
Abstract
The mineral reaction pathways that yield organic compounds of increasing complexity would have required a means of protective screening against strong ultraviolet radiation for macromolecular assembly on early Earth. In this study, a bacterial chromosomal plasmid DNA was used as a model biomolecule that represents a complex polymeric nucleic acid containing genetic information. The plasmid DNA was exposed to UV radiation through a medium containing air, water, iron (Fe3+), or silica-iron rich aqueous solutions. Our results demonstrate that the plasmid DNA underwent covalent breakage in an aqueous solution when exposed to UV radiation but was shielded against damage due to the presence of iron and silica. It is demonstrated that a suspension of ca. 40 nm colloidal particles of silica gel embedded with Fe3+ ions adsorbed on silanol groups that formed nanoclusters of noncrystalline iron hydroxide is an extremely efficient shelter against intense UV radiation. The implications for our understanding of primitive Earth and Earth-like planets, moons, and asteroids are discussed. The stability of a chromosomal DNA molecule against UV radiation in the presence of iron and silica may provide support on how macromolecules endured early Earth environments and brought forth important implications on early molecular survival against UV radiation.
1. Introduction
All efforts to explain the origin of life or how life was preserved on a severely UV-radiated early Earth must connect prebiotic chemistry with protobiological processes (Krishnamurthy, 2017). This prebiotic-protobiological connection requires the understanding of what type of environment may permit prebiotic chemistry to transition to that of biotic. Our current description is not satisfactory because what seems to be feasible by prebiotic chemistry ideology does not seem to be compatible with those that are biology-based. The opposite also holds true such that notions of biological origins appear to be inconsistent with what prebiotic chemistry can provide (Krishnamurthy, 2018). An example of this conundrum is the RNA world hypothesis (Copley et al., 2007; Neveu et al., 2013) that postulates RNA must have been the primary prebiotic material because it has the properties of being able to fold into a three-dimensional structure, and it is enzymatic and able to hold genetic information. The hypothesis postulates that the functions of prebiotic RNA eventually gave rise to DNA and proteins that have the independent ability to store genetic material and exhibit enzymatic activities, respectively (Gavette et al., 2016). The RNA world idea has been challenged as to whether it is geochemically plausible to have preservation of genetic material or enzymatic activities under the extreme conditions of the early Earth environment.
It is well known that solar UV radiation is not only harmful for living organisms but is harmful as well for the main biopolymers that sustain life. Today, living organisms on Earth are shielded by the ozone layer against our oxidant atmosphere. The composition of the atmosphere has changed significantly during the history of the planet, mainly due to the metabolic output produced during the course of biological evolution. It has also been shown that, during the early lifeless stages of Earth's history, when prebiotic chemistry was routed toward the first complex organic molecules and the earliest microorganisms started to colonize the planet, the atmosphere was devoid of oxygen and therefore lacking an effective ozone layer (Kasting, 1987). Without an ozone layer, the exposed surface of primordial Earth was a hostile place for any successful physicochemical pathway to biochemical polymers and most certainly for the existence of primitive life-forms. Other forms of UV screens included gases in the early Earth atmosphere such as SO2 and H2S from volcanic outgassing and CH4-generated hydrocarbon smog produced by methanogens or nonbiological processes (Sagan and Chyba, 1997; Cockell, 2001). In terms of natural widespread UV filters, the oceans and seas have been considered to be effective in blocking UV at depths greater than 30 m (Cockell, 2000). Finally, the subsurface crust has also been considered to be a protective haven against UV exposure as proposed by the hydrothermal theory for the origin of life (Baross and Hoffman, 1985; Holm, 1992). In considering all the ways UV can be blocked during the early Earth environment, none are satisfactory in explaining how UV is shielded at a geological timescale for prebiotic viability and stability—especially in areas favorable for concentrating organic molecules by evaporation, such as in shallow ponds (Mulkidjanian et al., 2012; Ranjan et al., 2019).
In this study, we model an efficient shielding environment in vitro against UV radiation that is geochemically relevant to a Hadean geochemical scenario with a reduced methane-rich atmosphere and initial silica-rich alkaline oceans (Saladino et al., 2019; García-Ruiz et al., 2020). The proposed idea is based on the ability of silica and iron solutions and their mixture to absorb UV radiation that covers most of the UVC range (190–280 nm) (Mloszewska et al., 2018). Silica and iron (upwelled iron) have been suggested to be early UV attenuators when atmospheric oxygen content of early Earth was inadequate to generate enough ozone to assuage solar UV radiation (Green and Miller, 1975; Olson and Pierson, 1986; Cockell, 1998, 1999). Iron screening effects against solar UV radiation have also been observed in present terrestrial aquatic extreme environments over phototrophic microorganisms (Pierson et al., 1999). In addition, Archean bacteria mineralized in an iron-silica solution have been shown to have marked resistance to UV compared to non-mineralized bacteria (Phoenix et al., 2001; Mloszewska et al., 2018). Early molecular synthesis and stability of the initial biomolecules prior to the formation of cellular life would have required a surrounding that shielded UV against the breakage of long-chain molecules. Complex polymeric nucleic acids (RNA or DNA) have been speculated to be the first biomolecules and subsequently the surviving material for the passage of genetic information.
It is known that UV radiation on nucleic acids can render various types of base modifications to single and double strand breaks caused by the induction and propagation of sugar and base radicals (Rastogi et al., 2010). These events can be manifested by the direct absorption of ionizing radiation of the DNA molecule or indirectly by water radicals generated by solvent ionizing radiation. In both cases, the consequences would be the production of radical cations, which on spontaneous deprotonation gives rise to DNA radicals (Hutchinson, 1985; von Sonntag, 1987).
We have studied the effects of UV radiation on a bacterial chromosomal plasmid DNA, pUC18, contained in quartz capillary tubes surrounded by air, water, silica, iron, and silica-iron solutions. The absorbance wavelengths of silica and iron solution are coincident with those that inflict DNA damage that results in base modifications and strand breakage (Hutchinson, 1985; Olson and Pierson, 1986); therefore, we have measured the effects of bacterial chromosomal DNA when exposed to long irradiation times of UV protected by silica, iron, and silica-iron solutions. We show that ferric iron and silica-iron mixtures may be essential for the protection of genetic material of the earliest microbial ecosystems and for preserving self-organized complex organic molecules that were presumably formed during the earliest Hadean eon.
2. Materials and Methods
2.1. Instrument and chemical reagents
A high-performance UV transilluminator with an internal cooling system (Model TFS-20V) was used for this study. All experiments reported here were carried out by using a UV light lamp with a wavelength of 254 nm and an intensity of 4700 μW/cm2.
Sodium metasilicate pentahydrate (Fisher Chemicals, USA) and iron (III) chloride (Sigma-Aldrich, USA) were the principal chemical components used for this study. Ultrapure agarose was obtained from NZYTech (Portugal). All chemical mixtures were prepared with ultrapure deionized nuclease-free and autoclave-sterilized water. A silica solution was prepared for the purpose of shielding UV. The concentration of the silica used for the experiment was 70 mM (4200 ppm) (Feth et al., 1961; García-Ruiz et al., 2017). This concentration was comparable to that found in the Aqua de Ney, a hyperalkaline (pH ∼12) silica-rich high methane mineral spring in Siskiyou County, California. The working concentration of Fe3+ was prepared at 0.55 mM, which is the iron concentration reported by Mloszewska et al. (2018) for cyanobacteria protection in early marine photic environments. Both silica and ferric chloride in solution had a maximum absorbance at 220 nm at pH 12.86 with a synergistic absorption at 254 nm (Fig. 1). The UV wavelength selected for the experiment was 254 nm (short UV wavelength), which is the absorbance wavelength of the silica and iron solution that coincides with the UV wavelength that inflicts DNA damage resulting in base modifications and strand breakage (Rastogi et al., 2010).
