
Abstract
Evidence of stable liquid water oceans beneath the ice crust of moons within the Solar System is of great interest for astrobiology. In particular, subglacial oceans may present hydrothermal processes in their abysses, similarly to terrestrial hydrothermal vents. Therefore, terrestrial extremophilic deep life can be considered a model for putative icy moon extraterrestrial life. However, the comparison between putative extraterrestrial abysses and their terrestrial counterparts suffers from a potentially determinant difference. Indeed, some icy moons oceans may be so deep that the hydrostatic pressure would exceed the maximal pressure at which hydrothermal vent organisms have been isolated. While terrestrial microorganisms that are able to survive in such conditions are known, the effect of high pressure on fundamental biochemical processes is still unclear. In this study, the effects of high hydrostatic pressure on DNA synthesis catalyzed by DNA polymerases are investigated for the first time. The effect on both strand displacement and primer extension activities is measured, and pressure tolerance is compared between enzymes of various thermophilic organisms isolated at different depths.
Introduction
Terrestrial life is possible under a large range of chemical and physical conditions. Extremophilic microorganisms push the boundaries of these ranges further than that expected from an anthropocentric point of view, inhabiting extreme environments previously thought to be too hostile for life. Yet, even extremophilic life is limited to environments above a certain level of water availability (Belilla et al., 2019; Hallsworth et al., 2007). Hence, looking for liquid water bodies remains essential for the search for extraterrestrial life and habitable environments. In this context, the subglacial oceans of icy moons of the Solar System are of particular interest (Jebbar et al., 2020; Taubner et al., 2020).
Among them, Europa and Enceladus retain most of the attention. Several lines of evidence show that the water would be in interaction with rock, which would enable enrichment of a subglacial ocean with various ions, metals, and organic molecules. Characterization of these interactions is indeed a major goal of future space missions (Cable et al., 2016; Europa Study Team, 2012). Moreover, these subglacial oceans may possess hydrothermal processes at their bottom. While stronger evidence supports this hypothesis for Enceladus (Hsu et al., 2015; Matson et al., 2007; Sekine et al., 2015; Waite et al., 2017), geochemical models also argue in favor of abyssal hydrothermal processes for Europa (Lowell and DuBose, 2005; Zolotov, 2007).
Such extraterrestrial abyssal hydrothermal environments would be expected to show similarities to their terrestrial counterparts: hydrothermal vents and black smokers. On Earth, these oceanic environments are observed at various depths, ranging from shallow waters to 5 km deep. While displaying extreme conditions such as temperature, pressure, and sometimes alkaline pH or heavy metal abundancy, these environments are actually associated with complex and diverse ecosystems, offering energy sources and chemical and physical gradients for life. In the absence of light, the primary producers of hydrothermal vent environments are extremophilic chemolithotrophs. These microorganisms which belong either to the Bacteria or Archaea domain, therefore make good models for testing habitability and have already been used as such (Taubner et al., 2018).
However, the physical and chemical conditions may differ in extraterrestrial abysses. On Earth, the deepest hydrothermal vent lies at ∼5 km below the sea level with in situ pressure of 50 MPa (Connelly et al., 2012). The deepest point in the ocean occurs in the Mariana Trench at 11 km below the sea level and at a pressure of ∼108 MPa. In shear contrast, subglacial oceans of several moons could be hundreds of kilometers deep (Schmidt and Manning, 2017). On Europa, for example, the ocean is expected to be, depending on the model, up to 100–200 km deep (Anderson et al., 1998, 1997; Marion et al., 2003; Pappalardo et al., 1999; Spohn and Schubert, 2003; Thomas et al., 2016) and beneath a 10 km thick ice shell (Park et al., 2015), with hydrostatic pressures as high as 130–260 MPa (Naganuma and Uematsu, 1998).
Although most of the terrestrial biosphere is permanently under high hydrostatic pressure (HHP), as 88% of the ocean volume is above 10 MPa (Jebbar et al, 2015), its effects on complex biological processes remain to be explored. Deep sea organisms have to cope with the effects of HHP at the molecular level. Indeed, following Le Chatelier's principle, increases of HHP promote a reduction of volume through production of charges and condensation of biomolecules (Abe et al., 1999; Gross and Jaenicke, 1994). In particular, dissociation of large molecular assemblies such as ribosomes (Gross et al., 1993) and DNA–protein complexes (Kawano et al., 2005) is favored under HHP. Upon a higher pressure increase, protein unfolding is facilitated (Jebbar et al., 2020); most proteins losing their tertiary structure at 400 MPa (Aertsen et al., 2009).
Before unfolding, HHP may have either an inhibitory or a stimulating effect on enzymatic activity, depending on the enzyme (Eisenmenger and Reyes-De-Corcuera, 2009). Notably, a peptidase complex from an abyssal hyperthermophilic archaea has been shown to be activated by HHP (Rosenbaum et al., 2012). DNA is also reversibly compacted by HHP but remains exceptionally stable as it resists denaturation at even 1 GPa (Girard et al., 2007; Hedén et al., 1964). Experimental data show that nucleic acids are stabilized by HHP (Dubins et al., 2001; Macgregor, 1996). As a consequence, all biological processes that require an opening of the DNA double-helix, such as replication, transcription, and translation, can be impaired by HHP (Macgregor, 2002).
Yet, DNA synthesis has never been studied under HHP. DNA replication is one of the most conserved processes, which must occur in every cell before cell division and, thus, to ensure successful inheritance of genetic traits. In cellulo, this process is catalyzed by a class of enzymes called DNA polymerases (DNAPs), which take part of a larger complex, the replisome. DNAPs require a DNA strand primed by a DNA or RNA strand with 3′-hydroxyl extremity, deoxyribonucleotide triphosphates (dNTPs), and divalent cations, Mg2+ or Ca2+ (Ralec et al., 2017). The simplicity of this process has made possible all existing PCR technologies, allowing fast and unexpensive amplification of DNA.
