summary of urable processing up to protocells
Powner and Sutherland (2011) introduced the concept of “Goldilocks chemistry” applied to geochemical settings that are capable of sustaining certain key prebiotic reactions. We argue here that individual chemical reactions occurring on a habitable planet are necessary but not sufficient to support the emergence of cellular life. Urability requires a series of interlinked and progressive stages at micro- and macro-scales in the landscape. Such stages could be equated to a fabrication process on a factory assembly line in which potentially functional components are added in a stepwise manner, and they are then tested at each stage. In the case of protocells, the components being synthesized are initially random polymers encapsulated in membranous compartments, and testing is performed by selection and amplification of increasingly robust and functional polymer assemblages.
We can now impose constraints that will guide initial experimental tests of urability. For instance, some of the polymers that are synthesized must attain a minimal length that is capable of folding into configurations that have catalytic properties. Other polymers must be able to contain genetic information in sequences of their monomers, and there must also be a process by which that information can be duplicated. Encapsulation of polymers in membranous compartments to form protocells is essential. The compartments must be small enough to enable crowding, thereby providing a concentrated medium that promotes chemical interactions; however, they should be sufficiently pliable to allow budding and division during the reproduction of encapsulated sets of polymers.
At the macro-scale, protocell populations would begin to evolve toward primitive cellular life when initially random sets of encapsulated polymers, each representing a natural experiment, were exposed to environmental stresses and combinatorial selection that preceded Darwinian natural selection of encoded heritable traits. For selective hurdles to work, a urable setting such as a small pool cannot be at equilibrium but instead must be exposed to regular inflows of energy and potential nutrients, and then they should be stirred by physical and chemical cycling. This is why cycling is part of the process.
It has the effect of “pumping” the system so that increasingly complex polymers accumulate in a kinetic trap and reach a steady state in which encapsulated combinations have a chance to exhibit biologically relevant properties. These can be as simple as the stabilization of otherwise fragile lipid membranes or pore formation that allows specific solutes to have access to the interior volume of a membrane-bounded compartment. More complex functions include catalysis and information storage, but these remain to be demonstrated in future research (Damer and Deamer, 2020).
5. Direct Comparison of Urability Factors Applied to an Origin of Life at Submarine Alkaline Hydrothermal Vents and Freshwater Hot Spring Pools
5.1. Urability of hydrothermal vents
To illustrate the factors that influence urability, we will compare two hydrothermal sites that have been proposed to be conducive for the origin of life on Earth. More than 99% of the liquid water on Earth is in the ocean, so a common assumption is that life began in seawater. The discovery of high temperature “black smokers” provided a potential energy source and was soon proposed to be a potential site for the origin of life (Corliss et al., 1979, 1981; Baross and Hoffman, 1985). However, when lower temperature alkaline hydrothermal vents were later discovered (Kelley et al., 2001), they became a preferred site. Russell (2021) recently provided an extensive summary of arguments as to why alkaline vents are the most plausible site for life's origin.
Several physical and chemical factors are related to the potential urability of vent environments. The formation of black smokers requires a significant source of magmatic heat to dissolve minerals as seawater percolates through rock formations beneath the sea floor, whereas alkaline hydrothermal vents require the chemical reaction of serpentinization in which seawater reacts with olivine minerals and releases hydrogen gas and strongly alkaline fluid. It is possible that hydrogen gas could serve as a source of reducing power that reduces carbon dioxide to biologically relevant compounds. Herschy et al. (2014) and Sojo et al. (2019) tested this possibility in laboratory simulations of alkaline hydrothermal vents and observed small amounts of formic acid apparently synthesized from dissolved carbon dioxide.
The interface between strongly alkaline fluids percolating through the porous vent minerals and slightly acidic seawater results in a pH gradient that could provide another energy source. Hudson et al. (2020) tested this idea in a laboratory simulation in which hydrogen gas and carbon dioxide were added to simulated seawater at 1.5 bars of pressure. Fluid at a higher alkaline pH was introduced into a porous mineral composed of Fe(Ni)S precipitates that was submerged in seawater at a more acidic pH. Using mass spectrometry, the authors observed a synthesis of ∼1.5 μM formic acid synthesized from the dissolved carbon dioxide.