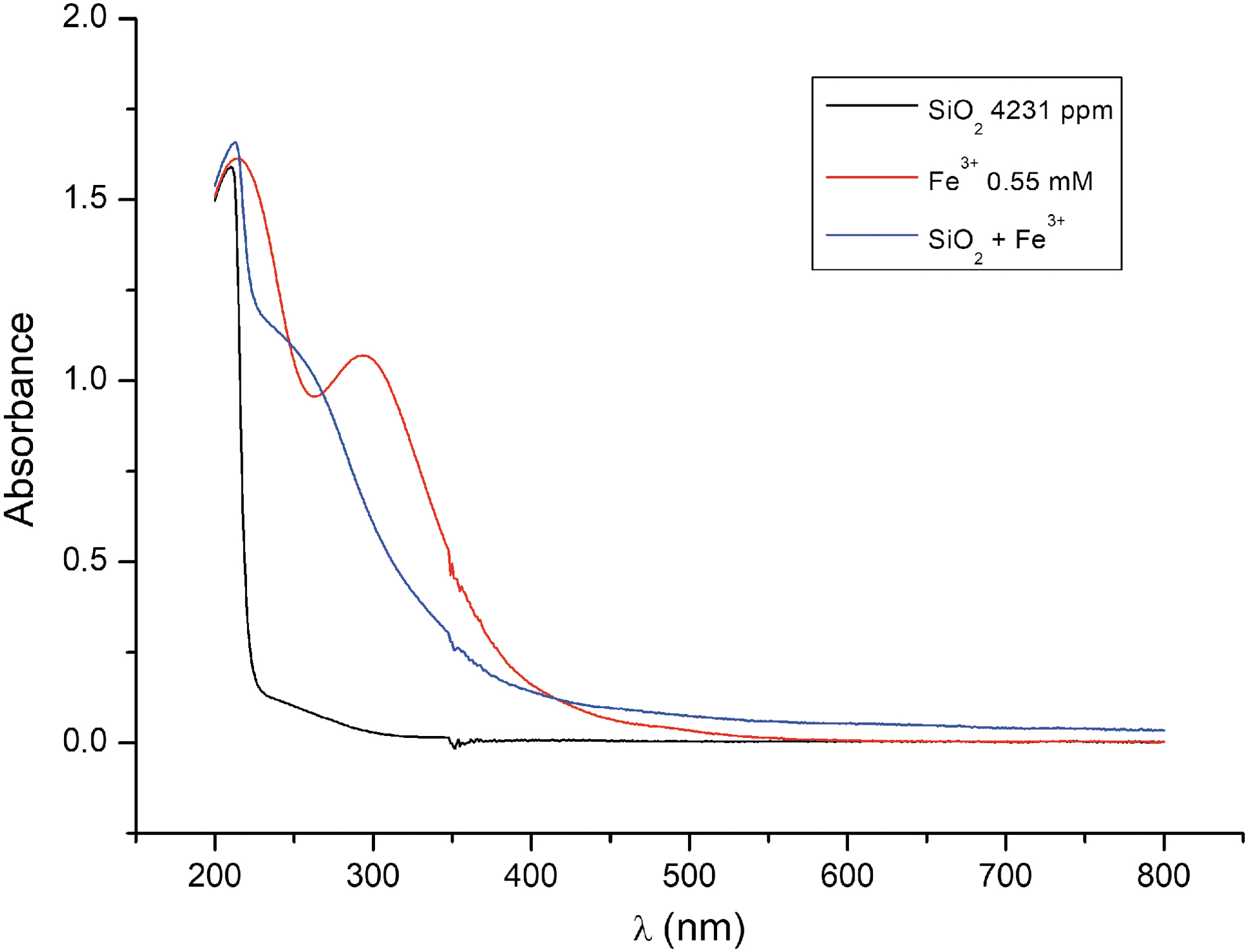
UV-visible absorbance spectra of silicate, Fe3+ chloride, and silica-Fe3+ in water.
The Escherichia coli plasmid cloning vector, pUC18, was used as the DNA substrate to access UV radiation damage. The plasmid was purchased from ThermoFisher Scientific (USA). pUC18 is a double-stranded circular DNA that has 2686 base pairs in length. The stock DNA plasmid concentration was 0.5 μg/μL. Prior to the experiments, the DNA solution was diluted 32-fold in 10 mM Tris-HCl plus 1 mM EDTA, pH 7.5 (TE) in a 1.5 mL sterile Eppendorf tube. The final working concentration of the DNA plasmid was 15.6 ng/μL in TE.
2.2. DNA samples and UV irradiation
Quartz capillaries were purchased from Hilgenberg GmbH (Germany) and used to contain the DNA samples for UV irradiation. Plasmid DNA was prepared at a concentration of 15.6 ng/μL as described above and placed in 0.7 mm inner diameter (with a wall thickness of 0.01 mm) quartz capillaries. A volume of 6 μL of DNA sample was used to fill up a single capillary by capillary action, and both ends were sealed with clay mineral plasticine. During the course of the experiment, the DNA-filled capillaries were secured by metallic clips placed in the bottom of a glass experiment chamber (Fig. 2) with dimensions measuring 15 cm (width) × 20 cm (length) × 20 cm (height) and volume equal to 4.5 L. The UV transilluminator was then placed in an inverted orientation such that the UV illumination can irradiate the entire solution area containing the capillaries. The temperature of the capillaries was measured to be constant at 20°C for all experiments.