In this context, thermostable DNAPs from organisms isolated in hot environments are routinely used. In particular, many B-family DNAPs from hyperthermophilic archaea, showing high thermostability, extension rate, and fidelity, have been characterized (Zhang et al., 2015). Some were isolated from deep sea organisms that are permanently exposed to HHP. While their activity is high at ambient pressure, there has been no characterization at physiological or even higher pressure conditions. To the best of our knowledge, it has only been showed that a pressure of 89 MPa strongly enhances thermostability of DNAPs from both deep sea and shallow waters (Summit et al., 1998).
In this study, the effects of HHP as high as 100 MPa on activity of five thermostable DNAPs are investigated for the first time.
Materials and Methods
DNAP production and purification
To facilitate the interpretation of the effects of HHP on polymerase activity, we only used enzymes without 3′ → 5′ exonuclease activity, which allows proofreading, either naturally deprived of this activity or by using exonuclease-deficient mutants (exo−).
Exonuclease-deficient Pyrococcus abyssi B-family DNAP (PabPolB exo−) (D215A) with an Nter his-tag sequence MGSSHHHHHHSSGLVPRGSH was produced and purified as previously described (Gueguen et al., 2001; Gouge et al., 2012). Briefly, enzyme was recombinantly expressed in Escherichia coli Rosetta 2 (DE3) cells transformed with a pET28 vector (Novagen) in which gene of PabPolB exo− was inserted between NdeI and BamHI restriction sites. Cells were grown to an optical density of 0.7 in Luria Broth medium at 37°C under shaking and in the presence of kanamycin (50 mg/mL). Expression was then induced by the addition of 1 mM IPTG, and cells were incubated overnight at 30°C.
After centrifugation at 3000 g for 20 min at 4°C, cell pellets were suspended in 50 mM Tris-HCl, pH 7, 200 mM NaCl, 9 mM imidazole, and 0.1% Triton X-100 buffer to which were added protease inhibitors (Roche). Cells were lysed by sonication, and lysate was incubated 1 h at ambient temperature after the addition of 50 μg/mL of DNase I. A heat treatment of 75°C during 30 min was applied to the supernatant before purifying the protein by column chromatography. HisTrap HP™ with nickel-affinity resin, HiTrap™ Heparin HP with affinity for DNA-binding proteins, and size exclusion Superdex™ 200 10/300 GL columns (GE Healthcare) were successively used.
After dialysis, protein was stored at −20°C in a final buffer containing 50 mM Tris-HCl, pH 7.5, and 2 mM MgCl2. Quality and purity of the sample were checked by electrospray ionization at the mass spectrometry platform at Institut de Biologie Structurale. Observed mass of dominant species was in agreement with a loss of the MGSSHHH fragment at the N-terminal part of the protein sequence. A final concentration of 1.16 μM (0.106 mg/mL) of protein was measured with a NanoDrop™ device from Thermo Fisher. Molar extinction coefficient was determined by an online calculator (
Other proofreading-lacking DNAPs were all purchased. Taq from Thermus aquaticus, Vent® (Tli) exo− from Thermococcus litoralis, and Deep Vent® (Psb) exo− from Pyrococcus species GB-D were purchased from New England Biolabs. Pfu exo− from Pyrococcus furiosus was obtained from Agilent. Concentrations of purchased polymerases were given by the manufacturers and were all equal to 2000 U/mL.
Oligonucleotides and template DNA
For extension assays, template DNA was the M13mp18 singled-stranded circular DNA, 7249 nucleotides (nt) in length, purchased from New England Biolabs. For experiments at 55.5–65.0°C, the template was primed with a 30 nt primer whose sequence was TGCCAAGCTTGCATGCCTGCAGGTCGACTC. For gel analysis experiments, this primer was labeled at the 5′ end with a Cy5 fluorophore. Since primer–template interaction was no longer stable at higher temperatures, we used an 87 nt primer instead whose sequence was TGCCAAGCTTGCATGCCTGCAGGTCGACTCTAGAGGATCCCCGGGTACCGAGCTCGAATTCGTAATC ATGGTCATAGCTGTTTCCTG for experiments at 70.0°C.
For strand displacement assays, template DNA was an oligonucleotide 87 nt in length whose sequence was CAGGAAACAGCTATGACCATGATTACGAATTCGAGCTCGGTACCCGGGGATCCTCTAGAGTCGACCT GCAGGCATGCAAGCTTGGCA and which was labeled with a 6-FAM fluorophore at the 5′ end and primed with a 30 t quencher whose sequence was TGCCAAGCTTGCATGCCTGCAGGTCGACTC. Fluorescence was initially quenched by a Deep Dark Quencher I 3′-modification harbored by a 30 nt oligonucleotide, whose sequence was ATTCGTAATCATGGTCATAGCTGTTTCCTG, complementary to the sequence of the template from the 5′ end.
All oligonucleotides were purchased from Eurogentec (Seraing, Belgium).
High-pressure real-time fluorimetry
All fluorescence measurements were performed at various temperatures using a Jasco (Tokyo, Japan) FP-750 spectrofluorometer modified to accommodate a high-pressure cell manufactured by Top Industrie (Vaux-le-Pénil, France). A total of 200 μL of reaction mixtures were prepared to be put in custom rectangular quartz microcuvettes manufactured by Jasco, which were sealed with plastic film and joint as follows: ∼1 cm2 of DuraSeal™ laboratory stretch film from Diversified Biotech (Dedham) was placed on top of the microcuvette, and an O-ring joint from Sephat (Palaiseau, France) was placed over it to maintain the seal. Mixture volume slightly exceeded microcuvette volume to prevent the entrapment of an air bubble. No evidence of sample leak could be observed with experiments and controls. In particular, no sign of mechanical constraint on plastic film or O-ring joint was seen. Excessive reaction mixture was removed before the experiment.
The measurement chamber was filled with distilled water at the desired temperature. The microcuvette was inserted in a custom metal adapter, which allows fluorescence measurement, then put inside the high-pressure cell while immediately starting signal acquisition. To prevent entrapment of an air bubble inside, excessive water was present in the chamber and removed during closure of the chamber. This allowed a better control and monitoring of temperature equilibrium of the sample.
