
Abstract
Active hot springs are dynamic geobiologically active environments. Heat- and element-enriched fluids form hot spring sinter deposits that are inhabited by microbial and macroscopic eukaryotic communities, but it is unclear how variable heat, fluid circulation, and mineralization within hot spring systems affect the preservation of organic matter in sinters. We present geological, petrographic, and organic geochemical data from fossilized hot spring sinters (<13 Ka) from three distinct hot spring fields of Yellowstone National Park. The aims of this study were to examine the preservation of hydrocarbons and discern whether the hydrocarbons in these samples were derived from in situ communities or transported by hydrothermal fluids. Organic geochemistry reveals the presence of n-alkanes, methylalkanes, hopanes, and other terpanes, and the distribution of methylheptadecanes is compared to published observations of community composition in extant hot springs with similar geochemistry. Unexpectedly, hopanes have a thermally mature signal, and Raman spectroscopy confirms that the kerogen in some samples has nearly reached the oil window, despite never having been buried. Our results suggest that organic matter maturation occurred through below-surface processes in the hotter, deeper parts of the hydrothermal system and that this exogenous material was then transported and emplaced within the sinter.
Introduction
Siliceous hot spring sinters are archives of geobiology, as sinter deposits precipitating from hot spring effluents rapidly trap and entomb in situ evidence of both microbial and macroscopic eukaryotic activity (Walter et al., 1996; Hurwitz and Lowenstern, 2014; Campbell et al., 2015). However, the detailed mechanisms of organic matter (OM) preservation in hot spring deposits are not well understood, in particular with regard to whether the OM recovered from sinter deposits is derived from the syndepositional microbial or eukaryotic community that inhabited the hot spring as the sinter was deposited or whether some or all of the OM is exogenous and introduced by the recirculation of hydrothermal fluids as the hot spring system evolved.
To date, most studies have focused on the preservation of microbial fabrics and their OM remnants in active hot springs and documented a wide range of microbial communities that vary depending on the temperature and chemistry (including pH) of individual hot spring pools (e.g., Shiea et al., 1990; Jahnke et al., 2004; Pancost et al., 2006).
However, detailed studies of the preservation of hydrocarbons in fossilized hot spring sinters are much less common (Kaur et al., 2008; Gibson et al., 2014; Campbell et al., 2015; Reinhardt et al., 2019; Teece et al., 2020, 2022). As more ancient fossilized hot spring environments are recognized on Earth (Djokic et al., 2017, 2021) and Mars (Ruff and Farmer, 2016; Ruff et al., 2020), the need to study modern analogs becomes more pressing to fully understand the mechanisms of hydrocarbon preservation and migration throughout the diagenetic history of sinter deposit, as hydrocarbons are the most likely portion of the extractable OM to survive from the ancient Earth.
The preservation mechanisms of OM in modern and ancient hot springs are complex as high amounts of heat promote fluid circulation, which can transport OM far from its source and potentially even heat OM derived from indigenous microbial communities to well above their temperature of habitation in fluids that are recirculated down through the crust. This mechanism, which was reported from active hot springs in Yellowstone (Gonsior et al., 2018), Pleistocene samples from Kenya (Reinhardt et al., 2019), and fossilized hot springs in New Zealand (Teece et al., 2020), indicates potentially complex processes of OM formation, mobilization, transport, and preservation. Such a model is supported in deep time by the study of Duda et al. (2018), who provided evidence for the downward transport of OM into fossilized hydrothermal pathways through hydrothermal pumping in a 3.5 Ga old hydrothermal setting.
The question arises, therefore, as to whether the maturation and redistribution of hydrocarbons is a common phenomenon in siliceous hot spring sinters. Knowledge about such a process is critical for understanding the syndepositional and perhaps subsurface microbial populations in ancient hot spring deposits, and for interpreting the variety of fabrics and element enrichments formed by different types of microbial communities. Documenting this process is also critical for determining whether hydrocarbons are syndepositional or exogenous to rocks that may preserve signs of life, or whether the hydrocarbon signal may even be abiotic in origin (e.g., Duda et al., 2018).
In this study, we explore these questions through examination of fossilized sinters from three distinct areas of Yellowstone National Park. Specifically, we use contextual data of samples, microbial and rock textural morphology, petrography, scanning electron microscopy (SEM), Raman spectroscopy, and gas chromatography-mass spectrometry (GC-MS) to help elucidate whether, and to what degree, the hydrocarbon fraction of OM found within Yellowstone siliceous sinter deposits formed in situ—that is, autochthonous material from syndepositional microbial populations—or was allochthonous material introduced by fluid transport and recirculation within the active hot spring system.
Here, we consider a critical first step as to whether an organic biosignature within a sinter is syndepositional or an artifact of hydrothermal overprinting. To evaluate these questions, we combined the above techniques to assess the organic geochemistry in combination with the geological and biological contexts for these signals. This approach is essential to our understanding of organic biosignatures of the early Earth and the martian rock record.
Geological setting
Yellowstone National Park in Wyoming, United States, is a 50 × 80 km diameter volcanic caldera basin that formed from hot-spot volcanic activity associated with a hot lower-mantle plume beneath the North American Plate (Hurwitz and Lowenstern, 2014). The related heat transfer to upper crustal levels promoted the formation of thousands of chemically diverse hot springs, which formed after the end of the last glaciation, making this the greatest concentration of hydrothermal surface features in the world (Licciardi and Pierce, 2018). The sampling sites for this work were fossilized sinter deposits in three separate hydrothermal areas as follows: Norris Geyser Basin (NGB), Lower Geyser Basin (LGB), and Midway Geyser Basin (MGB) (Fig. 1). Below, we present sample information.
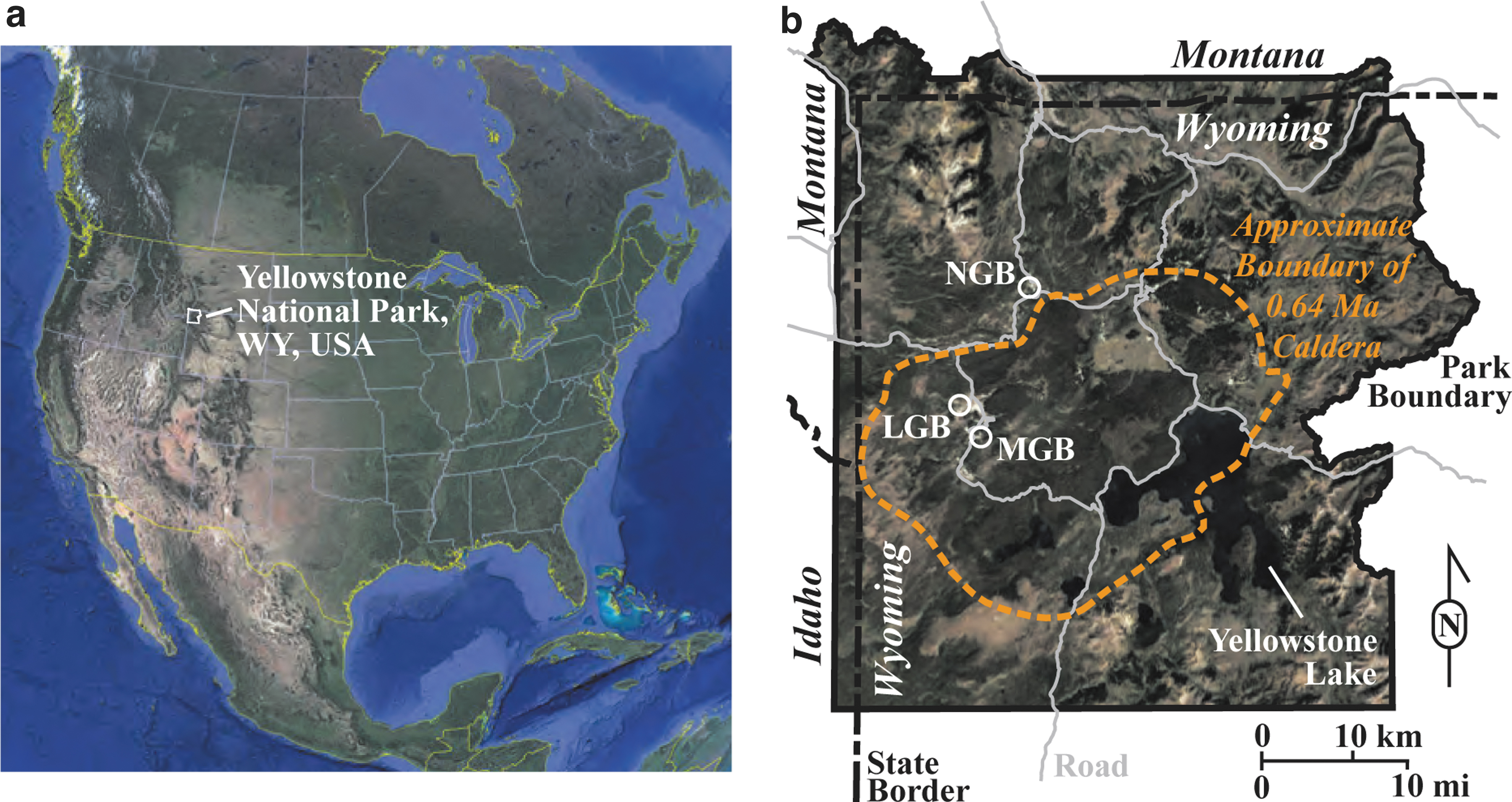
The NGB is located ∼5 km north of the 640 Ka Yellowstone caldera rim (Fig. 1) and is underlain by the A and B members of the Lava Creek Tuff, a gray, brown, or pink ash-flow tuff that is densely welded and devitrified (Christiansen, 1975). The Gap area within the NGB (Supplementary Fig. S1) is composed of stream gravels and hot spring siliceous sinter deposits.