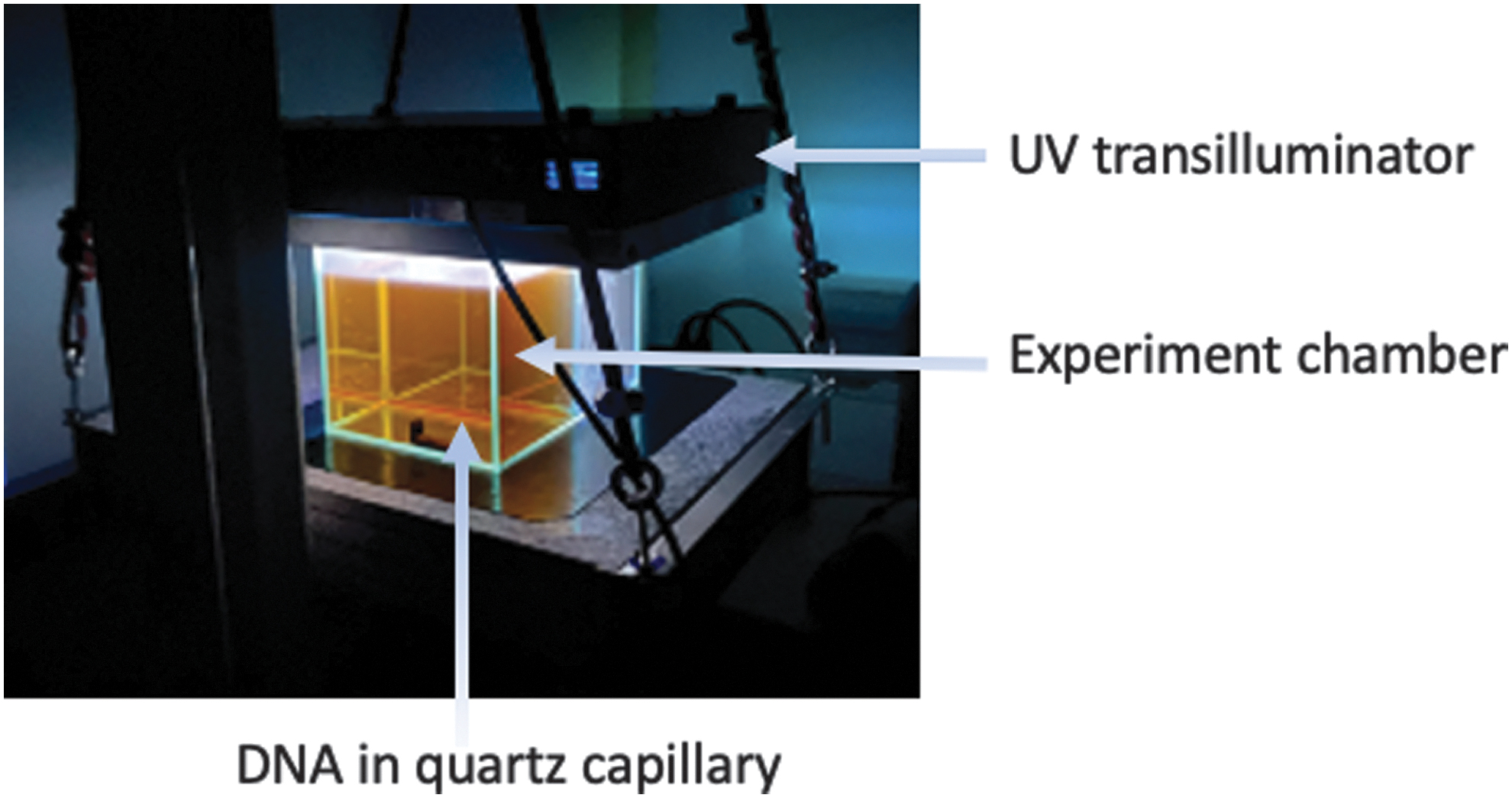
Experiment chamber used for DNA protection assay against UV. The chamber volume is 4.5 L with dimensions of 15 × 20 × 20 cm. DNA is sealed inside quartz capillary tubes and secured at the bottom of the tank. A UV transilluminator is placed in an inverted position such that UV can be irradiated directly through the chamber volume and on to the DNA samples in the capillary tubes. DNA conformation changes were assessed after UV was allowed to transmit through the chamber filled with air (empty chamber), water, silica solution, Fe3+ chloride, or a combined SiO2-Fe3+ solution.
The capillary tubes filled with DNA were submerged in the experimental chamber, which had been filled with air or water or silica solution (70 mM), or an Fe3+ chloride (0.55 mM) solution, or an SiO2-Fe3+ solution (70 and 0.55 mM, respectively).
UV exposure was performed by illuminating the experimental chamber with the UV transilluminator for a duration of time that ranged from 0 to 9 days. Thereafter, the DNA samples were carefully expelled from the capillary tubes into a separate sterile Eppendorf tube and analyzed for DNA damage.
2.3. DNA analyses
DNA integrity was determined by agarose-gel electrophoresis (AGE). The properties of different forms of DNA (topoisomers) were measured as a function of UV irradiation by comparing its electrophoretic mobility against a ladder of double-stranded DNA of known sizes. Three major topoisomers—supercoil, linear, and relaxed circular DNA—can be determined by measuring the DNA's electrophoretic mobility. Different conformations between these three major topoisomers can be observed by their migration rates in the gel. DNA damages that include covalent modification or strand breakage can render changes in the DNA conformation. Therefore, DNA modifications caused by UV exposure can be directly visualized when compared to that of a control. All AGE analyses were performed with 0.8% (w/v) agarose gel in 40 mM Tris acetate and 1 mM EDTA, pH 8.0 (TAE) running buffer, stained with GreenSafe Premium (NZYTech, Portugal), and visualized under UV illumination as described in standard molecular biology protocols (Lee et al., 2012).
2.4. Particle characterization
Colloidal suspensions formed after the mixing of sodium silicate, and ferric chloride solutions were centrifugated at 5000 rpm for 10 min. The collected sample was dried and analyzed by X-ray diffraction, using a high-resolution Bruker D8 Advance Series II X-ray diffractometer (at the Laboratorio de Estudios Cristalográficos) equipped with a Vario primary Ge (111) monochromator, which provides monochromatic Cu Kα1 radiation, and a Lynxeye PSD detector. Diffractograms were acquired in transmission mode, at 40 kV acceleration voltage and 40 mA current, with 2θ scans from 5° to 80° with a 2θ step of 0.02° s−1.
Raman spectra were recorded by using a JASCO NRS-5100 dispersive micro-Raman spectrometer equipped with a JASCO IRT-7100 microscope and an Andor DU 420 OE CCD detector. An excitation beam with a wavelength of 532 nm (frequency-doubled neodymium-doped yttrium aluminum garnet laser) and output power of 100 mW was used. Spectra were acquired with an exposure time of 5–10 s and accumulation of 10–60 times to improve the signal-to noise ratio. Fourier transform infrared (FTIR) spectra were recorded by using a JASCO 6200 spectrometer with Spectra Manager v2 software. Scanning electron microscopy was used to characterize the morphology and the local elemental composition of the precipitate with a Zeiss Supra 40VP field-emission scanning electron microscope (FESEM) equipped with an Oxford energy-dispersive X-ray analyzer (EDX) operating at 5–20 keV at the Center for Scientific Instrumentation (CIC) of the University of Granada (Spain). After complete desiccation, glass slides containing the precipitate were directly mounted on the SEM stub by using double-faced conductive carbon tape for electron microscopic analysis.
3. Results
The DNA protection assay was based on distinguishing significant topoisomeric changes of bacterial chromosomal DNA (plasmid DNA) after long exposure to UV in the presence and absence of water, silica, ferric chloride, or their combined mixture. Plasmid DNA has three principal topoisomers: supercoiled (SC), nicked circular (NC), and linear (L) based on their covalent modifications resulting from single- or double-strand cut. The topoisomers can be easily identified by their relative electrophoretic migration rate in an agarose gel. The migration profiles of plasmid topoisomers are well known (Hintermann et al., 1981). In short, the SC topoisomer represents the plasmid DNA form with minimal damage—there is no covalent modification or strand cuts. Thus, SC will be most compact and migrate the fastest. When the plasmid DNA undergo a single double-strand cut, the L form is produced, which gives rise to a slower migration rate that corresponds to its base pair length when compared to known molecular weight markers. Finally, if there is a single-strand cut, the NC form will be observed, resulting in a spread-out circle that will have the most drag in the agarose gel and will correspond to the slowest migrating band. This concept is schematized in Fig. 3.