After sealing, pressure was increased with a manual pump linked to the high-pressure cell through a water-filled metal capillary tube connected to the chamber. Pressure was monitored with a probe manufactured by Top Industrie, located between pump and high-pressure cell. HHP were typically reached in 1–2 min.
Temperature inside the chamber was controlled through the circulation of a heat transfer fluid made with tap water supplemented with 30% ethylene glycol and anti-algae Aqua Stabil from Julabo (Seelbach, Germany). This fluid circulated within the high-pressure cell separately from the chamber and the capillary and was heated with a CORIO CD-200F refrigerated/heating circulator purchased from Julabo. The temperature to which the circulator had to be set to reach the desired temperature for the reaction was empirically determined by measuring it in the chamber under ambient pressure with a calibrated thermocouple.
Real-time strand displacement assays
For real-time strand displacement assays, we used a modified tripartite DNA hybrid (Dorjsuren et al, 2009). Reaction mixtures consisted of 50 mM Tris-HCl, pH 8.8, 50 mM KCl, 2 mM MgCl2, 2 mM DTT, 200 μM dNTPs, and 200 nM tripartite DNA hybrid. PabPolB exo− enzyme concentration was 46.4 nM.
Excitation was made continuously at 480 nm, and emission at 525 nm was measured every second. Total run length was 15 min.
Real-time extension assays
For real-time extension assays, reaction mixtures consisted of 50 mM Tris-HCl, pH 8.8, 50 mM KCl, 2 mM MgCl2, 2 mM DTT, 200 μM dNTPs, and 418 pM primed M13mp18. PicoGreen™ (Thermo Fisher) was added at final 3200-fold dilution from the commercial solution. All commercial enzymes were incubated in the same buffer, except Pfu exo−, which was incubated in the buffer provided by the manufacturer (Agilent). PabPolB exo− enzyme concentration was 46.4 nM. Commercial enzymes were used at a concentration of 0.01 U/mL.
Excitation was made at 485 nm every 20 s, and emission at 520 nm was measured for 1 s. Total run length was either 15 or 30 min.
Gel analysis of products
Reactions were conducted similarly to the real-time extension assays in the same thermostated fluorescence cell with the fluorimeter turned off and without PicoGreen. The primer was labeled with a Cy5 fluorophore at the 5′ end. After a 30 min extension reaction under various pressures, decompression, chamber opening, and sample retrieval were performed as fast as possible, and reactions were stopped by adding an equal volume of 95% formamide and 10 mM EDTA on ice. This procedure took ∼60 s, from the start of decompression to the addition of the formamide. Primer extension products (20 μL) were loaded onto TAE-agarose 1% gels and separated by electrophoresis at 100 V and 100 mA for 5 h. Bands containing hybridized extended Cy5-labeled primer were revealed with a Gel Doc XR+ Gel Documentation System (Bio-Rad) used with the fluorescence mode.
Results
Gel analysis of products synthesized under HHP
We first determined the effects of HHP on global DNA synthesis by PabPolB exo− by 224 analyzing, after decompression of the sample, the length of primer extension products synthesized under various pressure values and at 55.5°C or 65.0°C. Fluorescent products labeled with Cy5 at the 5′ end of the elongated 30 nt primer were separated on agarose gel electrophoresis (Fig. 1).
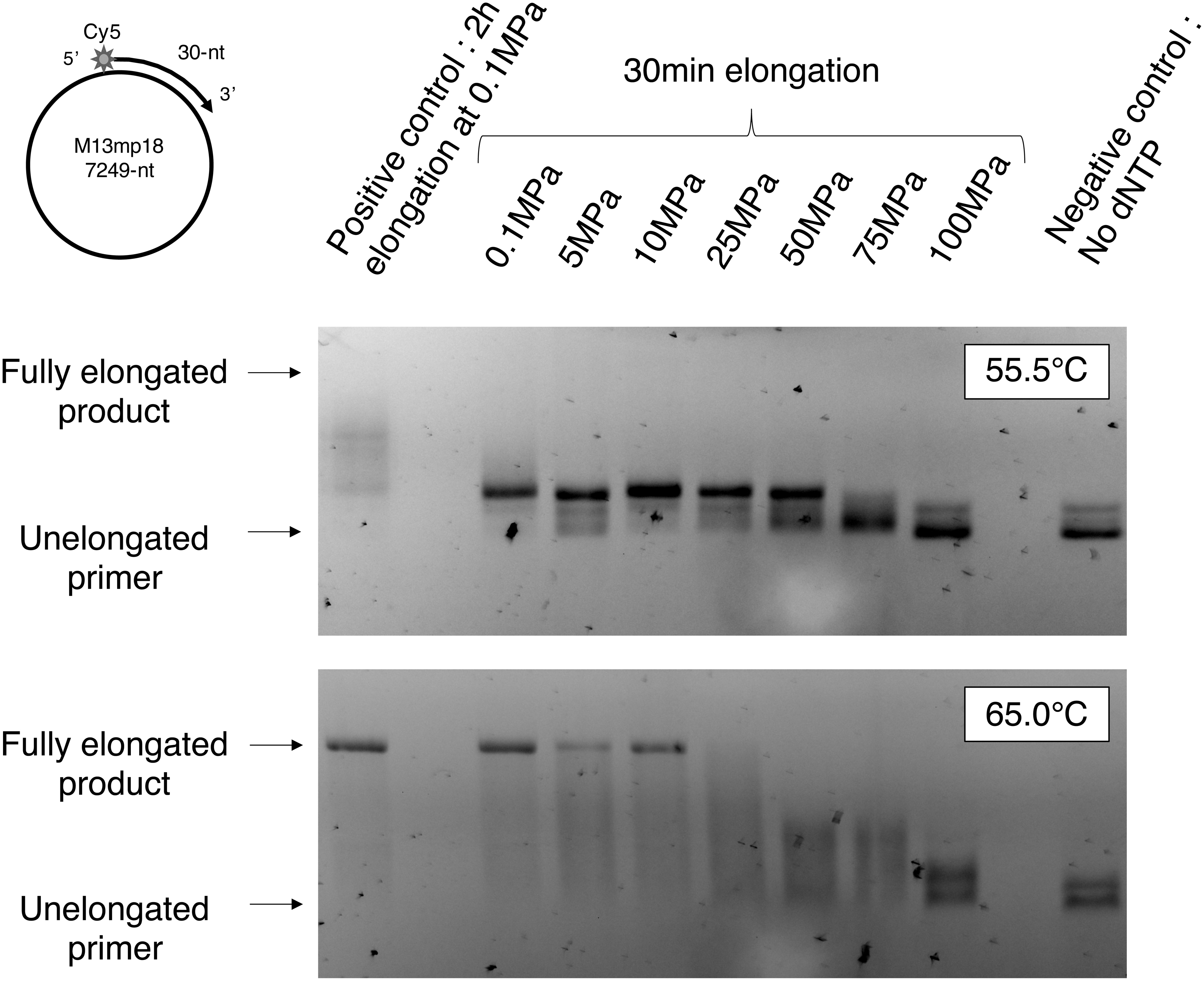
Gel analysis of products of PabPolB exo− under HHP. Extension of the Cy5-labeled 30 nt long primer hybridized on an m13MP18 DNA template of 7249 nt in length under pressure values is indicated above and temperatures are indicated on the left. HHP, high hydrostatic pressure.
