
Abstract
Developing robust microbial survival models for interplanetary and planetary spacecraft requires precise inactivation kinetics for vehicle bioburdens. To generate such data, reliable protocols are required for preparing, testing, and assaying microbial cells or spores on simulated spacecraft materials. New data are presented on the utility of the liquid droplet protocol for applying Bacillus subtilis spores to aluminum coupons. Results indicate that low-density spore monolayers should be created between 2 and 5 × 106 spores per cm2 on individual coupons to prevent the formation of aggregates or multilayers of spores. Such aggregation or multilayers will interfere with the precision of characterizing the effects of UV irradiation on spore survival. Optimum spore monolayers are defined as spore monolayers without overlapping or clustered cells and in which all spores will receive UV photons during assays. The best spore monolayers were created with sterile deionized water (SDIW) on uncoated aluminum coupons, or with SDIW + Triton X-100 (at 0.5 × of the critical micellar concentration) on either uncoated Al-coupons or on Chemfilm Class 1A–coated coupons. The Triton X-100 surfactant improved the uniformity of the monolayers without affecting the sensitivity of the spores to UV irradiation. Furthermore, spore layers created at either 2 × 107 or 2 × 108 spores/cm2 created multi-stacking effects that clearly reduced the precision of the UV irradiation assays. A set of standardized protocols is suggested for spacecraft processing and planetary protection communities to permit directly comparing results from divergent labs.
Introduction
Exposure of bacterial endospores arranged in multilayers or uneven monolayers on spacecraft materials can affect results from biocidal conditions relevant to interplanetary space. Divergent results on spore/cell survival that are driven by protocol issues might alter the scientific consensus on whether bacterial cells or spores can survive in space. For example, dense spore multilayers of several Bacillus spp. (approx. 2 × 108 spores per 8 mm coupons) used in some spaceflight experiments (e.g., Horneck et al., 2012; Panitz et al., 2015) may have inflated the ability of spores to survive the rigors of space due to buried spores being protected from solar vacuum and UV irradiation by overlying spores. In contrast, other studies of monolayers of Bacillus spp. spores exposed to simulated martian surface conditions have indicated that all spores exposed to equatorial solar UVC fluence rates were eradicated over short periods of time (Schuerger et al., 2003, 2006).
Clustering can also result in biphasic survival curves (Cerf, 1977; Schuerger et al., 2006) that may not accurately reflect biocidal effects. Self-shielding occurs in high-density spore layers where one spore does not adhere directly to the test surface but attaches to another spore that is adhered to the test surface. In multilayers, spores may not be exposed to the same dose of any given biocidal factor. For example, in a bacterial cell with a diameter of 1 μm, an estimated 78% of the irradiation at 250 nm (UVC) is transmitted to the cell center, and transmittance decreases at progressively shorter wavelengths (Coohill, 1986).
In direct comparisons, uniform spore layers have been observed with lower survival fractions than uneven spore monolayers (Fayolle et al., 2020). Concerns on the uniformity of liquid-deposited spore monolayers on coupons have also been addressed (Noell et al., 2013; Raguse et al., 2016; Fayolle et al., 2020). Several methods for creating even monolayers have been proposed including aerosol deposition (Edmonds et al., 2009; Levy et al., 2011; Raguse et al., 2016), membrane filtration and membrane transfer to coupons (Noell et al., 2013; Fayolle et al., 2020), direct deposit of sterile deionized water cell suspensions (Mancinelli and Klovstad, 2000; Schuerger, 2022; Schuerger et al., 2003, 2006), different methods for drying spore layers onto coupons (Mancinelli and Klovstad, 2000; Noell et al., 2013), and the use of additives such as surfactants or ethanol (Yunker et al., 2011).