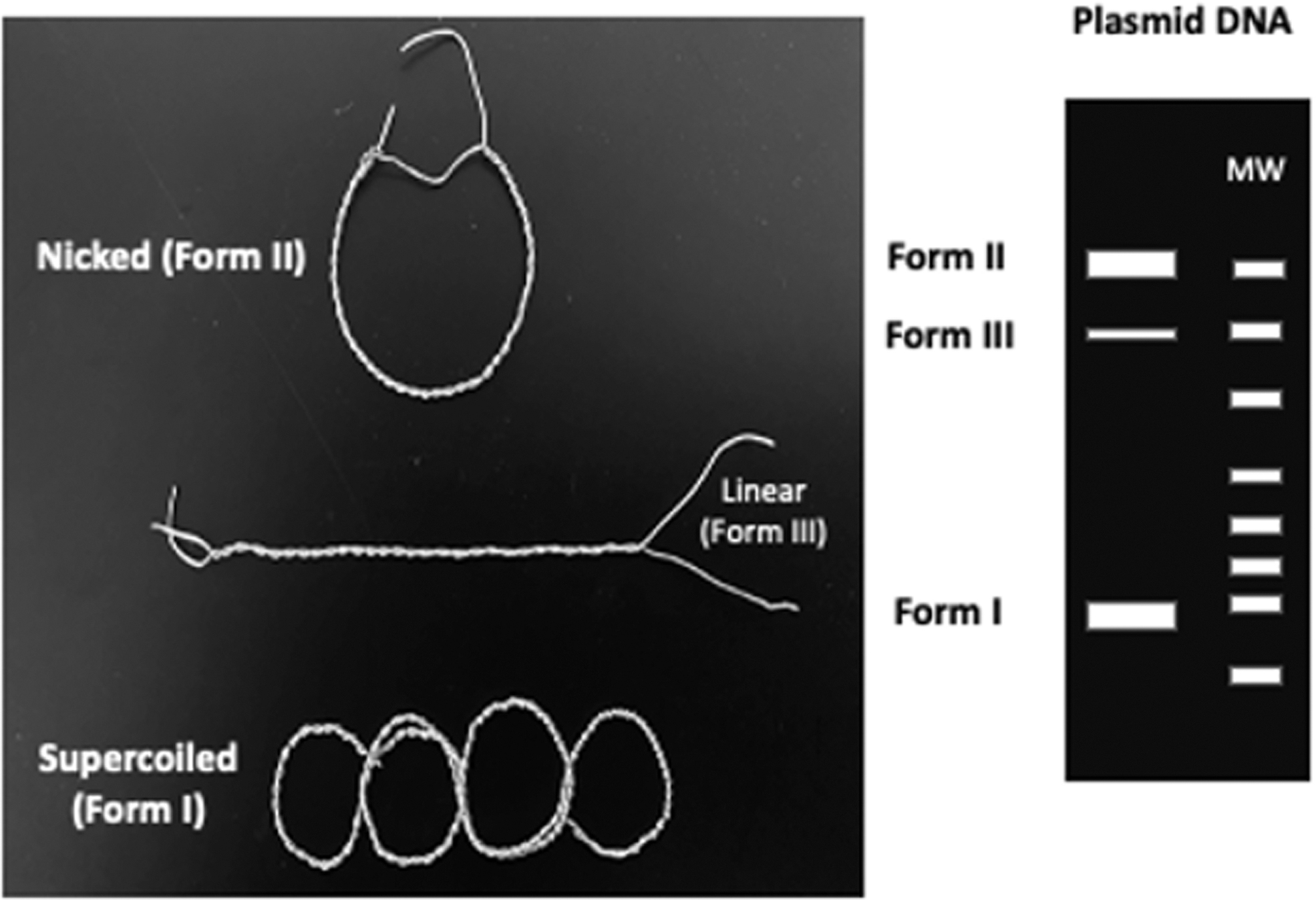
Schematic diagram of plasmid DNA topoisomers. Wired models of plasmid DNA topoisomers, NC, L, and SC shown as Forms II, III, and I, respectively (left panel). A virtual agarose gel profile is illustrated (right panel), showing the expected relative electrophoretic migration rates of the three major forms of plasmid DNA.
When bacterial plasmid DNA was exposed to UV radiation with just air space in the experiment chamber, significant changes in the DNA isomers were observed in as little as 5 min. Topoisomeric changes of plasmid DNA can be observed by agarose gel electrophoresis analysis (Fig. 4) where the progressive SC band shifts to the L form. UV radiation causes two types of lesions that include cyclobutene pyrimidine dimers and 6-4 photoproducts. These lesions distort the DNA structure subjecting it to formations of bends or kinks. In addition, UV radiation can elicit free-radical propagation that ultimately leads to covalent breakage (Rastogi et al., 2010). After only 25 min of UV exposure, the predominant DNA form was the linear isomer that is indicative of chemical sheering of the plasmid DNA.
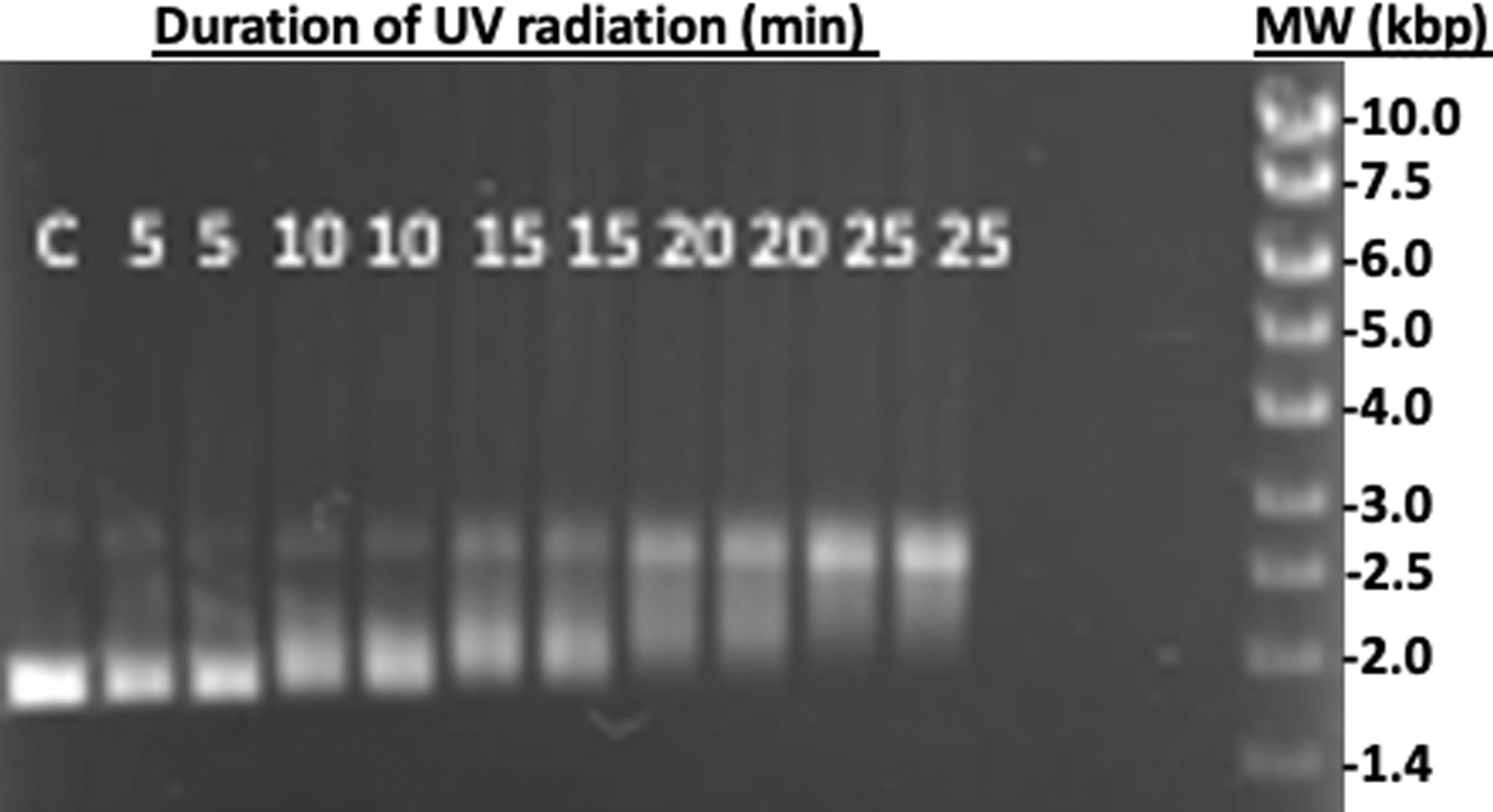
Topoisomeric changes of plasmid DNA as a function of UV radiation transmitting through air. Plasmid DNA was exposed to UV radiation for a duration of time between 0 and 25 min and analyzed by agarose gel electrophoresis. The DNA bands were visualized with GreenSafe Premium staining under UV illumination. Lane C is the control with 0 min UV exposure. The numbers on each lane indicate the exposure time in minutes. In exception to the control, all samples are shown in duplicates. Significant DNA form changes can be observed in about 10 min where the SC form progressively changes to the NC. In about 20 min of UV exposure, the predominant form is that of the linear, indicative of double-strand breaks.