Products were longer at 65.0°C than at 55.5°C. In contrast, a gradual decrease of product length as pressure increased was observed, from 50 MPa at 55.5°C and from 25 MPa at 65.0°C. Little or no increase in product length in comparison with the negative control was observed at 100 MPa in both conditions.
To analyze the effects of HHP in real time, we used a fluorescence-based tripartite oligonucleotide template system that relies on strand displacement activity by B-family DNAPs (Canceill et al., 1999; Henneke, 2012). The increase of fluorescence caused by strand displacement of the oligonucleotide quenching the fluorescent signal of 87 nt template was used as proxy for PabPolB activity. Strand displacement assays were performed for 30 min at 55.5°C under 0.1, 10, 50, and 100 MPa three times independently (Fig. 2).

Real-time monitoring of strand displacement activity of PabPolB exo− under HHP.
Fluorescence signal of positive control (6-FAM-labeled template annealed to extension primer without the quenching oligonucleotide) slightly decreased over time, presumably caused by photobleaching of the 6-FAM fluorophore. Pressure gradually delayed and slowed down the increase of fluorescence in the enzymatic assays, which showed an inhibiting effect on strand displacement activity. The inhibition could be observed from 10 MPa, and little or no activity could be observed at 100 MPa.
These experiments were also conducted at additional temperatures (55.5°C, 60°C, 65°C, and 70.0°C) and pressure values (0.1, 35, 50, and 100 MPa) (Supplementary Fig. S1). Fluorescence increase was faster at 60°C and 65.0°C than at 55.5°C. In contrast, at every temperature, HHP gradually delayed fluorescence increase. Fluorescence curve of the unquenched template was slightly higher at 65.0°C than at 55.5°C and 60.0°C, which may indicate that basal signal of the fluorophores is dependent on the temperature. At temperatures above theoretical melting temperatures (T m) of primer in the presence of the quenching oligonucleotide, respectively, 58.9°C and 67.1°C, spontaneous denaturation of the tripartite DNA may, however, have happened, as shown by the increase of fluorescence of the negative control at 70.0°C.
Primer extension activity was also directly monitored by measuring the fluorescence of PicoGreen, which increases as it intercalates in increasing amounts of double-stranded DNA during primer extension. The DNA template was single-stranded M13mp18, allowing extension up to the order of the kbp. We monitored primer extension activity for 30 min at 55.5°C under 0.1, 10, 50, and 100 MPa three times independently (Fig. 3).

Real-time monitoring of primer extension activity of PabPolB exo− under HHP. Fluorescence signal of PicoGreen intercalant during the extension of the unlabeled 30 nt long primer hybridized on an m13MP18 DNA template. The curves represent the average fluorescence over three independent experiments, and the colored area corresponds to the standard deviation. Controls are indicated.
Curves showed a clear biphasic behavior at 0.1, 10, and 50 MPa. Duration of the second phase was reproducibly increased by HHP. In contrast, no clear biphasic behavior could be observed at 100 MPa.
Variability was higher in this experiment compared with strand displacement assays, as indicated by larger standard deviation values, especially for the positive control (incubation at Pamb for 2 h before the fluorescence measurement to achieve maximal primer extension). As with strand displacement experiments, fluorescence decreased over time. All other things being equal, pressure tended to increase all curves, showing a putative effect of HHP on intrinsic PicoGreen fluorescence or insertion in double-stranded DNA.
Nonetheless, pressure inhibition of extension activity was still observable as the plateau was reached later, or not reached at all, at 50 and 100 MPa. The observed plateau unlikely resulted from dNTPs depletion as they were provided in large excess and could have resulted instead from unbounded PicoGreen shortage. In addition, maximum value of first derivative of the curves, corresponding to the maximum increase of fluorescence, decreased with pressure and the corresponding inflexion points, when applicable (0.1, 10, and 50 MPa) were delayed (Supplementary Fig. S2).
To obtain more precise and quantitative information about the fluorescence variation over time as a function of pressure, primer extension assays were replicated three times with additional pressure values and with shorter acquisitions (15 min). The curves were transformed so that the value of fluorescence for positive control was equal to 100% and the value of fluorescence for the negative control was equal to 0%, both at every instant. This way, fluorescence was analyzed relative to controls, and comparison between conditions was easier (Supplementary Fig. S3).
By calculating first derivative of these curves, we then determined relative increases of fluorescence as a function of time, roughly corresponding to elongation speeds. Fastest increase in fluorescence over the whole experiment and punctual fluorescence increase were calculated (Fig. 4). Maximum elongation efficiency was achieved during the whole 15 min run, and fluorescence increase at various moments of the run was negatively affected by HHP above 10 MPa. Milder pressure values (below 10 MPa) had little effect on fluorescence increase.