The Gap area is bound to the south-southeast by hydrothermally cemented kame deposits from the Bull Lake and Pinedale Glaciations (ended ∼140 and ∼13 Ka, respectively) and to the north-northwest by Pinedale glacial till, Bull Lake hydrothermally cemented kame deposits, chalcedonic sinter (diagenetically altered Opal A), and exposures of Member B of the Lava Creek Tuff (Waldrop and Pierce, 1975; Licciardi and Pierce, 2018). The Bull Lake hydrothermally cemented kame deposits are light gray to brownish gray poorly sorted, crudely to moderately well-bedded sandy gravels deposited at ice boundaries.
The ‘Has Been Hot Spring’ of the Gap Area is an extinct hot spring that retains the original hot spring bowl (∼8 m diameter), which is filled with siliceous sediment (sands and clays) but has half of the original (northwestern/down-slope) rampart exposed. The original hot spring likely post-dates the end of the Pinedale Glaciation (<13 Ka) as it exhibits textures characteristic of subaerial precipitation (see textural descriptions in Section 3.1). Although age-dating has not provided conclusive data, erosional evidence indicates that the hot spring is at most several thousand years old. Texturally, sinters at ‘Has Been Hot Spring’ most closely resemble those at Dante's Inferno in Yellowstone (Sylvan Spring Area, Gibbon Geyser Basin). Thus, the ‘Has Been Hot Spring’ likely had similar geochemistry to that of Dante's Inferno (T = 79.4°C, pH = 5.19), as described most recently by Havig and Hamilton (2019).
Two samples (HB C and HB E) were collected from the ‘Has Been Hot Spring’ (Fig. 2a, b; 44°43′41.8"N, 110°42′48.9″W). A third sample, ‘Biffy’ (Fig. 2c; 44°43′35.7″N, 110°42′33.2″W), comes from an extinct Fe-Si spring (Supplementary Fig. S1) that is close to two active springs named Perpetual Spouter (T = 87.3°C, pH = 7.06) and Red Bubbler (T = 91.8°C, pH = 2.85), which may be modern expressions of the same hydrothermal source that fed the original ‘Biffy’ source spring. The difference in pH between these two extant springs presumably relates to subsurface boiling and phase separation of the fluid and steam, resulting in the predominantly acidic vapor phase in Red Bubbler versus the predominantly neutral to alkaline liquid phase in Perpetual Spouter (Havig et al., 2021).

Photographs of the Yellowstone samples studied in this article:
Steep Cone (SC; 44°33′59.9"N 110°51′47.6"W) is a 30 m diameter, 6.6 m high, long-lived, and still active siliceous sinter mound (Supplementary Fig. S2) located in the Sentinel Meadows area of the northern part of the LGB. This area was deglaciated at the end of the Pinedale glaciation c, 13–14 Ka (Pierce, 2003). Sentinel Meadows is likely underlain by the 160 Ka Nez Pierce Creek rhyolite flow and is bound to the north by a flow front of the 108 Ka West Yellowstone rhyolite flow and to the south by poorly sorted kame deposits (Christiansen and Blank, 1974; Waldrop and Pierce, 1975; Christiansen, 2001). The meadow surrounding the SC silica mound is composed of glacial till deposited during the Pinedale glaciation, humic-rich alluvium, diatomaceous silt, and hydrothermal siliceous sinter deposits (Supplementary Fig. S2) (Waldrop and Pierce, 1975).
The source pool of SC is located on top of the sinter mound. It is 2.5–3 m in diameter and >9 m deep. Hot, circum-neutral water (T = 93°C, pH = 7.6; Havig et al., 2021) overflows from the pool and down the sides of the sinter mound, precipitating silica as it cools (Inagaki et al., 2001). We assume that precipitation of the SC subaerial siliceous sinter was initiated after the catastrophic draining of a glacially dammed lake that filled the LGB and MGB at the end of the Pinedale glaciation (c. 13–14 Ka), and that SC has been a continuously active hot spring ever since.
This assumption is based on a continuous sequence of subaerial siliceous sinter deposits that are exposed on a cut section of the cone created by incision of Sentinel Creek (Supplementary Fig. S2). These exposed deposits lack any obvious unconformity in the exposed sinter stratigraphy. Three microbial siliceous sinter samples were taken from the exposed inner stratigraphy of SC: SC Low, SC Mid, and SC High (Fig. 2d–f).
Anatomy Lab, Rabbit Creek, MGB
The MGB is located at the northwest end of the Mallard Lake resurgent dome within the Yellowstone Caldera (44°31′01.4"N 110°48′22.7"W). The Rabbit Creek Area within this basin is bounded by the Biscuit Basin rhyolite flow (black perlitic vitrophyre of flow-layered or flow-brecciated rhyolite, ∼516 Ka) to the northwest and the Mallard Lake rhyolite flow (large gray-pink-brown rhyolite flow with quartz and sanidine phenocrysts, ∼151 Ka) to the east and south (Christiansen and Blank, 1974; Christiansen, 2001). The Rabbit Creek Area is characterized by extensive amorphous siliceous sinter deposits that are 0.6–6.0 m thick and overlie Pinedale Glaciation glacial tills and kame deposits. Anatomy Lab (AL) (Supplementary Fig. S3) is located on kame deposits that we interpret as representing a glacial lake terrace that formed near the end of the Pinedale Glaciation.
It is our interpretation that this glacial lake was connected to the larger glacial lake that filled the LGB at the end of the Pinedale Glaciation, which then catastrophically drained following the failure of a glacial ice dam, resulting in the series of hydrothermal eruptions found in the LGB. With the dramatic lowering of the water table in the MGB, AL lost its original hydrothermal source, drained, and lost the source water that originally maintained the large pool and outflow.
AL is a hot spring that does not currently discharge hydrothermal fluids, but has hydrothermal fluids deep (∼3 m) within the source conduit of the spring (T = 60.9°C, pH = 6.3) (Supplementary Fig. S3). AL is a roughly rectangular basin or bowl (∼17 × 17 m) that remains largely free of infilling sediment. In lieu of effective age dating, we assume that the sinters deposited in AL are likely c. 13–14 Ka, based on the end of the Pinedale Glaciation and before the source conduit was shut off due to catastrophic draining of the glacial lake described above. Three samples were collected from AL (Supplementary Fig. S3). One sample was split into two subsamples, which are referred to as AL Fil and AL Base (Fig. 2g). The other samples are AL Strom and AL Cement (Fig. 2h, i).
Materials and Methods
Preparation
The samples in this study were broken off from the outcrops by hand and immediately placed in sealed plastic bags until analysis. To mitigate contamination from the sampling stage (i.e., from hands or the plastic bags), the outer 5–10 mm of all samples were removed by sawing with a diamond blade. Before sawing, the blade was cleaned by repeated sonication in dichloromethane-methanol (DCM-MeOH, 9:1 v/v), followed by repeated heating to 300°C for 4 h. If sufficient in size, the offcuts were prepared into thin sections, as required for petrographic and mineralogical analyses. AL Fil and AL Base were separately processed for organic geochemical analyses. All the samples were analyzed for total carbon on a Vario MICRO cube elemental analyzer from Elementar at the Macquarie University Chemical Analysis and Biotechnology Centre.
Optical microscopy and SEM
Fresh faces of sample offcuts were imaged by optical microscopy on a Leica DM2500P microscope at the University of New South Wales, Australia. SEM was carried out with a Hitachi TM-1000 (Hitachi High-Technologies Corporation) installed at the Continental Science Drilling Facility (CSD) within the Department of Earth and Environmental Sciences at the University of Minnesota. Backscattered electron images were carried out by using 15 kV acceleration voltage and 35–55 pA. The chamber vacuum was held between 30 and 50 Pa.
Raman spectroscopy
Raman spectroscopy analysis was carried out with a Horiba LabRam HR Evolution instrument installed at CSIRO Energy (Kensington, Australia). The Horiba instrument is equipped with a multichannel air cooled (−70°C) Synapse Visible detector, and a 100 mW continuous wave (532 nm) diode laser from Laser Quantum. For OM analysis, filtering decreased the laser power to ≤0.5–1.0 mW, which neither damaged the samples nor affected the acquired spectra by thermal degradation. Spot analyses were conducted with a 100 × objective, which equates to laser focusing to ∼0.72 μm beam width.