When UV was radiated through water, there was a modest attenuation of UV radiation against the DNA substrate. As shown in Fig. 5, when the plasmid DNA was exposed to UV radiation for a duration of time between 0 and 60 min, there was a notable topoisomeric transformation from SC to NC (occurring near 20 min) and ultimately to a complete linear isomer by 60 min. The intensity of the SC band progressively became less, while the intensity of the bands corresponding to the other isomers increased proportionally. Here, the three major forms of plasmid DNA are visible with the SC and NC forms converging to the L form as a function of UV radiation.
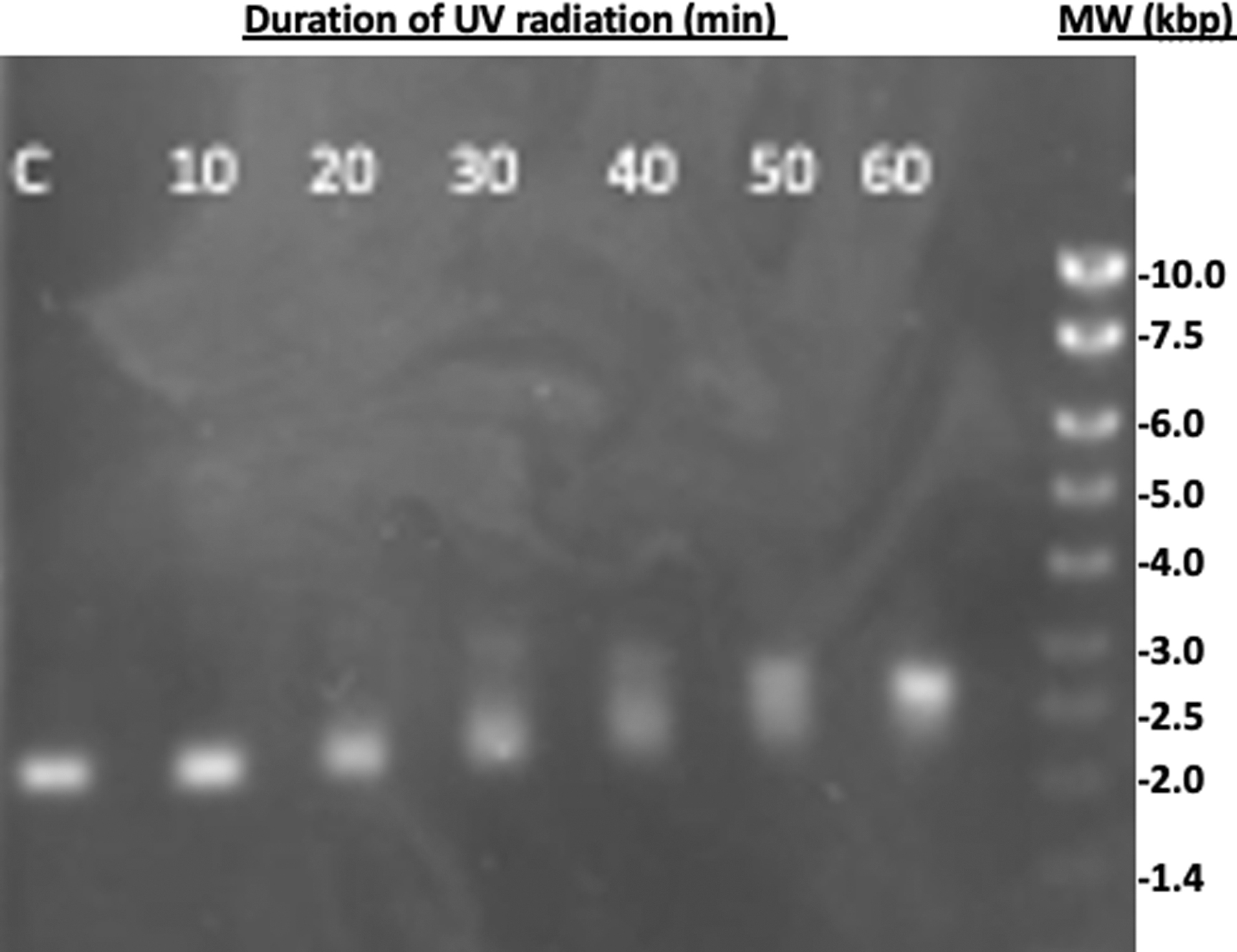
Topoisomeric changes of plasmid DNA as a function of UV radiation transmitting through water. Plasmid DNA was exposed to UV radiation for a duration of time between 0 and 60 min and analyzed by agarose gel electrophoresis. The DNA bands were visualized with GreenSafe Premium staining under UV illumination. Lane C is the control with 0 min UV exposure. The numbers on each lane indicate the exposure time in minutes. Topisomeric changes are not observable until 20 min of UV radiation. Complete linearization of the plasmid DNA appears between 50 and 60 min of exposure.
Topoisomeric changes of UV-radiated plasmid DNA were significantly diminished when shielded with silica solution (Fig. 6). The plasmid DNA was exposed to UV radiation for a duration of time between 0 and 180 min. There were no observable DNA conformation changes until after 60 min with the presence of the linear form (L). Even after 3 h of UV exposure, a significant amount of the SC isomer was retained. Sodium silicate concentration of 70 mM was sufficient to completely protect the DNA from any damage for as much as 1 h. The silica concentration is comparable to that measured in the water of Aqua de Ney spring (García-Ruiz et al., 2017).

Topoisomeric changes of UV-radiated plasmid DNA when shielded with silica solution. (
When ferric chloride (0.55 mM) was used to attenuate UV radiation, a resistance to topoisomeric changes was evident (Fig. 7). Plasmid DNA was exposed to UV radiation for a duration of time between 0 and 144 h (6 days) with little indication of DNA breakdown. In comparison to silica, Fe3+ attenuates UV drastically more, as no significant DNA conformation changes were observed until after 6 days of UV exposure where the predominant DNA form remained to be SC.