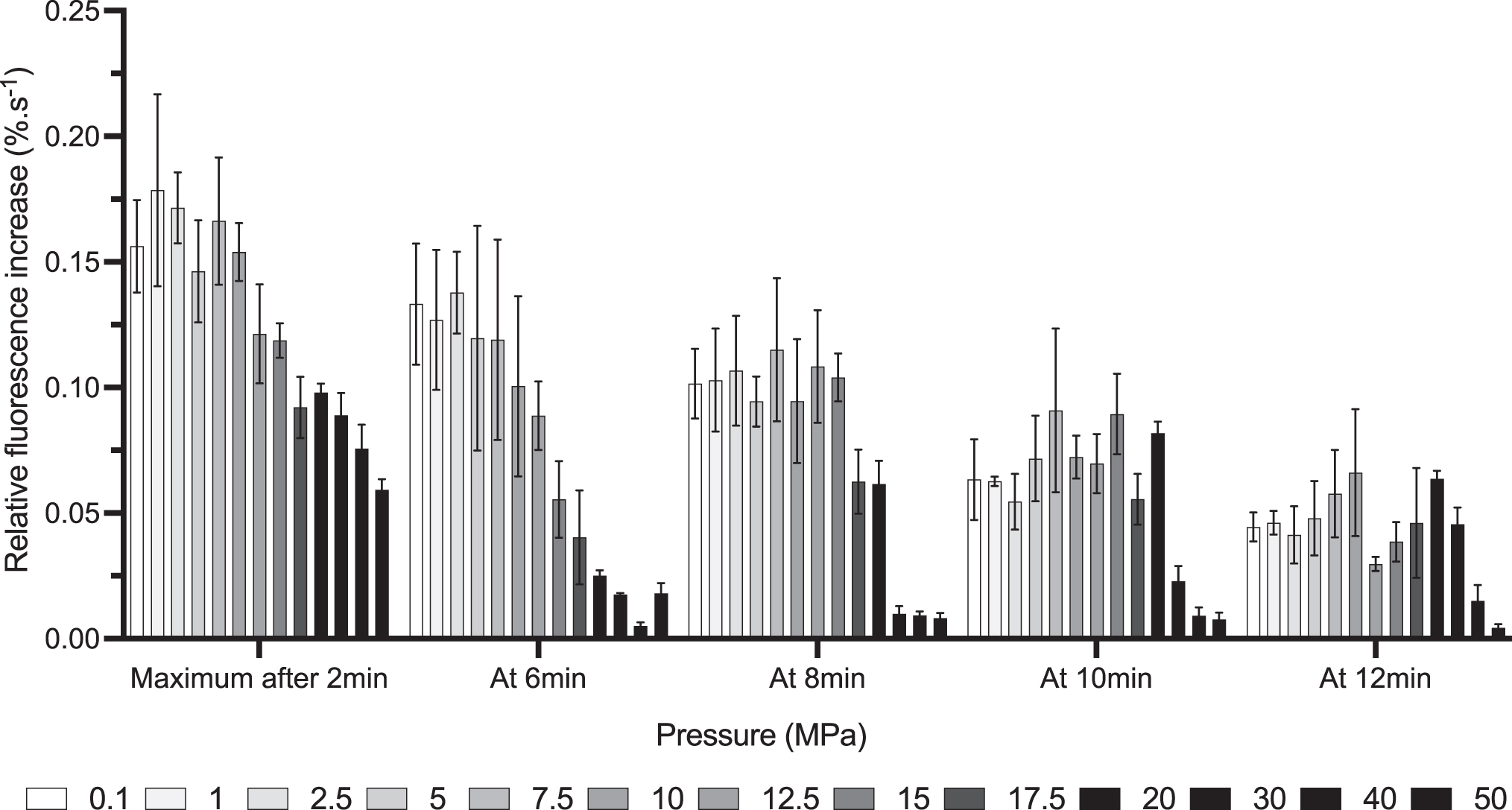
Effect of HHP on elongation speed of PabPolB exo−. Primer extension curves were normalized using positive and negative controls as, respectively, superior and inferior limits. First derivatives were calculated for each of the three independently obtained curves. Error bars correspond to standard deviation. Colors represent pressure value (see legend).
To determine whether the previously observed inhibiting effect of HHP on PabPolB exo− was irreversible, samples were incubated at 55.5°C and 100 MPa during 15, 30, or 60 min. Upon decompression, measurement was immediately started, and fluorescence was measured for 15 min. The experiment was performed three times independently, and average relative fluorescence curves were determined (Fig. 5).