The authors noted the similarity between these results and the Wood–Ljungdahl acetyl-CoA pathway used by cells today. They suggested that carbon fixation reactions such as this could have served to initiate primitive metabolic pathways in the vent environment. Compartmentalization in the vent environment has also been proposed to occur within mineral pores (Baaske et al., 2007) or gels (Westall et al., 2018). The organic products synthesized deeper in the vent environment could migrate into these concentrating compartments, possibly subjecting them to cycles of polymerization.
Longo and Damer (2020) summarized the stages underlying an origin of life in the hydrothermal vent environment: Simple molecules such as carbon dioxide, hydrogen sulfide, and molecular hydrogen bind to mineral surfaces and undergo reactions that synthesize more complex biologically relevant organic molecules such as amino acids and nucleotides. These have the potential to polymerize by condensation reactions on the surfaces of mineral compartments or within mineral gels. Circulation through pore openings selects for longer polymers that become encapsulated within amphiphilic lipid membranes to form protocells. As the protocells move vertically up the vent chimney, they are stressed by decreasing thermal energy and decreasing pH, promoting the evolution of increasingly resilient and robust populations. Finally, protocells would be released into the bulk of the ocean and adapt to conditions beyond the vents.
Because experimental investigations of the hydrothermal vent scenario have only advanced to stage 1 above, the urability of the vent environment to support subsequent steps has not yet been established.
5.2. Urability factors related to subaerial hot springs
An alternative to hydrothermal vents begins with water evaporating from salty seawater and falling as fresh water on volcanic land masses, where it accumulates in hot springs (Damer and Deamer, 2020). In contrast to hydrothermal vents, hot spring water undergoes continuous processing by wet-dry cycles that provide a source of free energy to drive condensation reactions. When solutions of amino acids or mononucleotides are exposed to multiple wet-dry cycles in the laboratory, the monomers undergo condensation reactions that produce oligopeptides linked by peptide bonds (Forsythe et al., 2015; Rodriguez-Garcia et al., 2015; Campbell et al., 2019) and mononucleotides linked by phosphoester bonds (Rajamani et al., 2008; DaSilva et al., 2014; DeGuzman et al., 2014).
Further, if amphiphilic compounds are present in the mixture, the polymers become encapsulated in membranous compartments to form protocells (Damer and Deamer, 2020). An origin of life in subaerial hot springs would, therefore, incorporate the following stages:
Collecting and concentrating exogenously sourced or locally synthesized organic compounds that can serve as monomers or components of self-assembled membranes.
Self-assembly of membranous compartments from amphiphilic compounds.
Condensation reactions that drive monomers to form polymers that accumulate in a kinetic trap.
A process by which polymers can be encapsulated in membranous compartments to form protocells.
Cycling protocells through physical processes that select for increasingly complex and robust systems of polymers expressing catalytic and informational functions, forming the earliest systems of biology.
Distribution of populations of protocells within networks of interconnected pools in a hydrothermal field and downhill to other locales in the landscape.
Given a source of monomers, steps 1–4 have been demonstrated in the laboratory. For step 5, mixtures of reactants must be in small pools subject to periodic, short-term cycles of hydration and drying. Repeated cycling is necessary to drive molecular evolution, which increases the robustness of protocells and their polymer cargoes (Da Silva et al., 2014). Distribution of robust protocell populations (step 6) would occur as water circulates through interconnected pools or by wind-driven transport of dry films containing protocells. Because four of the six steps listed earlier have been experimentally demonstrated, we consider subaerial hot springs to be examples of plausible urable zones.
One concern about hot springs in relation to the origin of life is that small bodies of water might be short-lived, which introduces a potential vulnerability for living systems to emerge and persist. However, this weakness is compensated by another property, the capacity of the products of urable processes such as protocells to be distributed among networks of multiple pools. The novel urable factor of robust distribution is expressed as an expanding zone of urability that allows increasingly complex systems to be distributed across diverse settings. Therefore, constraints placed on some urable zones, such as limited lifetimes, can be overcome by distribution to other zones.