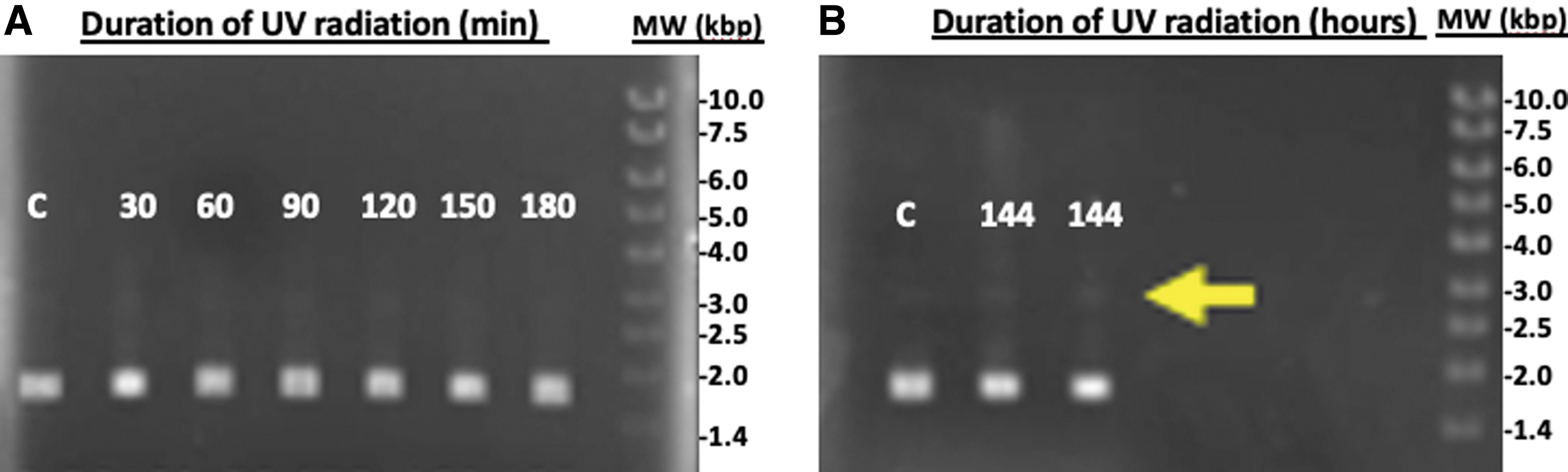
UV protection of plasmid DNA with ferric chloride solution. A resistance to topoisomeric changes of UV-radiated plasmid DNA are observed when shielded with the Fe3+ solution. (
When a combined solution mixture of silica (70 mM) and iron (0.55 mM) was tested for UV blockage against DNA damage, the SC DNA form was predominantly retained for up to 216 h (9 days) of UV radiation (Fig. 8). The silica-Fe3+ solution showed the most attenuation against UV compared to that of water, silica, or Fe3+ when they are individually prepared. The combination of silica and iron, under this study's setup, can extend the tolerance of UV exposure time associated with DNA damage to more than 2500-fold compared to that without any attenuation media (i.e., air).
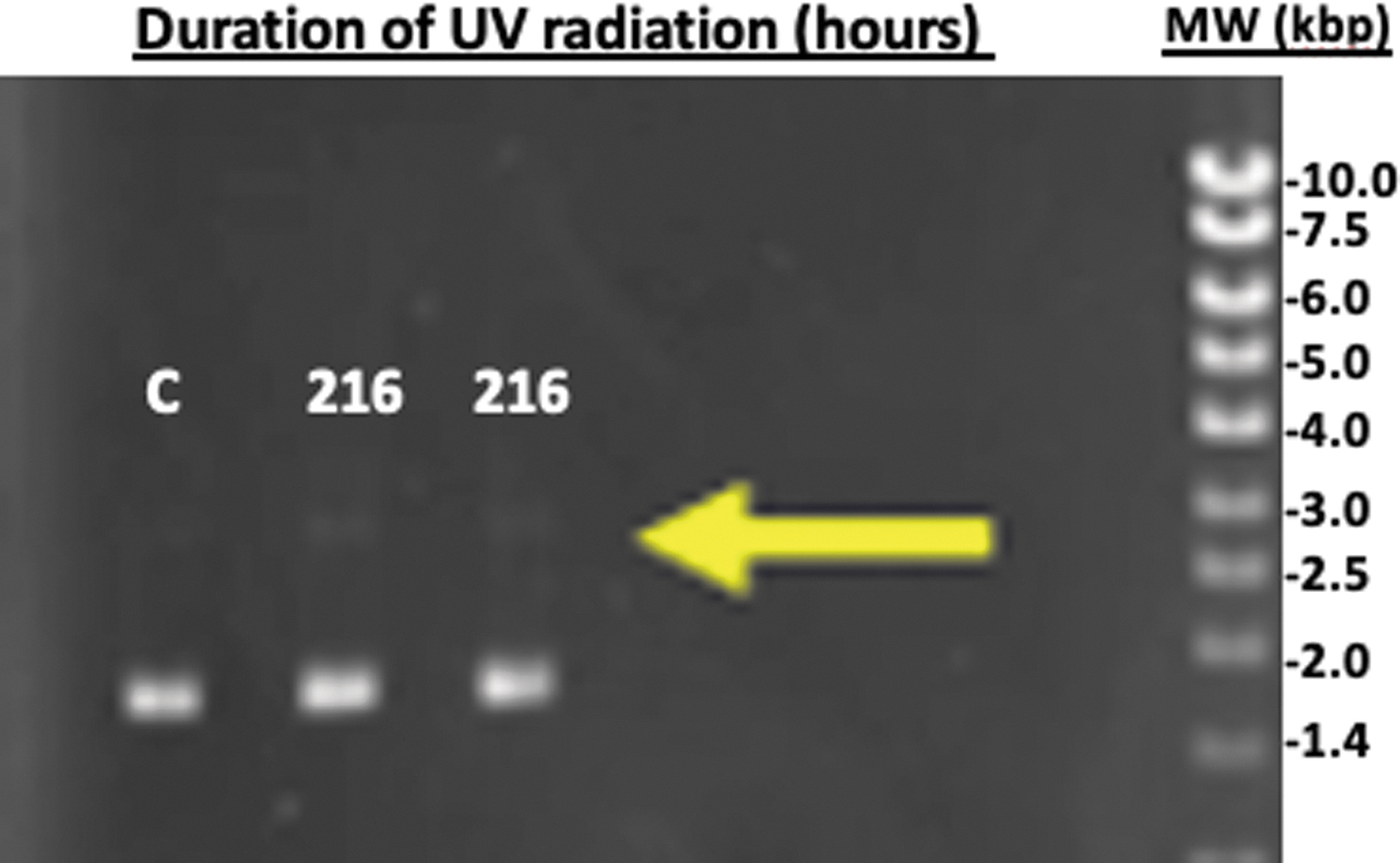
UV protection of plasmid DNA with silica-Fe3+ solution. Plasmid DNA was exposed to UV radiation for a duration of 216 h (9 days). Lane C is the control with 0 min UV exposure compared to DNA samples that have been radiated for 9 days. The predominant DNA form was SC showing small traces of the linear form (yellow arrow) after 9 days of UV exposure. All analyses were performed by agarose gel electrophoresis and visualized with GreenSafe Premium staining under UV illumination.