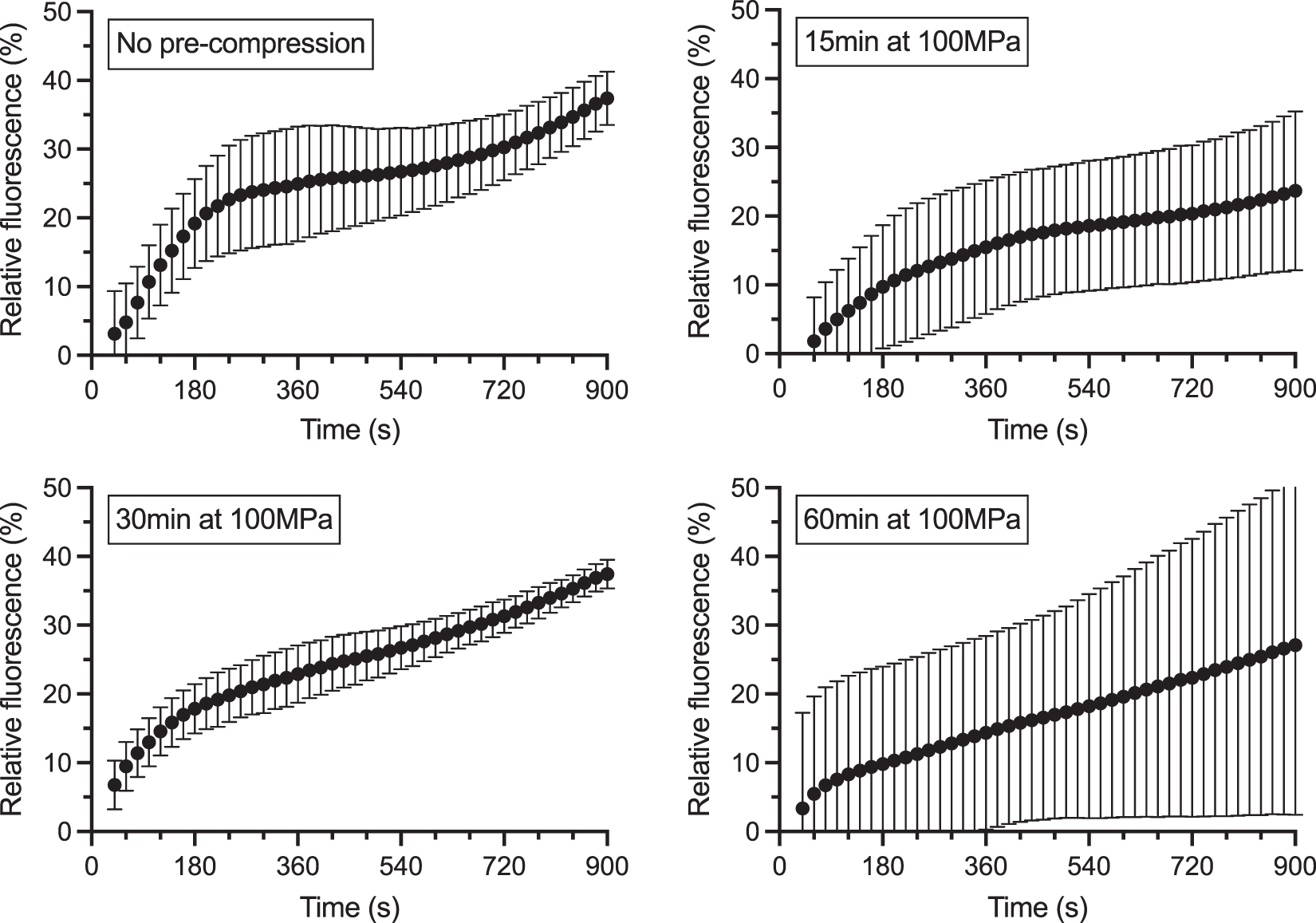
Effect of pre-compression on primer extension activity of PabPolB exo− at Pamb. Primer extension curves were normalized using positive and negative controls as, respectively, superior (100%) and inferior (0%) limits. The curves represent the average fluorescence over three independent experiments. Incubation under HHP before fluorescence measurement is indicated. Error bars represent standard deviation.
Preincubation of the sample under HHP, even for longest durations, did not impair significantly the primer extension activity of the enzymes when pressure was decreased to ambient pressure. This shows that the effect of preincubation on PabPolB exo− activity was largely reversible.
We then compared sensitivity with HHP of DNAPs from various organisms: TaqPol from the thermophilic bacteria T. aquaticus and B-family DNAPs from hyperthermophilic archaea. Only exo− mutants of archaeal DNAPs were used: exo− Pfu Pol (Agilent) and Vent exo− (NEB), respectively, from P. furiosus and T. litoralis, which have been isolated in a shallow water hydrothermal vents (Kong et al, 1993; Lundberg et al., 1991) and Deep Vent exo− (NEB) from Pyrococcus species Strain GB-D, which has been isolated in a 2 km deep hydrothermal vent (Jannasch et al., 1992), such as P. abyssi. All archaeal enzymes had high sequence identity (Supplementary Fig. S4).
Primer extension activity was measured for 30 min three times independently for each enzyme at 0.1, 10, 50 and 100 MPa, at 55.5°C or 70.0°C. For reactions at 70.0°C, single-stranded m13mp18 was annealed to a longer primer (87 nt rather than 30 nt) to prevent melting of the hybrid. Curves were transformed as described previously, and the average maximum fluorescence increase observed during the whole run was determined. For each enzyme, the value at 0.1 MPa was set to 100% to compare their elongation efficiency independently (Fig. 6).
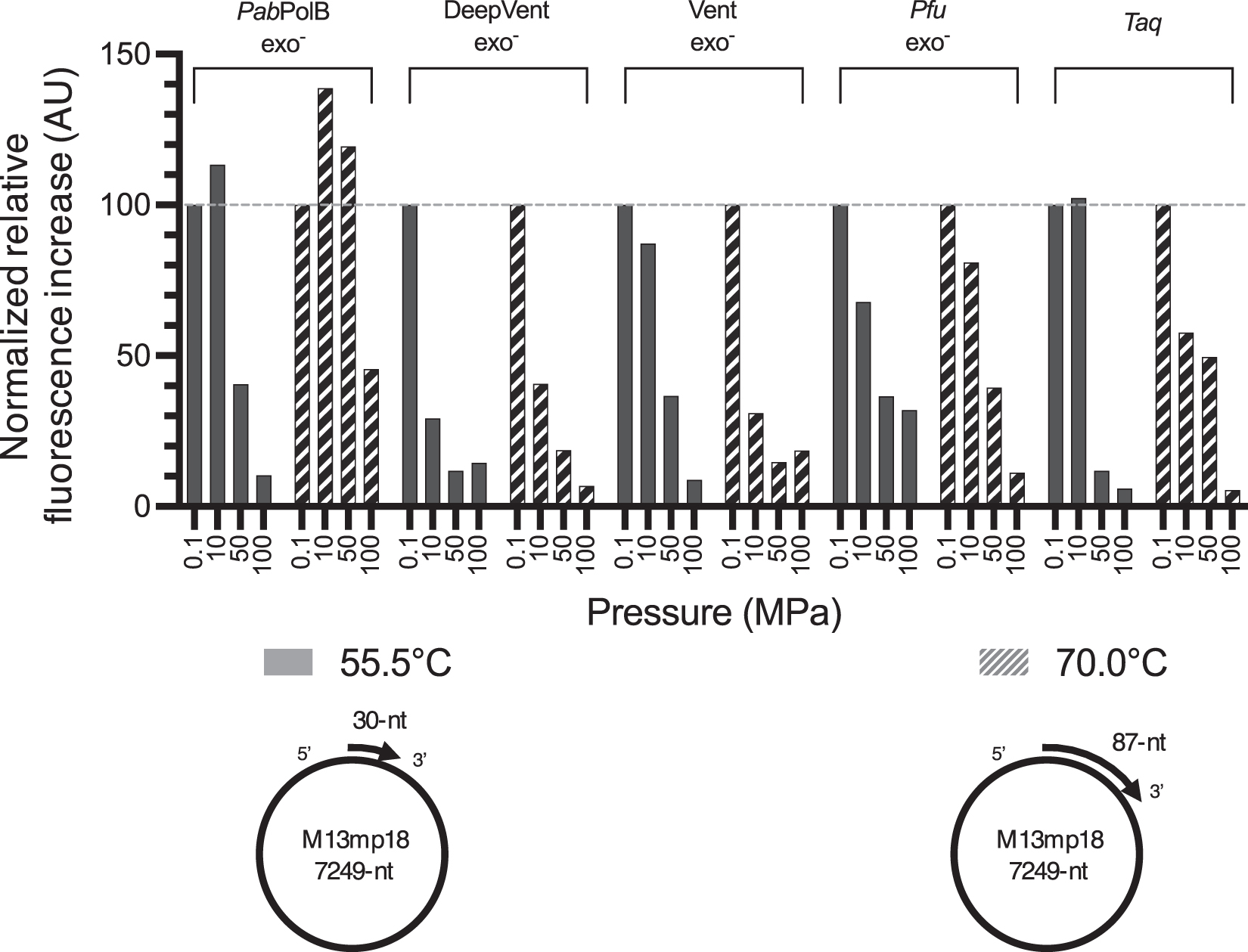
Comparison of pressure sensitivity of various DNAPs. Primer extension curves were normalized using positive and negative controls as, respectively, superior and inferior limits. First derivatives were calculated for each of the three independently obtained curves. Average maximum observed speed is plotted as a function on the enzyme and pressure/temperature combination. To facilitate observation of the effect of HHP on elongation speed, for each temperature and enzyme, maximum speed at 0.1 MPa was set to 100% and speeds at the other pressure values were compared to them. DNA template–primer hybrids used in each temperature condition are schematically represented below legend. DNAP, DNA polymerase.
All commercial archaeal enzymes showed gradual inhibition by HHP at both 55.5°C and 70.0°C. However, increasing the temperature compensated the decrease of activity induced by HHP. Taq enzyme turned out to be more pressure-tolerant at 70.0°C than Vent exo− and Deep Vent exo−. At 55.5°C, Deep Vent exo− was the most pressure-sensitive DNAP as a dramatical decrease of maximum achieved elongation speed already started at 10 MPa. In contrast, Pfu exo− DNAP showed reduced inhibition by HHP in most conditions compared with the other commercial archaeal enzymes. However, Pfu exo− was the only enzyme used in the commercial buffer that contained Triton X-100.