The spectra were collected with 600 grooves/mm in the ∼200–2000 cm−1 region, using 1-s exposure time and 300–500 accumulations. The spectra were calibrated against the peaks of a Si wafer. Baseline correction, which was done in the software MagicPlot, involved baseline fitting through spectral intervals containing background only. The putative vibration modes of OM were deconvoluted by using Gaussian peak functions in the ∼1050–1750 cm−1 region.
Organic geochemistry
All solvents and glassware were first checked for purity with a solvent blank by GC-MS, as described below. Glassware was cleaned in a Decon90 solution, washed with Millipore water, and combusted in an oven at 500°C for 8 h before use. Aluminum foil was ashed at 500°C for 8 h before use. Glassware was stoppered or covered with foil to prevent laboratory contamination. The rotary evaporator and accelerated solvent extractor were avoided as they are potential sources of cross-contamination. Instead, samples were solvent extracted by sonication and evaporated using a hot plate and gentle blowing under a stream of dried nitrogen. Care was taken to avoid evaporation of samples to dryness because this can remove low molecular weight compounds (Ahmed and George, 2004). Blanks were utilized to detect possible contaminants introduced during laboratory procedures.
Rocks were crushed with a tungsten mill, which was cleaned by MeOH between each crush. The weights of the rock powders are provided in Table 1. After crushing, the rock powder was placed in beakers and extracted by ultrasonication in DCM-MeOH, 9:1 v/v. The powder was sonicated in two cycles, each 10 min long, with a 5-min rest in between, during which the sample was stirred, before being decanted. This process was repeated three times per sample. The second and third sonication mixes were each added to the first. The samples were then evaporated to a minimum volume.
Hydrocarbon Ratios for the Yellowstone Samples
Hydrocarbon Ratios for the Yellowstone Samples
CPI22–32 = carbon preference index of n-alkanes: (2 × × (C23 + C25 + C27 + C29 + C31))/(C22 + 2 × C24 + 2 × C26 + 2 × C28 + 2 × C30 + C32); CPI16–26 = carbon preference index of n-alkanes: (2 × × (C17 + C19 + C21 + C23 + C25))/(C16 + 2 × C18 + 2 × C20 + 2 × C22 + 2 × C24 + C26).
AL = Anatomy Lab; HB = ‘Has Been Hot Spring’; nd = not determined; Ph = phytane; Pr = pristane; SC = Steep Cone; Tm = C27 17α-trisnorhopane; Ts = C27 18α-trisnorneohopane.
The extractable OM was fractionated on small scale silica columns in Pasteur pipettes, using a mixture of n-hexane and DCM (4:1 v/v, 3 mL) to elute an apolar fraction. The apolar fraction was further fractionated on a new silica column into aliphatic and aromatic hydrocarbon fractions using n-hexane (2.6 mL) and the 4:1 solution (3.5 mL). The solvent of each fraction was reduced to a minimum volume, and a standard solution of about 117 ng each of anthracene-d10 (98 atom%D, Isotec), p-terphenyl-d14 (98 atom %D, Isotec), and tetracosane-d50 (98 atom %D, Isotec) was added to each fraction. The fractions were then further evaporated to <100 μm and pipetted into a 100 μL vial insert.
The aliphatic and aromatic hydrocarbon fractions were analyzed by GC-MS on an Agilent gas chromatograph (6890N) coupled to an Agilent mass selective detector (5975B) in the organic geochemistry laboratory at Macquarie University. A 1 μL aliquot of each fraction was injected into a programmable temperature vaporization inlet operating in splitless mode with a J&W DB5MS capillary column (length 60 m, inner diameter 0.25 mm, and film thickness 0.25 mm).
The carrier gas was helium (1.5 mL/min., constant flow), and the temperature of the GC oven was ramped from 40°C (2 min. isothermal) to 310°C (30 min isothermal) at a rate of 4°C/min. The electron ionization source was operated at 70 eV. The MS data were acquired in a full scan (mass-to-charge ratio [m/z] 50–550) and single-ion monitoring (SIM) modes (for a list of SIM ions monitored see Supplementary Table S1). Compound identification was based on mass spectral data and retention time comparison with a North Sea oil standard (Weiss et al., 2000). Semiquantitative analyses of SIM data were performed using the internal standards. The limit of detection was 0.5 pg/g for n-alkanes, and 6 pg/g for hopanes and terpanes.
Results
Morphology and petrography
At a macroscopic scale, the hand sample HB C (Fig. 2a) from the ‘Has Been Hot Spring’ of the NGB appears to be well silicified, with only a few localized aggregations of 1 mm wide pores. Sample HB C is thinly laminated with alternating orange-to-brown and lighter colored laminates. In thin section, HB C (Fig. 3a) contains vertically elongated fenestrae, up to 1–mm long, within structureless opaline silica. The laminated texture is typical of thinly laminated microfacies, which can be associated with the palisade microfacies in sinters (Campbell et al., 2001, 2015; Guido and Campbell, 2011). In both microfacies, the micropillar structures are inferred to be silica-coated microbial filaments. Thinly laminated microfacies have been reported from modern to Paleozoic sinters (Walter et al., 1998; Campbell et al., 2015).

Plane polarized light thin-section photomicrographs of selected Yellowstone samples.
Hand sample HB E (Fig. 2b) from the ‘Has Been Hot Spring’ of the NGB is brown to beige colored and is roughly textured. Two well-defined horizons can be observed at a macroscopic scale. The 5 cm thick basal horizon has a high primary porosity (∼30%) with voids ranging between 0.5 mm to a few mm in width. Long (up to 5 cm long), thin, and subhorizontally oriented broken sheets of silica are embedded in the porous silica matrix. Overlying the basal horizon, a 9 cm thick horizon is characterized by a very high porosity (>50%) with voids up to 1 cm wide and showing no preferential orientation. At a microscopic scale, the matrix is mainly composed of opaline silica in which remnants of silica-coated microbial filaments can be observed, as well as clotted textures (Fig. 3b).
Sample ‘Biffy’ is a white, dark brown, red, and yellow sample (Fig. 2c), with irregular to coniform siliceous microstromatolites with wavelengths and amplitudes ranging between 0.2–0.5 and 1–2 cm, respectively (Figs. 2c and 3c). In thin section, the microstromatolites were observed to be well laminated, defined by alternating silica and iron-rich laminae (Fig. 3c). Interbedded with the microstromatolites are beds of fine-grained clastic sediment and crustiform layers of ferruginous material. This ferruginous material is intergrown with the siliceous stromatolites, particularly along stromatolite margins (Fig. 3d). Typically, ferruginous crusts have sharp basal contacts and stubby branching internal structures of possible microbial origin (Fig. 3d) (Böhm and Brachert, 1993; Bosak et al., 2013).
At a macroscopic scale, the hand samples SC Low and SC Mid from the SC fossilized hot spring display similar textural characteristics (Fig. 2d, e). The top part of the hand sample of SC Mid (Fig. 2e) is a single 2.5 cm thick layer containing large lenticular voids up to 1 cm wide. Both samples comprise two well-defined sinter layers. At the base, the 3.5 cm thick layer is highly porous and comprises five stacked terracettes of coarsely silicified and subvertically packed micropillar structures.
The contact horizon between the terracettes is darker in color than the light-colored terracettes. The dense and subvertical arrangement of micropillar structures in the terracettes is typical of the palisade microfacies, where micropillars represent silica-encrusted cyanobacterial sheaths (Walter et al., 1996; Campbell et al., 2001, 2015). The palisade microfacies have been reported in various siliceous hot springs from Holocene to Paleozoic age (Walter et al., 1996, 1998; Campbell et al., 2001, 2015).
At a macroscopic scale, the palisade textures are not as clearly identified in sample SC High (Fig. 2f) as in samples SC Low and SC Mid. In thin section, however, the palisade textures are well defined, as are small, fossilized filament splays that rise up from a single point (Fig. 3e).
SEM analysis of SC Mid revealed palisade textures within fossilized microbial mats, preserving bundles of filaments forming the framework around void spaces (Fig. 4a). These beds of “botryoidal grains and chains” are termed particulate sinters (Wilmeth et al., 2020) and represent colloidal silica precipitating on the outside of the filament bundles.

Scanning electron microscopy images of selected Yellowstone samples.
Samples AL Base and AL Fil from the MGB are two parts of one sample (Fig. 2g), separated because the lower white, ∼2.5 cm of the sample (AL Base), is more silicified than the upper, gray-green part of the sample (Al Fil) that consists of densely packed, friable unfilled filament casts. Thin-section petrography of sample AL Fil showed that it consists of widely spaced, large (750–1500 μm), crudely upward-branching knobby, opaque filaments within clear opaline silica (outlined in Fig. 3f). These filaments are highly irregular and much larger than characteristic microbial filaments.