Beyond the survival benefits accruing to dispersed protocells, a urable landscape can also be defined as a complexity landscape that is chemically and combinatorially diverse (Van Kranendonk et al., 2021). A hydrothermal field of interconnected pools would be able to subject protocell populations to various resources and selective barriers, thereby providing distribution and recombination of molecular innovations. The urable processes surrounding distribution are maintained as an operating principle of biology, exemplified by the fact that all life today actively distributes progeny by microbial motility, by seeds and spores of plants and fungi, and by animal locomotion.
5.3. Chemical and energetic selective stresses driving the evolution of protocell populations
Sections 4 and 5 described the physical and chemical properties that promote a process by which a mixture of small organic molecules will spontaneously increase in complexity up to the point that vast numbers of protocells are produced. These reactions have been demonstrated in laboratory simulations, and protocell model systems are available for further investigation (Budin et al., 2012; Adamala and Szostak, 2013). However, a urable planet will not only have properties that allow the assembly of primitive forms of life, but also another set of properties including selective stresses that promote prebiotic combinatorial selection and continued evolution after life emerges.
This stage represents the edge of scientific progress; so rather than speculating how evolution beyond protocells might be possible, we will list the representative environmental conditions that provide both amplifying (positive) stresses and negative selective barriers that can guide future investigations.
5.4. Positive selective stresses
Availability of pools with diverse compositions of ionic solutes and pH ranges.
Consistent sources of acidic pH caused by dissolved gases such as sulfur dioxide and carbon dioxide.
Nutrient availability to support metabolism.
Trace elements such as iron in solution to serve as cofactors in catalytic polymers.
Pigments to capture light energy and establish a primitive version of photosynthesis.
A mechanism that allows the replication and sharing of functional polymers encapsulated in protocells.
Processes and conditions that enable protocells to form aggregates of gel-like structures enabling the first niche construction, group selection, and networked interactions (Damer, 2016).
5.5. Negative selective stresses
Osmotic stresses imposed by exposure to varying concentrations of ionic solutes.
Temperature extremes that affect the permeability of membranes and promote the denaturation of essential polymers.
pH extremes leading to the hydrolysis of ester and peptide bonds.
Divalent cations that interact with carboxylate and phosphate groups to cause precipitation into insoluble aggregates.
Ultraviolet light damage to organic compounds that absorb those wavelengths.
Disruptive shear forces occurring in turbulent water that affect fragile prebiotic membrane stability.
6. Application of Urability to Planets and Moons of the Solar System and Exoplanets
We can apply the concept of urability to other planets and moons to evaluate the conditions that might support an origin of life. Perhaps cellular structures and molecular functions quite similar to those on life on Earth would also emerge on planets that are also urable, but there may be other possible outcomes, including “weird life,” which utilizes quite alien molecular materials and mechanisms (Toomey, 2013). Understanding the parameters that affect how life could begin and evolve on similar but also very different worlds will be central to the decisions to be made as we search for life elsewhere in our solar system and beyond.
6.1. Earth-like worlds: Mars
Mars resembles the Earth in terms of size, rocky composition, and the presence of liquid water early in its history. It is interesting to project what we know about the origin of life on Earth to Mars, beginning with positive urability factors. For instance, both planets have evidence of volcanic landforms and liquid water seas early in their histories, with similar availability of light energy, anoxic atmospheres, and organic compounds either delivered during late accretion or synthesized by atmospheric or geochemical reactions.
In addition, both planets had a dynamic surface environment with hydrological cycles that involved evaporation and precipitation. Given these factors, it is reasonable to think that Mars could have supported an origin and evolution of life reaching some stage of development of microbial communities.
Over the past 4 billion years, however, the two planets have diverged in terms of their surface conditions. The surface of Mars lost its habitability as its atmosphere and water evaporated into space, followed by the surface becoming highly oxidized and exposed to ultraviolet radiation. This does not exclude the possibility that a martian biosphere survived, retreated underground, and dwells today within the refuge of subterranean brine environments and hydrothermal plumbing systems. Such life would harness chemical energy by metabolic pathways similar to those used by lithotrophic microorganisms on Earth today.
There are other urability factors to consider. A key geochemical factor is that Mars could have had different relative abundances of cations in solution (Mg, Ca, Fe) that would affect the assembly of protocells. There is extensive evidence of evaporites and salty brine remnants on Mars that are linked to the loss of surface water (Sriaporn et al., 2020). Similar evaporative brines on the Earth limit microbial life to just a few halophilic species that have evolved mechanisms to protect against saturated salt concentrations.