Unlike what was observed at 55.5°C, maximum elongation speed of PabPolB exo− was significantly increased at both 10 and 50 MPa when reaction occurred at 70.0°C. None of the other tested enzymes showed such behavior. Moreover, at such temperature, the maximum observed speed was only halved by 100 MPa exposure.
We have shown, for the first time that, unlike many enzymes from both extremophiles and non-extremophiles whose activities can be enhanced by HHP (Eisenmenger and Reyes-De-Corcuera, 2009), DNAPs are inhibited by HHP above 10 MPa. Effects of HHP on proteins are rather complex and depend on the protein. As a general rule, response to pressure is governed by Le Chatelier's principle, which predicts that a system at the equilibrium will respond to an increase of pressure through a decrease of volume (Heremans, 2009; Jebbar et al., 2015). For proteins, this decrease may result from two distinct effects: reversible compression, especially for proteins with large internal cavities, and unfolding, which allows additional condensation of water molecules (Doster et al., 2003; Eisenmenger and Reyes-De-Corcuera, 2009).
At 400 MPa, most proteins are, hence, fully unfolded (Aertsen et al., 2009). Dissociations of molecular assemblies, such as the ribosome (Gross et al., 1993; Pande and Wishnia, 1986), are also promoted under HHP as it releases sites for water condensation, most assemblies being fully dissociated at 200 MPa (Aertsen et al., 2009). Yet, it is not possible to impute the observed HHP inhibition of DNAPs on unfolding. In particular, the post-pressurization experiments showed that, whatever its cause, the nearly full inhibition at 100 MPa was reversible, just like what was observed with an RNA polymerase from non-thermophilic abyssal bacteria (Kawano et al., 2004).
While gel analysis of primer extension products provides a simple way to assess the effects of HHP on DNA polymerization activity, the fact that this analysis can only be made after decompression may bias the results as fast pressure change could have specific effects such as sudden and quick stimulation of primer extension. Nonetheless, these experiments showed that the decrease of total elongated DNA synthesized by PabPolB exo− was observable from 50 MPa at 55.5°C and from 25 MPa at 65.0°C, indicating that pressure inhibition of DNAP activity depends on temperature, as confirmed by the primer extension assays. It should also be pointed out that PabPolB could realize rolling circle replication of which requires displacement of elongated primer and, hence, relies on strand displacement activity, which we showed was inhibited by HHP. Such effect could contribute, to some extent, to the observed inhibition.
In contrast, it has been shown that HHP, up to 89 MPa, actually increase thermostability of both DNAPs of Pyrococcus strains and Taq DNAP (Summit et al., 1998). Hence, the HHP inhibition may result instead from subtle changes in protein structures and dynamics that enhance stability at the expense of activity. Psychrophilic enzymes, which present flexibility-inducing traits, may therefore be less sensitive to pressure inhibition. However, no experimental comparison of pressure sensitivity between homologous thermophilic, mesophilic, and psychrophilic deep-sea enzymes has been published, to our knowledge. Nonetheless, synergy between psychrophilic and piezophilic protein traits has been pointed (Capece et al., 2013; Carré et al., 2022).