The concentrations of spores on coupons also affect the uniformity of spore layers. Fayolle et al. (2020) reported that spore layers created by liquid deposition of less than 1.0 × 106 per coupon did not result in the same self-shielding as higher concentrations of spores (>2.0 × 106). Studies on the International Space Station have investigated high concentrations of spores (up to 108 spores per coupon) in multilayers—which are not typically found on spacecraft—and the results exhibit long-term survival of bacterial spores under extreme space conditions (e.g., Horneck et al., 2012; Vaishampayan et al., 2012; Panitz et al., 2015). To apply findings to planetary protection, it is important to investigate realistic scenarios of spore concentrations and distributions across surfaces exposed to a diversity of space biocidal conditions. However, not all spore-deposition protocols are easy to utilize.
For example, aerosol deposition is observed to generate uniform monolayers (Levy et al., 2011; Raguse et al., 2016) but requires specialized equipment and increases the biosafety risk as containment of aerosolized bacterial spores is challenging. On the other hand, membrane transfer (Noell et al., 2013; Fayolle et al., 2020) also generates well-distributed monolayers but still requires extra equipment such as filter towers, membrane filters, and an exceptionally smooth transfer surface which may not accurately reflect spacecraft materials. Using the membrane transfer technique, the number of spores deposited per coupon is more complicated to calibrate and control, and still results in biphasic survival curves (i.e., also called tailing) to biocidal space conditions like UV irradiation (Fayolle et al., 2020), which suggests spores are clumping together.
Liquid-deposition methods considered here include varying concentrations of ethanol and varying surfactant solutions used to prepare spore suspensions that are deposited onto aluminum coupons. From our experience, the use of ethanol spreads out the area in which the spore suspensions are deposited on the coupons, which yields smoother monolayers than water. In addition, it is a common industry standard to ship Bacillus spp. spore suspensions in 40% ethanol, which makes it important to know the effects of ethanol on the quality of spore monolayers.
The use of surfactants to reduce strong interactions between ellipsoidal particles has been documented (Yunker et al., 2011; Krishna et al., 2016). A coffee ring effect is occasionally observed from liquid deposition, where spores spread out to the air-water interface and dry in position; however, this phenomenon is largely governed by the shape of the particles in the evaporated suspension (Yunker et al., 2011). Spherical particles deposited onto glass in liquid suspensions tend to migrate to the air-water interface at the edges of the droplets during evaporation to form a coffee ring, while ellipsoidal particles in evaporating droplets tend to congregate toward the center of the droplets with no ring formation. This is due to the strong interparticle attraction of ellipsoidal particles. Adding a surfactant at 1 × the critical micelle concentration (CMC) can eliminate the interparticle effects of ellipsoids and avoid the coffee ring effect. Adjusting the surfactant CMC may allow for better control of where spores in a droplet will settle after evaporation.
However, concerns exist that the addition of surfactants may result in spore/cell germination (Noell et al., 2013), particularly from non-ionic surfactants (Parker and Bradley, 1968; Setlow et al., 2003), cationic surfactants (Rode and Foster, 1960; Setlow, 2014), or anionic surfactants, such as sodium dodecyl sulfate (Yunker et al., 2011; Krishna et al., 2016). The denaturing properties of anionic surfactants are typically seen at higher concentrations (>0.1%). At concentrations as low as 1 × CMC, spore uniformity can be enhanced with minimal adverse effects.
The goals of the current study were to develop reliable methods for depositing bacterial spores on simulated spacecraft materials in monolayers for subsequent exposures to extreme biocidal conditions relevant to interplanetary space. The focus here was on (1) the use of surfactants to enhance spore monolayer quality while also not affecting the biocidal effects of UVC (220–280 nm) irradiation and (2) the effects of spore density on the lethality of a simulated equatorial martian UVC flux.
Methods
2.1. Preparation of Bacillus spores
Spores of Bacillus subtilis HA101 were prepared by the methods described by Mancinelli and Klovstad (2000) using the sporulation medium formulated by Horneck (1993). Spores were suspended in 10 mL sterile deionized water (SDIW) and stored at 4°C until used. The spores were enumerated with a Most Probable Number (MPN) assay (Mancinelli and Klovstad, 2000; Schuerger et al., 2003), and the stock suspensions contained ∼3.5 × 109 spores mL−1.