Sample AL Strom is a dense, finely laminated siliceous sinter (Fig. 2h). The 0.5 mm thick laminations can be observed within the entire sample and are wave-shaped. Perpendicular to the bedding, individual subvertical elongated voids up to several centimeters high are also observed. In SEM, this sample consists of a flowing mesh of silicified, fine filamentous microbes that surround ovoid to elongate fenestrae (Fig. 4b). Perpendicular cone shapes have been previously documented in conical tufted sinter facies from modern to Jurassic sinters (Guido and Campbell, 2011; Lynne, 2012). Conical tufts represent irregularly laminated, cone-shaped, finely filamentous cyanobacteria (Walter et al., 1996; Lynne, 2012).
Sample AL Cement is a featureless sample of siliceous cement in hand sample (Fig. 2i), but SEM analysis revealed a meshwork of silicified, relatively short, filamentous microbes around poorly defined fenestrae (Fig. 4c). The filaments are randomly oriented and do not exhibit the same columnar morphologies as the SC samples. The silica filament casings are infilled with silica and, thus, are not open porous space.
Raman spectroscopy analyses were acquired on multiple (≥5) separate areas in the samples HB C, ‘Biffy’, SC High, AL Fil, AL Base, and AL Cement. All the samples studied have a broad band in the ∼200–500 cm−1 range (Supplementary Fig. S4), which indicates the presence of opal-A silica (Curtis et al., 2019). Each sample was carefully screened for Raman signals of OM, but only ‘Biffy’ and AL Cement provided clearly resolvable bands of OM. The OM in both samples shows near-identical spectral features in the ∼1050–1700 cm−1 region with distinctive maxima at ∼1365–1370 and 1590 cm−1, which can be assigned to the disorder induced (D) and graphitic (G) bands of OM, respectively (Fig. 5a).
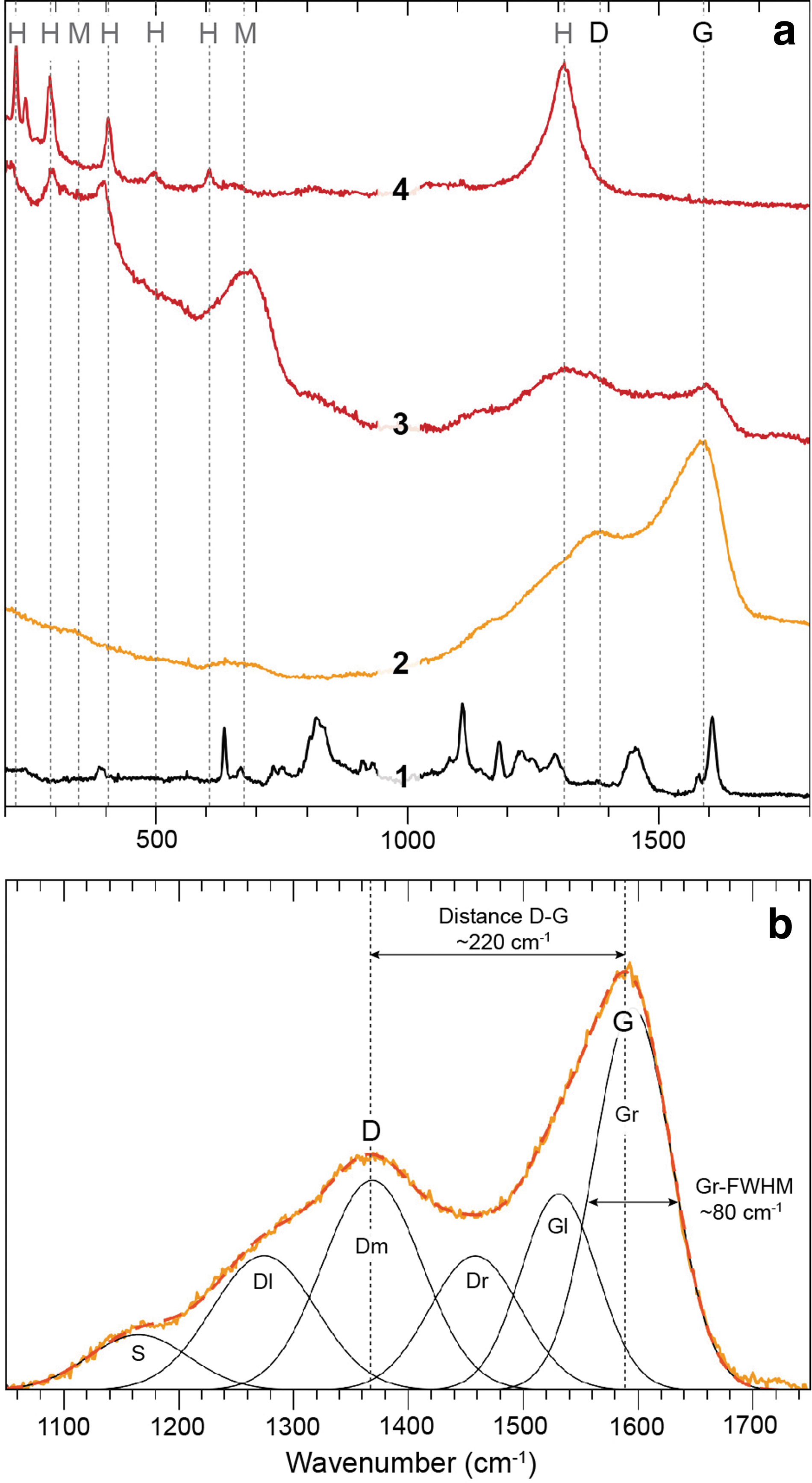
Raman spectroscopy analyses of the ‘Biffy’ Yellowstone sample.
Raman spectroscopy also indicates that the OM in ‘Biffy’ is commonly overgrown or replaced by Fe-oxides, presumably hematite and maghemite, as is indicated by a series of diagnostic peaks in the low-wavenumber (200–700 cm−1) and high-wavenumber (1310 cm−1) ranges (Fig. 5a) (Bellot-Gurlet et al., 2009). Spectral deconvolution in the 1050–1750 cm−1 range following Schito et al. (2017) resolves spectral features typical of immature kerogens, as is indicated by the low spectral distance between the D and G band (‘Biffy’ = ∼220 cm−1 and AL Cement = ∼226 cm−1), and the large full width at half maximum (FWHM) of the ordered right (Gr) band at ∼1590 cm−1 (both samples ∼80 cm−1), both features that indicate a thermal maturity below the oil window (Fig. 5b) (Schito et al., 2017).
All samples have total carbon that is below the detection limit (<50 ppm).
Only aliphatic hydrocarbons were detected in the total hydrocarbon fraction of the extractable OM of the 10 samples, with no detectable peak in the aromatic hydrocarbon fractions (e.g., Supplementary Fig. S5). All integrations were performed on SIM data, and the descriptions of compound distributions are of SIM data, unless otherwise stated (Section 3.3.4).