Strand displacement activity assays were limited by the fact that interaction between the DNA template and quencher-harboring oligonucleotide is sensitive to pressure. While short simple double-stranded DNA fragments can be destabilized by HHP (Gunter and Gunter, 1972; Hughes and Steiner, 1966), HHP tends to increase the stability of DNA hybrids (Macgregor, 1996; Rayan and Macgregor, 2005). Therefore, if no interference caused by the Deep Dark Quencher I 3′-modification and 6-FAM fluorophore is assumed, interaction between the oligonucleotides was reinforced under HHP.
Hence, in addition to any direct effect on the enzyme, strand displacement activity was inhibited by the strengthened interaction between the DNA template and the strand that the enzyme had to displace. Indeed, total inhibition at 100 MPa was observed during strand displacement assays, whereas some activity could still be observed during primer extension assay. Therefore, assays that directly measure extension activity should be preferred to more indirect methods for HHP studies of DNAPs. One should notice that, at higher temperatures, the decreased stability of double-stranded DNA could compensate for HHP inhibition of strand displacement activity.
The biphasic behavior observed in primer extension activity curves is consistent with the biphasic model of DNAP enzymatic activity. In such a model, the first short phase corresponds to the addition of the first dNTP to the primer by all enzymes, which is faster than the following dNTP polymerizations, which correspond to the second phase (Berdis, 2009). Our data suggest that this second phase is more affected by HHP as the quick initial increase of fluorescence was observed at 10 and 50 MPa, whereas the second more gradual increase appeared to be delayed at these pressure values and was mostly absent at 100 MPa. This stresses the hypothesis that HHP impairs DNAP activity through direct inhibition of primer extension rather than substrate binding.
Surprisingly, high tolerance to HHP was observed with Taq DNAP, which originates from T. aquaticus, a surface thermophilic bacterium. Since T. aquaticus is less thermophilic than the archaea from which the other tested DNAPs were from, its proteins may be less thermostable, thus less resilient and less rigid (Aguilar et al., 1997; Tehei and Zaccai, 2007; Zaccai, 2013), which could explain its low sensibility to HHP. However, Taq is A-family DNAP, whereas all the other tested enzymes are from the B family. Depending on the fold and especially the internal cavities, intrinsic flexibility and, hence, sensitivity to HHP could vary between DNAP families.
When compared with the other exo− B-family archaeal DNAPs, PabPolB showed increased tolerance to HHP. As little sequence variation is observed between these enzymes, subtle structural changes, at the level of internal cavities or interdomain hinges, may explain these differences. PabPolB exo− was the only tested enzyme that was not purchased but produced and purified by us. In general, little information about piezophilic adaptation can be inferred from protein sequences as they are mostly dictated by temperature in abyssal environments (hot hydrothermal vents or cold waters and sediments) (Nath and Subbiah, 2016). Nonetheless, in comparison to their non-piezophilic homologs, enzymes from deep-sea organisms tend to have denser cores with smaller residues, less salt bridges (Carré et al., 2022).
Still, increasing temperature compensated HHP inhibition of all tested enzymes. This could result from two separate causes. First, the evaluated enzymes are hyperthermophilic, their optimal temperature being between 72°C and 80°C for archaeal DNAPs (Zhang et al., 2015); increasing the temperature from 55.5°C to 70.0°C may, hence, be sufficient to account for the increase of the maximum elongation speed. Second, temperature may directly reduce the inhibiting effects of HHP through the increase of molecular motion and flexibility (Daniel and Cowan, 2000). Nonetheless, the present work stresses the importance of heat sources in deep environments where organisms are constantly under HHP. This is especially true for putative extraterrestrial abysses, which could be deeper and exposed to pressure values above 100 MPa (Naganuma and Uematsu, 1998).
Pressure adaptation of abyssal microorganisms still remains unclear. In particular, complex processes such as DNA replication can be affected by HHP in a complex manner as it involves multiple protein, multimeric assemblies, proteins, and DNA. Moreover, care should be taken when extrapolating data obtained in vitro on simple protein systems studied in buffers of given composition to the physiological intracellular context. Cytoplasmic crowding, interactions with piezolytes or with the other replisome proteins, may indeed limit the effects of HHP on DNAPs. While HHP tends to disrupt large molecular assemblies and, hence, could impair replisome formation, depending on the volumetric parameters of each protein–protein and protein–DNA interaction, GINS or PCNA complexes could limit pressure sensitivity of DNAPs. In particular, it has been shown that the addition of PCNA improves strand displacement activity (Henneke et al., 2005) and fidelity (Castillo-Lizardo et al., 2014) of PabPolB in vitro.
Nonetheless, the deepest hydrothermal vents on Earth are found at a depth of 5 km, and most are found above 2 km deep, limiting the pressure conditions to which these organisms are actually exposed. Another possibility could be that high temperatures are sufficient to compensate inhibitory effects of HHP. Indeed, most of our experiments were made at 55.5°C, which is below the temperature range at which Thermococcales species such as P. abyssi grow (Erauso et al., 1993).
This could stress even more the dependency on temperature of abyssal hyperthermophilic microorganisms and the need to search for heat sources in the extraterrestrial deep oceans.
Conclusions
In this work, the effects of HHP on DNAPs of abyssal and surface (hyper)thermophilic microorganisms was explored for the first time. By using various fluorescence-based approaches, including real-time fluorimetric assays, we showed that HHP inhibits total DNA synthesis, strand displacement activity, and extension rate. These effects were shown to be reversible, mitigated by higher temperature, and dependent on the enzymes.
Footnotes
Acknowledgments
IBS acknowledges the integration into the Interdisciplinary Research Institute of Grenoble (IRIG, CEA). High-pressure experiments were conducted with the BioHP platform of the IBS.
Authors' Contributions
All authors proposed and contributed in their respective expertise and participated in writing the article. L.C. designed the research, conducted the experiments, analyzed the data, wrote the article, and made the figures. G.H., E.H., and D.F. designed the research and corrected the article. E.G. and B.F. designed the research, and wrote and corrected the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the French National Research Agency in the framework of the Investissements d'Avenir program (ANR-15-IDEX-02), through the funding of the “Origin of Life” project of the University of Grenoble Alpes.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Abbreviations Used
Associate Editor: Don Cowan
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.