Biocidal effects of test media
If spores are to be deposited in liquid droplets onto coupons, it is important to determine the best carrier fluid for that task. The carrier fluid must not negatively impact spore survival or cause premature germination and growth in the absence of other nutrients. Both situations would affect the UV resistance of spores, and thus, resistance to interplanetary space conditions.
Tested carrier fluids included SDIW as a control, 15%, 25%, and 40% ethanol in deionized water, as well as the surfactants presented in Table 1: Tween 80, Triton X-100, and sodium dodecyl sulfate (SDS). Low concentrations of surfactants—half the critical micellar concentration (0.5 × CMC)—were prepared by serial dilutions in SDIW. All test media were filter-sterilized through 0.2 μm membrane filters prior to use. Liquid media were tested with and without added nutrients in the form of trypticase soy broth (TSB) to look for biocidal or growth-stimulation effects in nutrient-rich and nutrient-poor media.
Concentration of Surfactants Used as Test Media in Depositing Spores of Bacillus subtilis HA101 onto Aluminum Coupons to Achieve Smooth Monolayers
Concentration of Surfactants Used as Test Media in Depositing Spores of Bacillus subtilis HA101 onto Aluminum Coupons to Achieve Smooth Monolayers
First, the spore stock was diluted in SDIW to create inoculum of Bacillus subtilis spores at concentrations of ∼2.0 × 107 spores mL−1. Second, the inocula were diluted 1:100 in triplicate culture tubes containing each carrier fluid; each tube contained a T = 0 concentration of ∼2.0 × 105 spores mL−1. Cultures were incubated at 30°C while mixing at 120 rpm for 48 h. At the end of the incubation period, spore concentrations were measured with MPN assays, and values were compared to the starting concentrations of spores.
Aluminum coupons (special-order quote #25695183; Seton, Inc., Williamsville, NY, USA) with dimensions 17 × 54 × 1 mm were used as uncoated Al-coupons (UN) or coated with Chemfilm Class 3 (CL3) or Class 1A (1A) (coated by Synergy Metal Finishing, Titusville, FL, USA) as described by Schuerger (2022). All coupons were sterilized at 130°C in a convection oven for 24 h prior to use. Generally, the spore stocks were diluted to target concentrations of 2.0 × 107 spores mL−1 in SDIW, or one of the previously mentioned carrier fluids, and 100 μL of the suspensions were deposited onto each sterile coupon. The coupons were carefully moved to a stable and vibration-free benchtop surface to permit the settling of spores overnight (16–18 h). The following day, the carrier fluids were evaporated by exposing the applied droplets to a flow of sterile air within a biosafety cabinet (model NU-440-600, Class 2, Type A2, NuAire, Plymouth, MN, USA). After drying, spore monolayers measured approximately 1 cm in diameter on aluminum coupons.
Monolayer microscopy
The uniformities of dried spore layers deposited on all three types of Al-coupons were imaged under a dissecting microscope (Zeiss Stemi 2000C, White Plains, NY, USA) equipped with a digital camera (Jenoptics ProgRES Speed Xt3, Jupiter, FL, USA). Select coupons were observed with scanning electron microscopy (SEM) at the University of Florida's Interdisciplinary Center for Biotechnology Research (ICBR), Gainesville, Florida. Spores on coupons were first inactivated by exposure to UV for 4 h under UVC flux of 4 W/m2 (200–280 nm), coated with gold-palladium in a sputter coater (Denton Vacuum Sputter Coater, model Desk-V, Moorestown, NJ, USA), and then imaged in a Hitachi S-4000 SEM (Hitachi, Ltd., Greenville, SC, USA). UV irradiation of the spores ensured the safety of personnel in the ICBR/SEM facility. Spores were not fixed prior to SEM imaging.