The composition and properties of brines depend on the atmosphere and source rocks. For instance, volcanism, acidic atmospheric conditions, and mafic crustal composition have led to widespread magnesium sulfate brines ∼3.7–2.9 Ga on Mars (Moore et al., 2010). This is another significant geochemical factor, a selective barrier on early Mars that would limit the emergence of protocells.
On the other hand, the problem of high ionic solute content becomes less significant when we take into account the possible subaerial hydrothermal systems on Mars. Even if high salt concentrations limited the urability of seawater, evaporation from the seas would fall on volcanic land masses as distilled precipitation, thereby supplying freshwater for hot springs. The freshwater pools would undergo continuous wet-dry cycles as they do on Earth today, thereby concentrating potential organic reactants and driving condensation reactions to produce biologically relevant polymers. Significantly, such pools tend to be moderately acidic due to the acidic gases carbon dioxide and sulfur dioxide that are commonly present in volcanic outgassing. Acidic pH ranges protonate phosphate groups and thereby promote condensation reactions that synthesize ester bonds, such as those linking the monomers of nucleic acids.
We conclude that the surface of Mars was transiently urable in its early history, so life may have emerged independently on both Mars and the Earth. It is well worth searching for preserved biosignatures to test this possibility, but the smaller size of the planet, loss of the magnetosphere, and atmospheric escape would have rapidly reduced the urable zone on the surface of Mars when compared with the Earth and its global ocean (Longo and Damer, 2020).
6.2. Icy moons: Europa and Enceladus
Jupiter's moon Europa and Saturn's moon Enceladus are ocean worlds surrounded by multi-kilometer thick ice shells. Lacking atmospheres, their surfaces are exposed to radiation that would be sterilizing. Liquid water is buried tens of kilometers deep beneath the ice, so the ability to harvest light energy could not evolve. However, it is possible that hydrothermal vents are present at the ocean-rock interface of Enceladus and Europa, which possess some urable properties, particularly organic molecule synthesis and geochemical energy (McKay et al., 2008).
However, if an origin of life requires dynamic exposed surfaces, a mechanism for polymer synthesis, membrane self-assembly, and selective barriers for evolution, neither Europa nor Enceladus is urable, yet they may be habitable based on our current definition of habitability. It is conceivable that at some point in their early history microbial life adapted to energy sources of hydrothermal vents could have been delivered from other planets. Even if the icy moons are not urable, they may be habitable; thus, searching for biosignatures in the plumes emerging from the south pole of Enceladus, for example, is a worthwhile goal.
6.3. Cryogenic hydrocarbon worlds: Titan
Titan is a sizable moon that undergoes long seasonal transitions as it orbits around Saturn. Its axis of rotation and orbit are stabilized by Saturn, tidally locking one side of the moon to Saturn. Titan is characterized by a thick hydrocarbon-rich atmosphere and cryogenic temperatures. The low temperature range allows liquid methane and ethane to condense into lakes with a landscape of ices composed of water and nitrogen. Despite the chemical differences, the observed surface structures are analogous to geologic features on Earth, including dunes, mountains, rivers, lakes, and even cryovolcanoes (Lopes et al., 2007).
These features clearly indicate the presence of a dynamic surface environment that is capable of facilitating complex chemical interactions, including transport and concentrating mechanisms. The atmosphere undergoes chemical evolution today and likely in the past (Clarke and Ferris, 1997), and the organics go through a process of synthesis and cycling from different levels in the atmosphere to the surface and back again, through evaporation and atmospheric transport mechanisms.
The atmosphere is anoxic and generates complex organic molecules that accumulate as tholins in the lower atmosphere and then rain down to coat the surface of Titan. Tholins at ordinary temperatures are viscous fluids. When exposed to water, tholins form what appears to be compartments, some of which break away to form spherical cell-sized structures (Kawai et al., 2013). These are composed of carbon, nitrogen, and hydrogen and are referred to as azotosomes. Stevenson et al. (2015) proposed that azotosomes could serve as compartments similar to the membranous compartments that assemble from amphiphilic compounds.