n-Alkane and acyclic isoprenoid distributions
A unimodal series of n-alkanes was detected in all samples (Fig. 6). The distributions are relatively similar (Fig. 6, Supplementary Fig. S5, and Table 1), but there are subtle differences as described below. All the Yellowstone samples studied here, except for the ‘Has Been Hot Spring’ samples, contain a strong enrichment of n-C17 relative to n-C18 [e.g., Figs. 6 and 7; n-C17/((n-C16 + n-C18)/2) values >1 in Table 1]
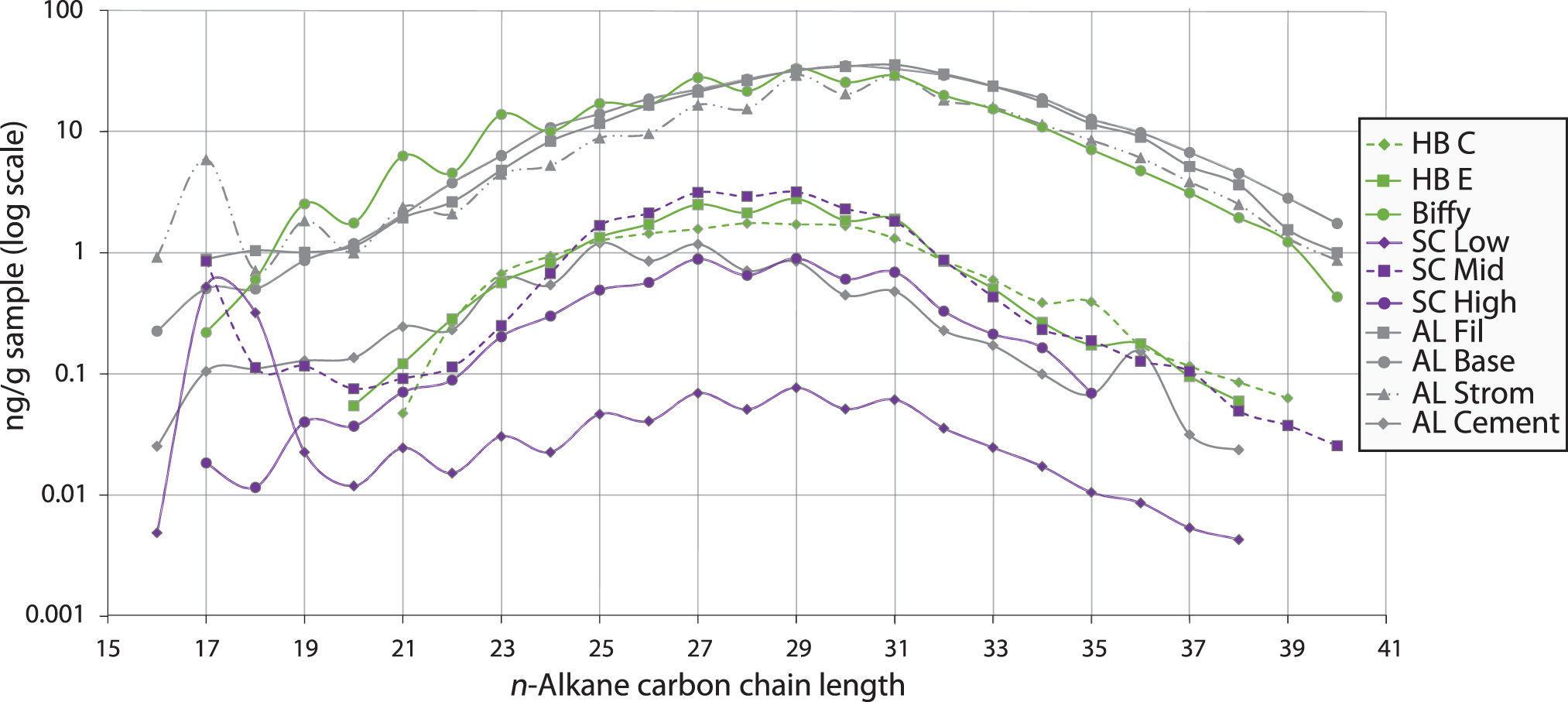
Amount (ng/g sample) and distribution of n-alkanes in the Yellowstone samples in this study.
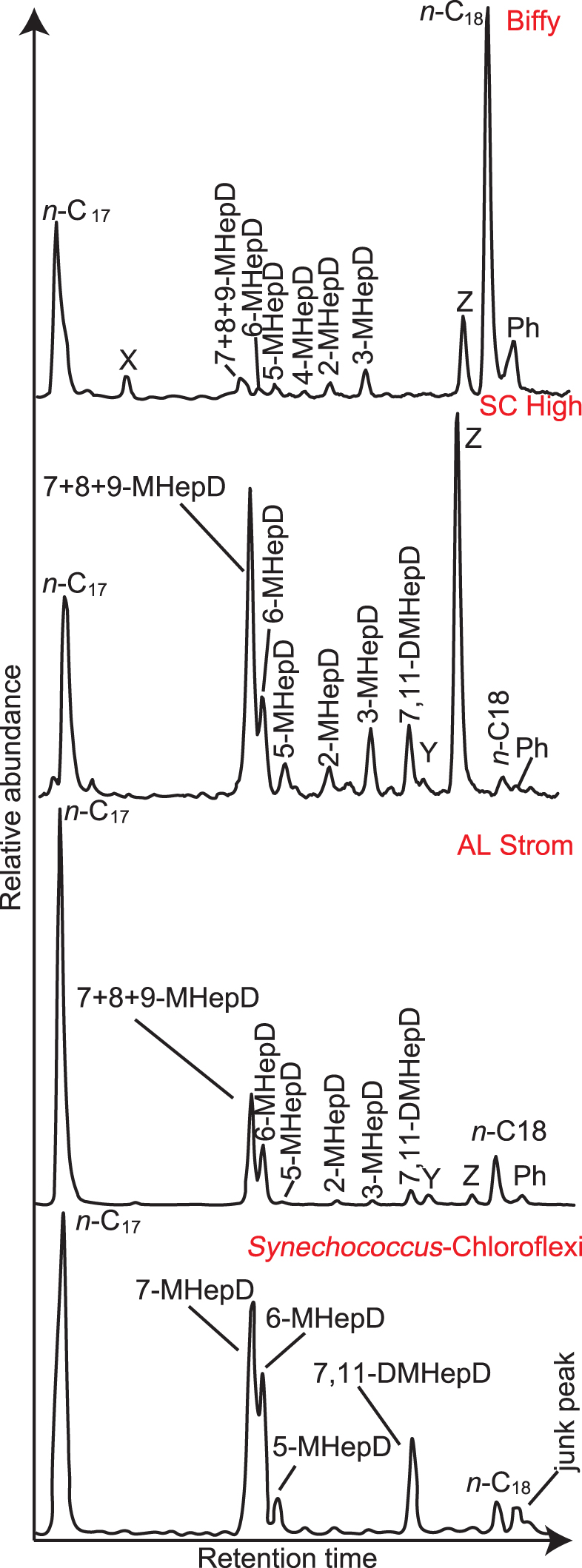
Partial m/z 57 mass chromatograms showing the distributions of the n-C17 and n-C18 n-alkanes, MHepD isomers, and a DMHepD isomer (7,11-DMHepD) in the ‘Biffy’, SC High, and AL Strom samples from Yellowstone. The peak labeled 7 + 8 + 9-MHeD is due to co-eluting 7-MHepD, 8-MHepD, and 9-MHepD (the mid-chain isomers). The peaks labeled “X,” “Y,” or “Z,” refer to unidentified isomers, which were detected in single-ion monitoring mode. For comparison, a similar partial mass chromatogram is provided for a Synechococcus-Chloroflexi mat collected from Chocolate Pots in Yellowstone, as previously published by Parenteau et al. (2014). The mass of the chromatogram was not provided by Parenteau et al. (2014); the peak labeling is from that article. DMHepD, dimethylheptadecane; MHepD, methylheptadecanes; m/z, mass-to-charge ratio.
The two HB samples have similar, convex n-alkane distributions that range from about n-C20 to n-C38, and both have similar quantities of n-alkanes (∼2 ng/g of crushed sinter at the maxima of n-C28–n-C29). Sample HB E has a slight odd-over-even carbon number predominance (OEP) for the n-alkanes, indicated by a carbon preference index (CPI22–32) of 1.28, whereas sample HB C has virtually no OEP (CPI22–32 = 1.03). The ‘Biffy’ sample also has a convex n-alkane pattern, but over a much wider distribution, with n-alkanes ranging from n-C17 to n-C40, with a maximum at n-C29 (Fig. 6). ‘Biffy’ is enriched in n-alkanes by more than a factor of 10 compared to the two HB samples, with ∼35 ng/g for n-C29. This sample has a stronger OEP for the n-alkanes than the HB samples, both at high molecular weight (CPI22–32 = 1.41) and at medium molecular weight (CPI16–26 = 1.58; Table 1 and Fig. 6).
The three SC samples have similar, broadly convex, n-alkane distributions (Fig. 6). SC Low contains n-alkanes from n-C16 to n-C38, with a maximum at n-C17, whereas n-alkanes in SC Mid and SC High range from n-C17 to n-C35/n-C40 and have maxima at n-C28 and n-C29, respectively. These three samples have a strong OEP for medium molecular weight n-alkanes (CPI16–26 = 1.07–1.65), and CPI22–32 ratios >1 (Table 1).
The AL samples also have generally similar n-alkane distributions to each other, extending from ∼n-C16 to >n-C38/40 (Fig. 6). AL Strom, AL Base, and AL Fil reach maxima of ∼30 ng/g at n-C30 or n-C31, whereas AL Cement reaches a maximum of ∼1.2 ng/g at C25. Both CPIs of AL Base and AL Fil are ∼1, whereas AL Strom and AL Cement have a strong OEP, with CPIs of 1.5–1.6 (Table 1).
Pristane (Pr; 2,6,10,14-tetramethylpentadecane) and phytane (Ph; 2,6,10,14-tetramethylhexadecane) were detected in some samples (Table 1). ‘Biffy’, AL Fil, and AL Cement have Pr/Ph values of 0.36–0.55 (Table 1). ‘Biffy’, AL Fil, and AL Cement have Pr/n-C17 values of 0.25–0.45. Ph/n-C18 ratios of 0.22–0.67 were measured in ‘Biffy’ and the four AL samples.