Spore recovery from coupons
Spores on coupons were extracted in sterile 50 cc polystyrene conical tubes (Falcon or Corning tubes, Fisher Scientific, Pittsburg, PA, USA) with 20 mL of SDIW and 1 g of sterile silica sand (SDIW + silica sand), as described by Schuerger (2022). Each sample was vortexed at the maximum setting for 2 min, with vigorous shaking every 30 s. The spores extracted from coupons were enumerated using the MPN assay.
Attachment and recovery of spores deposited with ethanol
Bacillus subtilis HA101 spores were diluted in SDIW, plus 15%, 20%, 25%, or 40% ethanol. Liquid suspensions of spores were deposited in 100 μL aliquots onto UN, 1A, and CL3 coupons at ∼2.0 × 106 spores per coupon. Within 1 week of preparing the coupons, viable spores were extracted from each coupon using the SDIW + silica sand protocol (henceforth called the standard MPN assay).
Attachment and recovery of spores deposited with surfactants
Bacillus subtilis spores were diluted in SDIW or the surfactants Tween 80, SDS, or Triton X-100 diluted to 0.5 × CMC. Six coupons of each material (UN, 1A, and CL3) were doped with 100 μL of one of the surfactant-spore mixtures with ∼2.0 × 106 spores per coupon. Viable spores were subsequently extracted from each coupon at random and enumerated by the standard MPN assay. The number of spores recovered divided by the initial number of spores (expected) was plotted as log10 (N/N 0) values.
Mars UV exposures
Mars-surface UV irradiation under clear sky conditions was simulated in a Planetary Atmospheric Chamber (PAC) (previously called the Mars Simulation Chamber by Schuerger et al., 2008) using a xenon-arc UV lamp (model 6269, Oriel Instruments, Stratford, CA, USA). To calibrate UVC flux to Mars-normal conditions of 3.2 W m−2, the exposure time within the chamber was scaled to account for the difference between the measured UVC flux of the xenon-arc UV lamp, and the desired UVC dose. Exposures were conducted under standard temperature (24°C) and pressure (101.3 kPa) with an Earth-normal gas composition (i.e., 78% pN2, 21% pO2, and trace gasses). The simulated equatorial Mars UV model has been described previously by Moores et al. (2007) and Schuerger et al. (2003, 2006).
UV exposure of smooth versus uneven spore layers
To determine the effectiveness of UV sterilization on smooth versus unevenly distributed spore layers, three experiments were developed. Each experiment was conducted with 2 spore-layer preparation methods with 6 replicates sampled at 8 different time points (0, 1, 5, 15, 30, 60, 120, and 180 min) for a total of 96 data points in each experiment. Prior to UV exposures, the numbers of spores recovered at T = 0 from all coupon preparation protocols (i.e., Class 1A + Triton, 1A + SDIW, etc.) were enumerated, and the average numbers of spores (n = 6) served as N 0.
Experiment-1. Spores suspended in SDIW were deposited in 100 μL aliquots on uncoated-Al and Class 1A coupons at ∼2.0 × 106 spores per coupon. Coupons were placed in random positions under the UV beam and irradiated in the PAC. At each time-step, six coupons of each carrier fluid were removed from the UV beam. Spores were recovered and enumerated via the standard MPN assay; the numbers of viable spores recovered divided by the numbers of spores initially deposited onto coupons were plotted as log10 (N/N 0) values.
Experiment-2. To test for UV-shielding effects by surfactants used to deposit spores, Class 1A coupons were doped with ∼2.0 × 106 spores/coupon deposited with either SDIW or 0.5 CMC Triton X-100, both of which formed similarly uniform layers. Coupons were placed in random positions under the UV beam and irradiated in the PAC. At each time-step, six coupons per carrier fluid were removed from the UV beam. Spores were recovered and enumerated via the standard MPN assay. The numbers of viable spores recovered divided by the numbers of spores initially deposited onto coupons were plotted as log10 (N/N 0) values.