Throughout all the experiments reported here, the DNA solution was kept at pH 7.5 in the quartz capillary with a constant temperature of 20°C and pressure of 1 atm such that any conformational changes in the nucleic acid were attributed to only the UV radiation and not to pH variations or any other biophysical parameters in the nucleic acid sample. In regard to the chemical stability of iron and silica and their mixtures, we have also confirmed that the chemical composition responsible for attenuating the UV radiation was stable. In the case of the iron-silica solutions, the pH was measured to be constant at about 12 prior to the onset of UV exposure and at the end of each experiment. The longest duration was 9 days in which the pH remained at 12 throughout the experiment. When iron chloride and silica were mixed at pH 12, we observed the formation of a colloidal suspension of yellow/brown color in agreement with other published studies (Schwertmann et al., 1984), which remained stable for the entire duration of the experiment; there were no color changes, there was no precipitation, and there were no changes in pH as measured before and after each experiment. No solid precipitation was observed when viewed with the naked eye or under a light microscope.
To know the nature of the suspended particles, the samples collected at the beginning and at the end of the experiment were centrifuged, and the solid deposits were analyzed by X-ray diffraction with monochromatic Cu Kα1 radiation. As shown in Fig. 9B, the diffractograms do not show the diffraction characteristic of crystalline materials. The broadband observed at 2θ = 30° has been interpreted (e.g., Dyer, 2010) to correspond to amorphous silica, but it is most probably one of the bands of the disordered ferrihydrite (Cornell and Schwertmann, 2003). Our X-ray diffraction experiments do not show the presence of crystalline iron oxide-hydroxide particles, neither ferrihydrite, goethite, nor lepidocrocite. These results agree with those reported by Percak-Dennett et al. (2011), who found no significant crystalline mineral product peaks in the X-ray analysis of co-precipitates of Fe3+and silica prepared by hydrolysis of FeCl3 in high-silica solutions. Field emission scanning electron microscope and elemental X-ray analysis show that the ultra-centrifugated suspension is made of 40 nm spherical and spheroidal particles (Fig. 9A). Their atomic composition is 36.3% silica and iron at approximately 1:1 ratio, 53.3% oxygen, and 9.7% sodium. Noticeably, chlorine was not found. The collected colloid suspension was also examined with Raman and FTIR microspectroscopy. As shown in Fig. 9C–9D, both the Raman and FTIR spectra are dominated by the vibration mode of silanol Si-O-H and Si-O, groups mixed with vibration frequencies characteristic of Fe-OH.
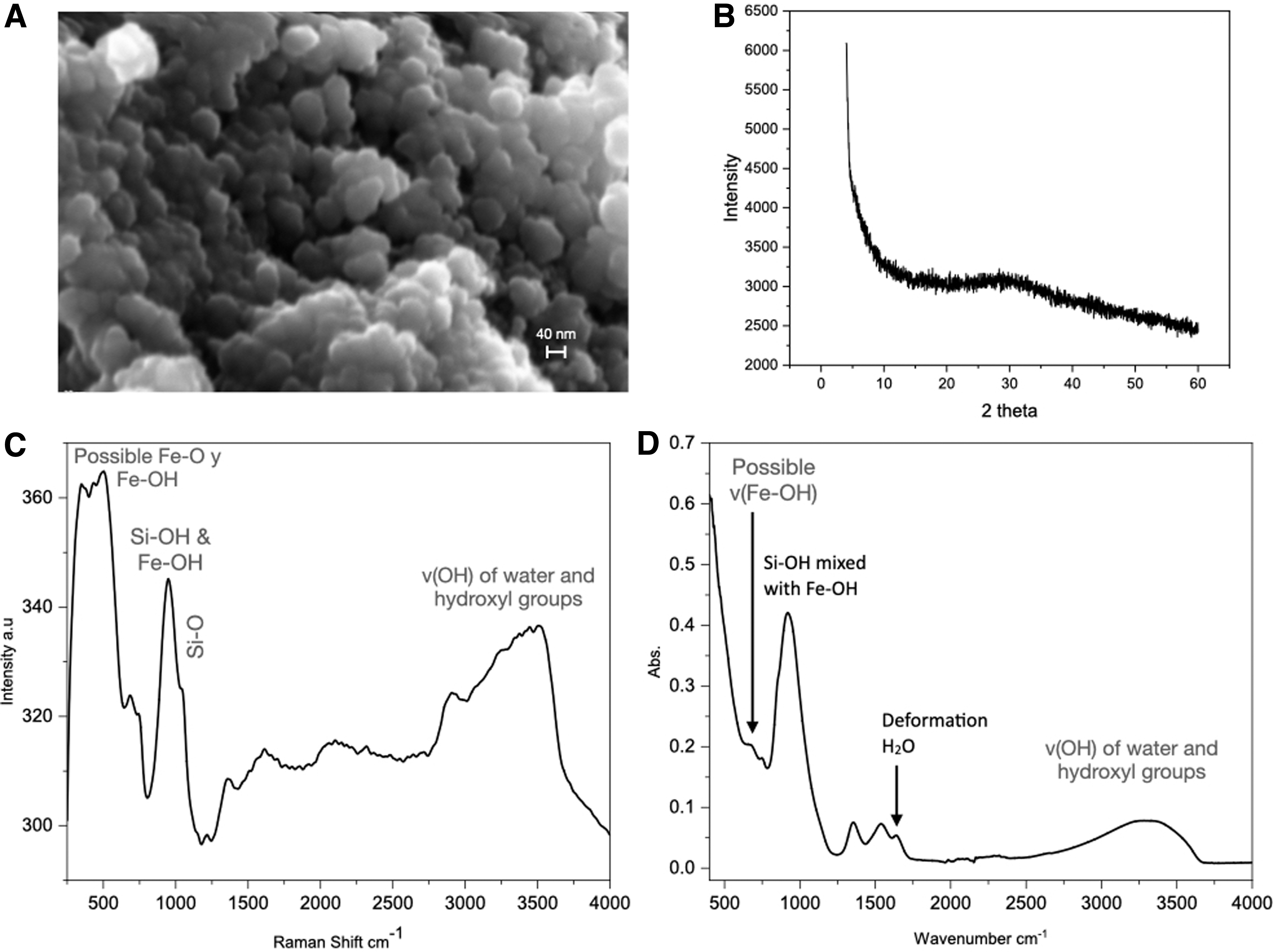
Properties of the colloidal suspension obtained after mixing sodium silicate and iron chloride solutions. Colloidal particles are observed by FESEM after centrifugation (
Taken together, our analyses suggest that the particles that shield intense UV radiation so efficiently are 40 nm colloidal particles of silica gel embedding Fe3+ ions that may be adsorbed by silanol groups or forming tiny disorganized Fe3+ oxyhydroxide nanoparticles, without crystalline order. This ability of silica to inhibit the crystallization of ferrihydrite and its further conversion to lepidocrocite or goethite by chemisorption (Schwertmann et al., 1984; Jones et al., 2009; Cismasu et al., 2014) or upon the formation Fe-Si complexes (Rzepa et al., 2016; Schulz et al., 2022) has been many times proposed in the literature (Carlson and Schwertmann, 1981; Dyer, 2010; Percak-Dennett et al., 2011; Wu et al., 2011).