Titan exhibits a very different set of environmental properties than Earth-like worlds and there are several primary physical and chemical barriers that reduce Titan's habitability and also its urability. In particular, the thermodynamic stability of complex organic molecules at the surface has been argued to be unfavorable (Sandström and Rahm, 2020). In addition, thermal activation energy is virtually absent in the cryogenic temperature of liquid methane and ethane, so it would be necessary for other mechanisms such as photochemical energy in the upper atmosphere to activate polymerization.
Although Titan might support an unknown form of life that does not require liquid water, the physical and chemical conditions fall outside the factors we are using to define habitability and urability.
6.4. Exoplanets
In regard to exoplanets, they are clearly the next frontier of searching for evidence of life over an enormously diverse range of potentially habitable and urable worlds. Although we can obtain information about observable exoplanets' size, mass, interior structure, bulk composition based on stellar metallicity, distance to the star, and in some cases even their atmospheric composition (Deming and Seager, 2017), we do not know their properties in enough detail to establish urability. However, mapping urability factors onto actual or notional exoplanets can guide research approaches beyond current considerations of habitability.
One example of applying urability to exoplanet environmental models would be that in addition to the habitability requirement of liquid water present as oceans, origin-of-life scenarios might also require the presence of subaerial landscapes supporting volcanism and hydrothermal cycling. This requirement could then be used to constrain the set of worlds that might be plausibly urable.
7. Conclusions
Although there is a certain degree of overlap, it is obvious that urable environments must support different contributing factors than are required for habitability. It is also important to note that if these factors are not present, the only way for a habitable planet to harbor life is for microorganisms to be delivered from an originally urable planet where life has already begun. Finally, there is a paradox to consider: Once life has arisen on a urable world, that planet may then become inurable and prevent a second origin of life.
An example of this curious situation possibly occurs on our own planet, which Darwin (1871) understood intuitively. In his well-known letter about life beginning in a “warm little pond” he considered the possibility that prebiotic protein compounds might form there and predicted that “at the present day such matter would be instantly devoured, or absorbed, which would not have been the case before living creatures were formed.”
The concept of urability can be applied to the search for life elsewhere in our own solar system (Longo and Damer, 2020) and guide future investigations related to the urability of a growing number of observed and modeled exoplanets. In its early history the Earth was clearly urable, and perhaps to a lesser degree so was Mars. However, with the passage of time both planets became inurable for different reasons. The decreasing urability and habitability of Mars was primarily related to loss of liquid water on its surface.
In contrast, the Earth has abundant liquid water, but ∼2.5 Gya cyanobacteria began to produce an oxidizing atmosphere by oxygenic photosynthesis. Molecular oxygen is required by many forms of life today, but it is highly reactive and the reduced organic compounds required for life's origin would be degraded by oxidation. This is why an anoxic atmosphere is an important positive urability factor. For instance, Miller-Urey reactions that produce amino acids in reducing atmospheres have negligible yields in an atmosphere of oxidized gases such as carbon dioxide and nitrogen (Stribling and Miller, 1987).
In regard to the icy moons of the outer solar system, Barge and Rodriguez (2021) recently investigated the possible sources of methane that have been detected in the polar plumes of Enceladus and suggested that the methane is plausibly biological in origin, alluding to a possibly inhabited global ocean beneath the ice. Given its lack of other urability factors, we agree with these authors who state that “…the odds of life emerging on Enceladus are somewhat unfavourable.”
Although interplanetary spacecraft and simulation studies have determined that several bodies in our solar system might be habitable and able to sustain life as we know it, the more rarified conditions required for life to begin have only recently started to emerge. There is a growing consensus that the origin of life is an event that could plausibly take place at distinct locales that were prevalent during the dynamic early window of a planetary body's history. We hope that for future investigators, the working terminology and attendant set of factors proposed here may serve to expand and refine our understanding of how life can begin on worlds such as the Earth and Mars, but also elsewhere in the universe.
Footnotes
Authors' Contributions
D.D. conceived the term “urability” and wrote the first draft manuscript. B.D. expanded the scope of the draft, suggested the design of the urability graph and additional text throughout. F.C. wrote portions of Section 6 and performed significant editing of the entire article.
Acknowledgments
The authors thank several of their knowledgeable colleagues who read the first drafts of this article and provided their valuable comments.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Abbreviations Used
Associate Editor: Norman Sleep