Experiment-3. To test for material interactions with surfactants, spores suspended in Triton X-100 were applied on uncoated or Class 1A coupons at ∼2.0 × 106 spores per coupon, both of which formed similarly uniform layers. At each time-step, six coupons from each carrier fluid were removed from the UV beam. Spores were recovered and enumerated via the standard MPN assay. The numbers of viable spores recovered divided by the numbers of spores initially deposited onto coupons were plotted as log10 (N/N 0) values.
UV exposure of spore layers with diverse spore densities
Spores were deposited on uncoated Al-coupons using spores suspended in Triton X-100 at three different spore densities averaging 1.69 × 106, 1.32 × 107, or 1.16 × 108 spores per coupon. Coupons were imaged with light-microscopy and SEM, as described above. Coupons doped with each spore density were exposed to Mars-normal UV radiation for the equivalent of 0, 2, 8, 24, 48, 96, or 168 h. At each time point, six coupons of each spore density were removed from the UV beam, and the spores were recovered and enumerated by the standard MPN assay. The number of viable spores surviving each dose of UV radiation were divided by the number of spores initially deposited and are reported as log10 (N/N 0) values.
Statistics
Statistics were performed in RStudio Version 1.3.1093. Data from all experiments were screened by the removal of outliers and assessments of both normality and homogeneity of variance conducted. To correct for nonhomogeneous variances of treatment means, log-transformed data were most often used in the analyses. The data from the ethanol and surfactant attachment and recovery experiments were both subjected to ANOVA, followed by least-squares mean separation (LSmeans) tests for multiple comparisons where applicable. Data for spore-density assays versus UV irradiation, and for viability of UV-exposed smooth versus uneven monolayers, were modeled in piece-wise functions using LINEST in Excel. Spline points were calculated using an open-source online package available at
Results
Biocidal effects of test media