The samples contain medium to high molecular weight monomethylalkanes (MMAs) (Supplementary Fig. S6). All the SC samples contain C18 and C26–C36 MMAs. Sample HB C contains C23–C36 MMAs, sample HB E contains C25–C36 MMAs, and ‘Biffy’ contains C18–C40 MMAs. The AL samples have similar distributions to ‘Biffy’, with AL Strom, AL Fil, and AL Base containing C17–C40 MMAs, whereas AL Cement contains C18–C36 MMAs. While the mid-chain methylheptadecanes (MHepD) are more dominant than 2-MHepD and 3-MHepD (Fig. 7), in the longer chain MMAs, it is the 2- and 3-substituted isomers that are typically the most dominant (Supplementary Fig. S6). The exceptions are the mid-chain methylpentatriacontanes (C36) in several samples (e.g., ‘Biffy’ in Supplementary Fig. S6), which are more dominant than the 2- and 3-substituted isomers.
For all samples that contain MHepDs, the mid-chain MMAs (especially co-eluting 7 + 8 + 9-MHepD) are much more abundant than 2-MHepD and 3-MHepD. The distribution of MHepDs is varied in all samples, even between samples from the same site (Fig. 8). ‘Biffy’ is the only sample from the NGB that was found to contain MHepDs, which are dominated by 3-MHepD and the three co-eluting mid-chain isomers. Some samples contain a peak eluting after 3-MHepD based on relative retention times (Kenig et al., 1995; Parenteau et al., 2014) and mass spectral characteristics from SIM data are identified as 7,11-dimethylheptadecane (DMHepD) (Figs. 7 and 8).

Pie charts, showing the distribution of methylheptadecanes (MHepDs) and a dimethyl heptadecane isomer (7,11-DMHepD) in the samples from this study. X, Y, and Z refer to unknown peaks eluting in the same general molecular weight range (n-C17–n-C18; see Fig. 7).
‘Biffy’ also contains large amounts of two unidentified isomers that we refer to as peaks X and Z, which are present in several of the samples. Peak X elutes after n-C17, but before the MHeDs, whereas peak Z elutes directly before n-C18. We infer, based on SIM data and retention time behavior, compared to the work of Kenig et al. (1995), that peak Z is an unidentified DMHepD (Fig. 7).
In sample SC Low, peak Z is the most abundant peak of the branched alkanes in the n-C17–n-C18 range, and 6-MHepD is the most abundant MHepD (Fig. 8). In contrast, sample SC Mid contains co-eluting 7 + 8 + 9-MHepD as the most abundant peak. In sample SC High, peak Z and co-eluting 7 + 8 + 9-MHepD are almost equally most abundant, with lesser amounts of 6-MHepD and 5-MHepD. A third unidentified isomer (peak Y) is present in sample SC High in approximately the same proportion as 5-MHepD and 6-MHepD (Figs. 7 and 8).
The AL samples are likewise diverse. Samples AL Base and AL Fil are similar to one another, with the two most abundant peaks being peak X and co-eluting 7 + 8 + 9-MHepD. Sample AL Strom is dominated by co-eluting 7 + 8 + 9-MHepD (∼50%) and 6-MHepD (Figs. 7 and 8). AL Cement has no single most abundant peak, but rather peak X, 3-MHepD, co-eluting 7 + 8 + 9-MHepD, and 5-MHepD, each represent about 1/5 of the total composition.
A series of hopanes were detected in HB E and ‘Biffy’ from the NGB and in the AL samples but were not detected in the SC samples or the blank (Supplementary Fig. S7 and Table 1). The hopanes detected include C27 18α-trisnorneohopane (Ts), C27 17α-trisnorhopane (Tm), C29 and C30 αβ and βα hopanes, C31–C35 αβ homohopanes, and C29Ts. C30 αβ hopane is the most abundant hopane in all the samples, and C29/C30 αβ hopane ratios vary from 0.71 to 1.0 (Table 1).
The C31 and C32 aβ 22S/(22S+22R) homohopane isomerization ratios of ‘Biffy’ and AL samples range from 0.55 to 0.63, and 0.59 to 0.67, respectively. The C32 aβ 22S/(22S+22R) ratio could not be determined for AL Fil and AL Strom due to an unknown peak co-eluting with the S isomer. The C29 and C30 αβ/(αβ+βα) hopane ratios range from 0.60 to 0.73 and 0.56 to 0.79, respectively (Table 1). The samples have moderate Ts/(Ts + Tm) ratios (0.44–0.69).
C20–26 tricyclic terpanes and C24 tetracyclic terpane are present in the samples in much lower abundance than the hopanes (Supplementary Fig. S7). No methylhopane, sterane, or diasterane was detected in any sample, despite careful checking of the appropriate SIM mass chromatograms.
Common contaminants
We searched in full scan mode using m/z 127 for branched alkanes with quaternary carbon (BAQCs), which are common contaminants in organic geochemistry, and confirmed that these are not major constituents of any of the samples analyzed (e.g., Supplementary Fig. S8). While there are some phthalates and siloxanes present in all samples (m/z 73 and m/z 149) (Supplementary Fig. S9), these compounds are minor components, except in the SC High sample. The phthalates and siloxanes do not interfere with any of the compounds that were integrated in most samples. In SC High, the siloxane component is more than 10 × as large as the n-alkanes and elute at a similar time as the 3-methylalkanes. Due to this interference, we make no interpretation about the 3-methylalkanes in SC High.
Discussion
Syngeneity and thermal maturity of OM
Before evaluating the source of the OM in these samples, it is important to consider whether the OM is syngenetic, as OM is known to be transported and migrated throughout sinter aprons (Gonsior et al., 2018; Reinhardt et al., 2019; Teece et al., 2020, 2022).
Isoprenoid/n-alkane ratios can be used to determine thermal maturity, because the cracking of kerogen generates more n-alkanes, so that the ratio of acyclic isoprenoids relative to n-alkanes decreases (Tissot et al., 1971; Blumenberg et al., 2012; Mißbach et al., 2016). The low Pr/n-C17 (<0.5) and Ph/n-C18 (<0.7) ratios detected in the Yellowstone samples ‘Biffy’, AL Fil, and AL Cement are typical of samples in the early oil window (Aharonovich and George, 2016).
The Pr/n-C17 can also be source related if the thermal maturity is low (Lijmbach, 1975; George et al., 2001). For example, if there is a strong input of terrigenous OM. Conversely, the high n-alkane OEPs in many of the samples are consistent with low thermal maturity, as well as terrigenous plant input based on CPI22–32 > 1 (Table 1). Some samples have CPIs close to 1.0, which could indicate a more thermally mature input (Peters et al., 2005).
The 22S/(22S+22R) homohopane isomerization ratios increase with increasing thermal maturity and tend to reach equilibrium at ∼0.57–0.62 in the early oil window (Seifert and Moldowan, 1980; Peters et al., 2005). The C31 and C32 aβ 22S/(22S+22R) homohopane ratios of the ‘Biffy’ and AL samples (0.55–0.67) suggest that they have been thermally matured at least into the early oil window. In support of this conclusion, no hopenes or ββ hopanes were detected in the samples (Supplementary Fig. S7). The C29 and C30 αβ/(αβ + βα) hopane ratios can also be used as thermal maturity parameters, since the βα hopanes are less thermally stable than the αβ hopanes, so these ratios increase with increasing thermal maturity (Peters and Moldowan, 1991; Zhang and Li, 2018), from ∼0.2 for immature OM to about 0.95 in the peak oil window.
The ‘Biffy’ and AL samples have αβ/(αβ + βα) hopane ratios of 0.56–0.79, which suggest that the samples have been matured to within the main part of the oil window. Because Ts is more stable than Tm during thermal maturation, the Ts/(Ts+Tm) ratio increases with thermal maturity, although a source influence can potentially affect this ratio (Seifert and Moldowan, 1980; Zhang and Li, 2018). Nevertheless, all the analyzed samples come from the same facies, meaning that any source interference is likely negligible, and the Ts/(Ts+Tm) ratios of 0.44–0.69 for the Yellowstone samples are also consistent with thermal maturities in the oil window.
The thermally mature hopane biomarkers detected in ‘Biffy’, AL Base, AL Fil, and AL Strom (Table 1), together with the isoprenoid/n-alkane ratios in the same samples, are inconsistent with the lack of aromatic compounds detected in any of the samples in this study and the thermally immature n-alkane signal (CPI22–32 > 1.4) in the ‘Biffy’ and AL Strom samples. Raman spectroscopic data from ‘Biffy’ and AL Cement show a distance between the D and G bands of ∼220 cm−1, which correlates with a vitrinite reflectance equivalent (VRE) of ∼0.45%, and a full width at half maximum of the Gr band (Gr-FWHM) of ∼80 cm−1 (VRE = 0.55%) (Schito et al., 2017).
These data are consistent with the OM in these samples, having nearly reached the oil window, despite the samples never having been buried. Moreover, the thermally mature signals for samples AL Base and AL Fil (hopanes, low isoprenoid/alkane ratios, and CPIs of ∼1) are interpreted to come from migrated OM, as no kerogen signal could be detected in these samples by Raman spectroscopy or total carbon measurement. These two samples are friable and porous, which means they would have been suitable conduits for migration of hydrothermal fluids.