The iron solution at 0.55 mM had a measured pH of 3. As mentioned previously, the iron concentration in our experiments was based on reported estimates of iron content in the Archean ocean, between 2 and 50 ppm (0.04 and 0.90 mM) (Johnson et al., 2008; Mloszewska et al., 2018). Under this condition, Fe3+ chloride is a stable solution. Previous experiments titrated with NaOH to pH 12 resulted in the crystallization of iron hydroxide and oxide particles that drastically affected the screening experiments. Therefore, we decided to perform this part of the experiment with stable Fe3+ chloride solutions to compare with that of soluble iron-silica solution experiments, the focus and core of this investigation. In regard to silica, the solubility of silica increases exponentially with pH from 180 ppm at pH 8.3 to more than 4000 ppm at pH 12 (Schenk and Weber, 1968; Iler, 1979; García-Ruiz et al., 2020). At the pH range of our experiments, the silica solution is fully stable as shown in previous studies (García-Ruiz et al., 2020).
4. Discussion
We report the effective shielding properties of silica and ferric iron against UV radiation to prevent DNA damage. In particular, colloidal suspensions of silica and Fe3+ show exceptional UV attenuation. These suspensions are formed by colloidal particles of ca. 40 nm in size embedding Fe3+ ions adsorbed on silanol groups that may also form small nanoclusters of iron hydroxide. Our results clearly show that crystalline precipitates of neither iron hydroxides nor iron oxides are required to achieve effective UV shielding. These findings have significant implications for our understanding of early Earth chemistry and its transition to early biological processes because iron and silica solutions were universally present in Hadean and Archean times. The geological record of the Archean period suggests that upwelled ferrous iron changed readily to ferric iron in the presence of alkaline water, and this mixture provided UV attenuation (Holland, 1984; Olson and Pierson, 1986; Pierson et al., 1993; Cockell, 1998, 2000; Garcia-Pichel, 1998).
UV photons have wavelengths that fall between visible light and gamma radiation. UV energy can be subdivided into UVA, UVB, and UVC components based on electrophysical properties, with UVC photons having the shortest wavelengths (100–280 nm) and the highest energy. During the Archean period, the three UV types were present, but the most harmful type to organic and biological molecules was UVC. The intensity of UVC radiation (200–280 nm) on Archean Earth was 310 μW/cm2. Currently, this value is near zero due to the ozone layer (Cockell, 1998, 1999, 2000). The surface radiation levels on Archean Earth were several orders of magnitude higher in the 200–300 nm wavelength range than those of today. It can be said that any form of life that might have been present on Earth's surface 4–3.5 billion years ago must have been exposed to a much higher dose of damaging radiation compared to the present day (Cnossen et al., 2007). Considering that the UV illumination used in this work had a light intensity 15 times higher than that reported for Archean Earth (Cockell, 1998), Fe and silica were demonstrated to still contribute significantly to the protection of DNA against UV radiation.
Other UV-absorbers have been proposed for enabling terrestrial prebiotic chemistry and early life formation. These include the fractal size distribution of organic compound clusters formed by photolysis of CH4 (Pavlov et al., 2001; Kasting, 2005; Wolf and Toon, 2010). The methane-rich atmosphere may have been important in the earliest Hadean, but it is unlikely that such a reducing atmosphere was continuously present when microbial life initially started to colonize the planet. Episodic global atmospheric UV screening during the Hadean and Archean periods has also been proposed to be caused by an organic haze with varying concentrations and particle sizes (Sagan and Chyba, 1997). The existence of banded-iron and silica formations in early Earth during the Archean era (Precambrian up to 2.5 Ga) with their UV-shielding capacity has rendered silica and iron to be significant protectors of UV radiation against early life-forms. Iron screening effects against solar UV have been observed in present terrestrial aquatic extreme environments (Pierson et al., 1993; Mloszewska et al., 2018). Archean bacteria mineralized in an iron-silica solution have been shown to have marked resistance to UV compared to nonmineralized bacteria (Phoenix et al., 2001). While early life-forms needed an environment capable of absorbing UV radiation, even more essential was that environment to protect the early macromolecules resulting from prebiotic chemistry. The stability of a chromosomal DNA molecule against UV radiation in the presence of iron and silica may lend support as to how macromolecules endured early Earth environments and engendered early molecular survival against UV radiation.
The concentrations of Fe3+ and silica used in this study were consistent with those found on early Earth and in extant alkaline environments such as at Lake Nye, California. Mloszewska et al. (2018) suggested that the direct contact between silica/Fe3+ with cyanobacteria produces a kind of layer that allows the system to be reproduced. In our investigation, circular plasmid DNA served as a model for a primitive chromosome or genetic material that, when exposed to damaging UV radiation, mimics geochemical reactions that occurred on early Earth. High concentrations of silica and ferrous iron converted to ferric species in the ocean water column, as well as in terrestrial hot spring environments, would have greatly contributed to a UV screening mechanism that protected complex prebiotic molecules as well as the earliest forms of life. These are geologically plausible scenarios that are directly linked to hydrothermal alteration of an ultramafic planetary surface. The UV-attenuating properties of both silica and iron may be an essential connection between the occurrences in prebiotic chemistry and the formation of the biological world, an issue that is pertinent to the origin of life.
In general, silica and iron mixtures that screen UV radiation could be an important prerequisite for habitability on other Earth-like rocky planets in the Universe (Craven et al., 2021). Many new exoplanets are being discovered every day, and harmful radiation from their respective stars is likely to engender UV-screening for the development of life (Cockell, 1999). Water-rock interaction and the resulting enrichment of a water body with silica and iron, therefore, will constitute another important basic factor in the definition of habitability.
5. Conclusions
We have experimentally demonstrated that alkaline silica- and iron-rich aqueous solutions and their mixtures are very efficient UV-attenuators. Aquatic alkaline environments enriched with silica and iron in small and large bodies of water may have acted as UV shelters for prebiotic processes in the early Earth environment. These aquatic alkaline environments would have been a perfect niche for the following: (a) protecting chemical reaction pathways that led to the synthesis of prebiotic molecules during the early stages of water condensation on the surface of Earth during the Hadean and Archean periods; (b) preserving prebiotic compounds of increasing complexity and self-organization that led to life as we know it; and (c) preserving the earliest forms of life based on carbon (bio)chemistry. These iron/silica-rich aquatic niches were most likely widespread on early Earth, but they may also exist on other Earth-like planets and moons.
Footnotes
Acknowledgments
The authors acknowledge Prof. Fernando Rull from the Universidad of Valladolid for the helpful contribution to the analysis of Raman and FTIR spectra. They also acknowledge Dr. Isabel Lopez-Guerra and Bendición Funes Martin from the Center of Scientific Instrumentation (CIC) of the University of Granada, Dr. Cristobal Verdugo from Laboratorio de Estudios Cristalográficos for technical help, and Ms. Antonia Sánchez-Marín for the English revision of an earlier version of the manuscript. The authors thank the European Research Council under the European Union's Seventh Framework Program (FP7/2007-2013)/ERC grant agreement no. 340863. J.M.G-R. and L.A.G-R acknowledge the Spanish Ministerio de Ciencia e Innovación for financial support to the project PID2020-112986GB-100 and Junta de Andalucía for financing the project P18-FR-5008. A.M. acknowledges DGAPA-UNAM (Mexico) for the support during a sabbatical year at the University of Granada.
Abbreviations Used
Associate Editor: Nita Sahai