Origins of OM
The low levels of carbon detected in this study are consistent with the interpretations of Havig et al. (2021), that most of the hot spring biomuculus is made up of silica, with carbon only comprising a small fraction. All samples from this study contain evidence for microbial organisms, as deduced from organic geochemistry, as well as diagnostic macrotextures and microtextures observed in hand samples and thin sections. The hydrocarbon composition of samples contains evidence of bacteria, along with other inputs, such as from land plants, as reflected in the n-alkane distributions (Fig. 6). Previously reported major hydrocarbon components of hot spring sinters are typically n-C17 and mid-chain MHepDs (e.g., Shiea et al., 1990; Teece et al., 2020).
Specifically, 7-MHepD and 8-MHepD are considered to be biomarkers for cyanobacteria (Shiea et al., 1990; Jahnke et al., 2004). In modern cyanobacteria, 6-MHepD is not considered to be a major product, but some cyanobacterial species common in hot springs (such as Fischerella) produce more 5- and 6-MHepDs than 7- and 8-MHepDs (Coates et al., 2014). Previously, Teece et al. (2020) and Parenteau et al. (2014) reported these compounds within hot spring sinters in the El Tatio hot spring field, and hot spring pools within the Taupo Volcanic Zone and Yellowstone National Park. Other MMAs can be considered to be biomarkers of bacteria more generally, as are hopanes (Ourisson et al., 1979; Brocks and Pearson, 2005).
The wide variety of distributions of MMAs in the samples of different hot springs across the Yellowstone Basin (Fig. 8) is consistent with the diversity of microbial communities observed in different hot springs in Yellowstone (e.g., Reyes et al., 2013; Schuler et al., 2017; Havig and Hamilton, 2019) and in hot springs more generally (e.g., Cady and Farmer, 1996; Farías et al., 2013; Toneatti et al., 2017; Sriaporn et al., 2020).
Parenteau et al. (2014) analyzed the total organic profile of several different types of mats found within Chocolate Pots in Yellowstone. As this study considers OM that has preservation potential in deep time (i.e., hydrocarbon skeletons), we compared the hydrocarbon distributions of our Yellowstone MHepD data with GC-MS data from a phototrophic Synechococcus-Chloroflexi mat from Chocolate Pots in Yellowstone (Fig. 7) (Parenteau et al., 2014), as we would if we were considering more ancient or thermally matured OM (e.g., Hoshino and George, 2015). Our data show similarity between the extant mat and the hydrocarbon composition in the samples SC High and AL Strom.
16S ribosomal RNA (rRNA) gene sequences and metagenomic data recovered from Chocolate Pots, as well as other similar locations, show that Chloroflexi and Synechococcus biomasses can be major constituents of phototrophic mats in at least some Yellowstone hot springs at the time of sinter formation (Bennett et al., 2020). The possibility of matching the distribution of MHepDs in modern samples to DNA sequencing studies may help determine which bacteria were present in a given geological sample.
Genes, chromosomes, and especially DNA are usually not well preserved in geological samples (Armbrecht et al., 2019), with some exceptional cases of preserved ancient DNA in ∼400,000-year-old cave sediments (Willerslev et al., 2003) or permafrost settings up to 1 million years old (Smith et al., 2003; Orlando et al., 2013). However, branched alkanes, such as MMAs, are present in rocks throughout much of the geological record and have even been reported in the early Archean (Marshall et al., 2007; Duda et al., 2018).
Long-chain MMAs (>C18) are present in all the samples (Supplementary Fig. S6), but the source of these compounds is more unclear as they can have multiple sources. Examples include insect waxes (Kenig et al., 1995; Zhang et al., 2011) and sulfate-reducing bacteria that produce both 2- and 3-methyl fatty acids, which can be transformed into 2- and 3-methylalkanes (Peckmann et al., 2004). Furthermore, algae and higher plants are also sources of long-chain MMAs (He et al., 2009; Ji et al., 2009). The 2- and 3-methylalkanes are dominant at most chain lengths >C18 reported in this study (Supplementary Fig. S6).
Inorganic processes such as thermal maturation and catalysis can also produce long-chain MMAs (Kissin, 1987; Peters et al., 2005). So, while the MHepDs in the samples are clearly linked to biological sources, the source(s) of the longer chain MMAs are more ambiguous and cannot be clearly linked to a single source, as all the sources mentioned are possible. However, given the strong higher plant signal in the n-alkane series in most of these samples, it seems likely that higher plants played a role in the presence of these compounds.
Other OM components of the Yellowstone samples include n-alkanes with an OEP for both the medium-chain (C16–26) and long-chain inputs (C22–32), indicating both an algal and a terrigenous (plant) input (Eglinton and Hamilton, 1967; Maffei et al., 2004), except for AL Base and AL Fil, which exhibit no chain length preference.
Pr and Ph are most commonly derived from the phytol side chain of chlorophyll (Powell and McKirdy, 1973; Rontani and Bonin, 2011), and the ratio of Pr/Ph is correlated to redox conditions, with values <0.8 typical of anoxic conditions (Powell and McKirdy, 1973; Holba et al., 2003; Evenick, 2016), but this ratio can be affected by high thermal maturity. The Pr/Ph ratios obtained in this study (0.36–0.55) are consistent with a hypersaline, evaporative environment (Peters et al., 2005), and are similar to those obtained for hot springs in El Tatio and the Taupo Volcanic Zone (Teece et al., 2020).
To summarize the interplay between bacteria and OM, the two HB samples contain no evidence of bacteria, and the bacterial nature of these samples is only interpreted through petrography. The ‘Biffy’ and AL samples contain hopanes, terpanes, and/or MHepD, which are evidence of bacterial input. Finally, the SC samples contain MHepDs and n-C17/((n-C16 + n-C18)/2) values >1, also consistent with bacterial input.
Preservation of OM
Previous work on hydrocarbons and the detection of life in fossilized hot springs (Preston et al., 2008; Teece et al., 2020) suggest that the crystallinity of silica may be a primary factor in the preservation of OM. Such a relationship between substrate crystallinity and OM preservation can be explored further based on the Yellowstone samples in this study. Raman spectroscopy and previous work by Gangidine et al. (2020) indicate that the samples studied here are dominantly composed of opal-A (Supplementary Fig. S4). Furthermore, the SC samples also exhibit botryoidal silica growth in threads in open pore space (Fig. 4a), which can be attributed to opal-A precipitation on microbial filaments (Preston et al., 2008).
However, although n-alkanes could be detected by SIM in the SC samples, they are of low abundance, the least of all samples studied here (Fig. 6). These observations are inconsistent with the proposition that better OM preservation occurs in samples dominated by opal-A. Furthermore, the previously reported microcrystalline quartz sample from Te Kopia in the Taupo Volcanic Zone (Teece et al., 2020) has a similar abundance of n-alkanes as the SC samples, which indicates that crystallinity may not be the sole factor that controls OM preservation.
Teece et al. (2020) concluded that crystallinity was a more important control on preservation than age, based on the abundance of hydrocarbons in the Taupo Volcanic Zone samples with ages ranging from modern to ∼9.4 ka. However, as biomass varies both within and between hot springs (Waples, 1980; Havig et al., 2011; Hamilton et al., 2019; Havig and Hamilton, 2019), this result may simply be an artifact of the original biomass variability during deposition and formation of the sinters.
Other studies have reported that the prevalence of lipids in hot spring deposits is linked with textural evidence for micro-organisms (Preston et al., 2008). However, The SC samples studied here are composed of opal-A and have a subtle green tinge from phototrophic communities and open pore spaces when observed in SEM (Fig. 4a), yet they contain no hopane biomarkers and have relatively fewer n-alkanes than the other, more lithified, samples. All the SC and AL samples show macroscopic and microscopic mat textures with abundant microbial filaments, whereas the HB samples do not show textural evidence for the presence of microbial mats. While the AL Base and AL Fil samples contain both mat textures and hopane biomarkers, the open pore space within the samples may have allowed for the outward migration of fluids containing soluble OM. Thus, the correlation between textural features and soluble OM can be discounted here.
We conclude on the basis of these observations that the OM of these samples most likely reflects in situ overprinting of immature OM by migrating fluids, rather than a purely syngenetic depositional signal of immature OM. Therefore, unambiguous associations between textures and lipids cannot be determined for this Yellowstone sample set. Thus, drawing conclusions about the effect of crystallinity on preservation in hot spring sinters is limited without prior knowledge of biomass, and intersite and interthermal spring comparisons.
Mechanism for maturation of OM
The mixture of immature and mature signals in the Yellowstone hot spring sinter deposits documented in this study has previously been documented within active hot springs in Yellowstone National Park (Gonsior et al., 2018), in fossilized hot springs in the Taupo Volcanic Zone in New Zealand (Teece et al., 2020), and in Pleistocene hydrothermal cherts (Reinhardt et al., 2019). We document it here for the first time in fossilized hot spring deposits in Yellowstone and try to link these observations to contextual details. One possible explanation for the heterogeneous maturation of OM is that the OM gathered in surface runoff may have been heated in the near-boiling conditions of an active hot spring due to fluid recirculation. However, it is unclear whether this process would sufficiently mature the OM in surface hot spring pools where the sinter was not consistently submerged. Furthermore, this process would not explain why some of the kerogen is in the sub-oil window or how differential maturation of OM might occur within a single sample.
Alternatively, both Gonsior et al. (2018) and Teece et al. (2020) proposed that OM could be matured in deeper, higher temperature parts of the hot spring system underground and then recirculated and incorporated into indigenous OM within surface deposits, a phenomenon termed “the hydrothermal pump hypothesis” (Duda et al., 2018). For example, Reinhardt et al. (2019) suggested that OM that has both a different origin and thermal maturity can be rapidly incorporated into kerogen during deposition, preserving heterogeneous OM that is then distributed in the environments.
Gonsior et al. (2018) applied this model to active hot springs in Yellowstone, but it may also apply more widely, as there are several reports of OM in hot springs in places where this material would not be expected to occur. For example, there is a report of sterols (a biomarker for eukaryotes) at the opening of a fumarole in Ethiopia (Carrizo et al., 2019). Fumarolic temperatures can reach 161°C, while the maximum survival limits of most eukaryotes are ∼60°C (Tansey and Brock, 1972; Angilletta, 2009). Although Carrizo et al. (2019) attributed the transport of OM to above-ground processes, such as wind transport or surficial water transport, alternative subsurface mechanisms could be considered.
Clifton et al. (1990) described the presence of hydrothermal oils restricted to the northeastern part of Yellowstone National Park. These oils were generated in the subsurface and transported by hydrothermal fluids but are different in composition to the compounds detected in the samples described in this study. The OM in the Calcite Springs oils was largely aromatic and contained <1% saturated compounds. These Calcite Springs oils had similar thermal maturity to the compounds detected in this study, with C32 αβ 22S/(22S+22R) hopane ratios of ∼0.59, although the Ts/(Ts+Tm) ratios are much lower (∼0.3), and these oils contain immature ββ hopanes. The Rainbow Springs oil was largely composed of n-alkanes, like the samples analyzed in this study, although it had a Pr/Ph ratio >5, indicating that it came from a terrigenous source, which is not consistent with the Pr/Ph ratios described in this study.
Further evidence that the compounds in the samples described in this study are not derived from the same source as the hydrothermal oils (Clifton et al., 1990) is that HB E, HB C, and ‘Biffy’ are the samples that come from the NGB, closest to Calcite and Rainbow Springs. Samples HB E and HB C do not contain thermally mature cyclic biomarkers, nor similar n-alkane distributions or Pr/Ph ratios as the hydrothermal oils.
Terrestrial hot springs are transient, and the temperature can vary widely from the proximal apron to the distal apron (e.g., Campbell et al., 2015). Moreover, temperatures are not necessarily consistently hot or cool in specific parts of an ever-changing and evolving hot spring system, so that signatures within a single sinter sample might represent OM from both thermally heated events and cooler events. For the Yellowstone samples in this study, the transport and migration of OM require a mechanism for maturation that seems inconsistent with above-ground processes, as Raman results from ‘Biffy’ and AL Cement indicate OM that is nearly in the oil window, despite lack of burial. Furthermore, the lack of abundant kerogen in AL Base and AL Fil, based on the Raman data, indicates that the extractable OM of these samples was likely transported.
The temperatures to which the OM was exposed before migration within a hot fluid to the surface cannot be readily determined, as thermal maturation depends on two factors: time and temperature. To reach temperatures within the main part of the oil window, as indicated by hopane ratios for some of the Yellowstone samples, requires temperatures of about 120°C, based on slow burial over geological time in a sedimentary basin (e.g., Waples, 1980; Killops and Killops, 2005). However, for shorter timeframes, higher temperatures are required to mature the hydrocarbons to the same amount (Lewan and Ruble, 2002). It is likely that these higher temperatures would be reached below ground in a hot spring environment, such as Yellowstone, given that the heat source of the hydrothermal system is from a crustal magma chamber.
We therefore infer that OM maturation occurred as a result of below-surface processes in the hotter, deeper parts of the hydrothermal system, and that this exogenous material was then transported and emplaced within the sinter and its microbial community during pulsed outflow events of hot, deep fluids. This process led to overprinting of thermally matured OM with syngenetic, immature OM that formed in the hot spring surface environment, resulting in the preservation of two, mixed OM components (exogenous and indigenous) in the sinters. Thus, based on these results, the hydrothermal pump hypothesis suggested by Duda et al. (2018) and mechanisms recognized by Reinhardt et al. (2019) are favored here to explain the complex pattern of OM distribution and thermal maturity at Yellowstone.
Conclusions
In this study, we examined how contextual studies can help to determine whether the OM found within siliceous sinters is exogenic—introduced through transport and circulation—or developed in situ by microbial communities, or a mixture of both. Morphological, mineralogical, and geochemical data are presented for fossilized sinters from three distinct areas of Yellowstone National Park. Organic geochemistry of the extractable OM reveals the presence of n-alkanes, methylalkanes, hopanes, and other terpanes, but no aromatic compounds.
The extractable OM preserved within these samples contains a mixture of immature and mature hydrocarbons, while analysis of the kerogen by Raman spectroscopy reveals that OM nearly reached the oil window without burial. Based on previous reports of below-ground hydrothermal transport of OM in Yellowstone and considering the general mechanisms for thermal maturation of OM, the hydrothermal pump hypothesis suitably explains the molecular heterogeneity of OM in the studied samples. However, above-ground processes for the maturation of the OM cannot be completely discounted.
Previously reported GC-MS data of an active Synechococcus-Chloroflexi mat were compared with new GC-MS biomarker data on methylheptadecanes from the fossilized hot spring sinters. The distribution of methylheptadecanes is consistent with observations of Synechococcus and Chloroflexi as abundant members of extant mats in alkaline hot springs. This approach is a unique opportunity to infer microbial community composition in the geological past, from samples in which body fossils and genomics may not be preserved well.
This work furthers the investigation of silica phases and their control on the preservation of OM in hot springs. The dynamic environments in hot springs require contextual and multidisciplinary knowledge and considerations. A study that examines OM in Yellowstone holistically, that is, the polar, apolar, and insoluble geochemistry in combination with metagenomic analyses will be an important next step, but was out of scope for this research, which examined the apolar OM compounds, as they are the ones most likely to be preserved through geological time.
Specifically, a systematic investigation of OM preservation, especially in relation to substrate crystallinity and biomass maturation, is key to reliably assessing the interplay between the varied geological and biochemical processes in such dynamic systems. Taken together, the results and interpretations in this study are critical for better analysis of the evolution of extant and ancient hot spring environments on Earth and elsewhere in the solar system, especially Mars.
Footnotes
Acknowledgments
We would like to acknowledge and pay our respects to the Bedegal people (UNSW), the Wattamattagal Clan of the Darug Nation (Macquarie University), and the 26 tribes of First Nations people with historic connections to Yellowstone National Park, especially the Tukudeka (Shosone-Bannock), Apsáalooke (Crow), and Nimiipuu (Nez Perce). We would also like to acknowledge helpful and constructive comments from reviewers of this article, and reviewers of B.L.T.'s PhD thesis. Yellowstone National Park samples were collected and analyzed in accordance with Yellowstone National Park Research Permit number YELL-2017-SCI-7020 held by J.R.H. and T.L.H. Samples may not be obtained/analyzed by any individual outside of this permit unless a material transfer agreement has been established with J.R.H. and T.L.H. and the Yellowstone Permit Office. Tara Djokic and Andrew Gangidine provided assistance in the field. We thank Julien Bourdet (CSIRO Energy) for access to the Raman Spectroscopy laboratory and help with data acquisition.
Authors' Contributions
B.L.T. assisted in project design, ran the experiments, collected data from GC-MS and Raman spectroscopy, interpreted data, assembled figures, wrote the initial draft of the article, and managed and edited the subsequent versions. J.R.H. assisted in project design, carried out the field work and SEM analyses, interpreted geochemical data and contextual implications, assembled figures, and wrote parts of the article. S.C.G. provided access to the organic geochemistry laboratory, assisted in interpretation of data, and wrote and edited parts of the article. T.L.H. assisted in project design, carried out the field work, interpreted microbiological implications, and edited the article. R.J.B. collected Raman data and assisted in interpretation of that data, assembled figures, and edited the article. J.H. assisted in interpretation of data and edited the article. M.J.V.K. assisted in project design, contextual interpretations, and writing the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
B.L.T. was supported by an Australian Government Research Training Program Scholarship. T.L.H. was supported by NASA Exobiology grant number 80NSSC20K0614. M.J.V.K. was supported by The Australian Research Council through DP180103204. Organic geochemical analyses were carried out in the Organic Geochemistry laboratory of Macquarie University, which is supported by funding from the Australian Research Council and internal sources, both of which are gratefully acknowledged.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Figure S9
Supplementary Table S1
Associate Editor: Kathleen Campbell