None of the carrier fluids used to apply spores to coupons resulted in significant gain or loss of spores over 48 h when incubated in the carrier fluids without nutrients (Fig. 1, dark bars). Additionally, none of the ethanol carrier fluids without nutrients supported growth at 48 h. When incubated with added nutrients (Fig. 1, light bars), however, all concentrations of ethanol significantly inhibited growth in TSB (P > 0.05) compared to SDIW (P ≤ 0.0001) with nutrients. The use of Tween 80 or Triton X-100 surfactants showed no significant inhibitory effects on growth of B. subtilis spores in the absence of TSB (P > 0.05) compared to the SDIW with nutrients. Interestingly, SDS with added nutrients resulted in a significant loss of viable spores (i.e., greater than 3 orders of magnitude [henceforth given as logs] compared to the SDIW controls; P ≤ 0.0001). Estimations in growth of the bacterial populations in all samples were based on changes in the MPN assay results; we did not directly observe germination of spores.
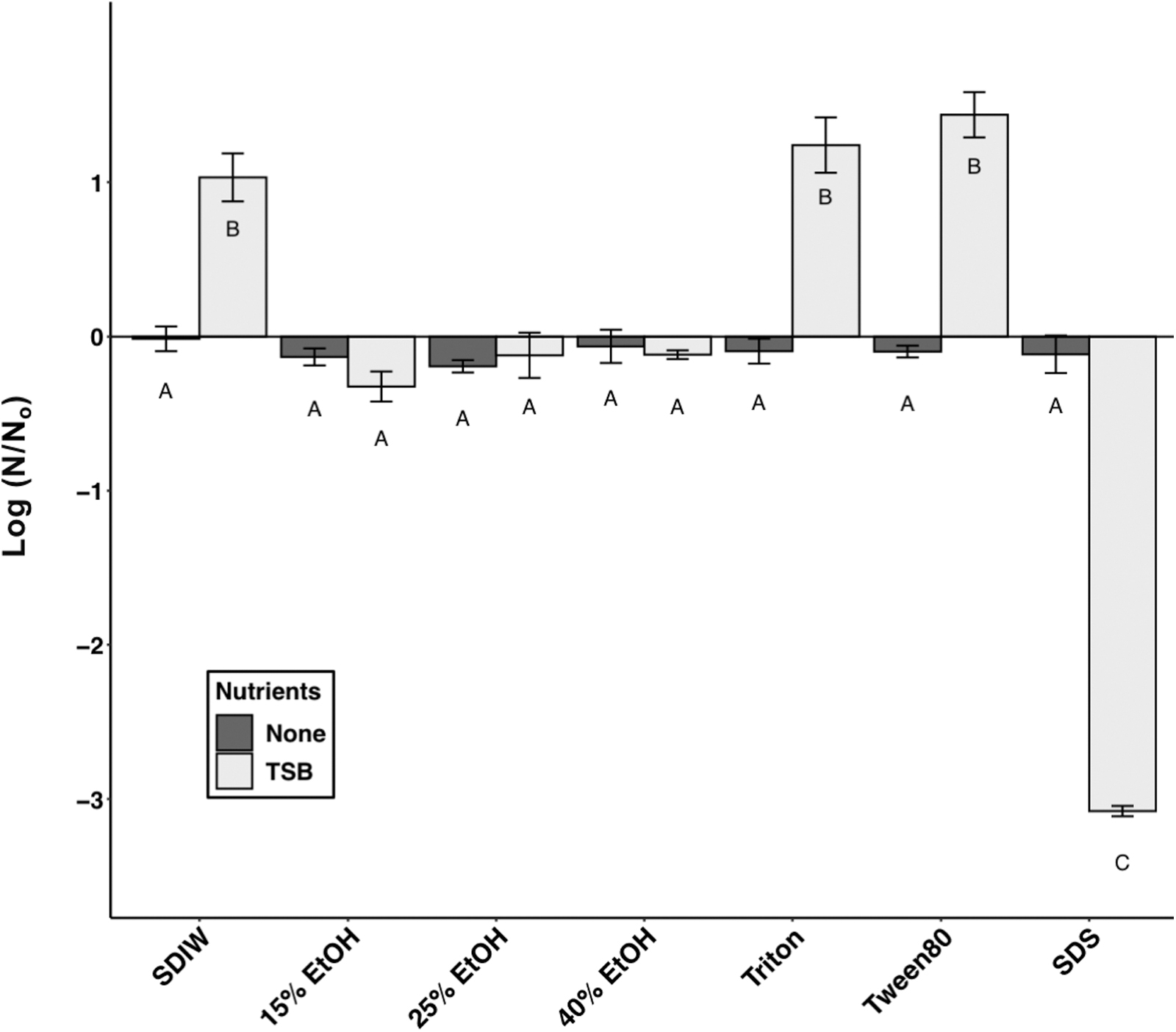
Growth or loss of viable spores of Bacillus subtilis HA101 in nutrient-poor (dark gray bars) and nutrient-rich (TSB-supplemented; light gray bars) liquid media. Sterile deionized water (SDIW) served as the control (0-line on y-axis). Letters are used to represent different groups based on ANOVA and protected LSmeans tests (P ≤ 0.05; n = 3; data in Supplementary Table S1). Those marked “A” are not significantly different from the control SDIW, and the spore concentrations did not change from the initial densities deposited. Error bars are standard errors of the means.
Three combinations of coupon material and carrier fluids resulted in consistently smooth monolayers upon visual inspection. The best combinations of fluids and coupon materials were spores deposited onto (1) Class 1A coupons in SDIW droplets (Fig. 2C) and (2) spores deposited in Triton X-100 droplets on either UN (Fig. 2M) or 1A (Fig. 2O) coupons. Spores deposited in SDIW on UN and CL3 coupons resulted in irregular distributions of spores on >90% of the samples. Coupons doped with Tween 80 or Triton X-100 were generally smooth (Fig. 2G, 2H, 2I, 2N), but these desired results were only observed on approximately two-thirds of the test coupons for each carrier fluid. In contrast, coupons doped with spores suspended in SDS—regardless of surface material—resulted in high-density spore regions (e.g., Fig. 2J, 2K, 2L).
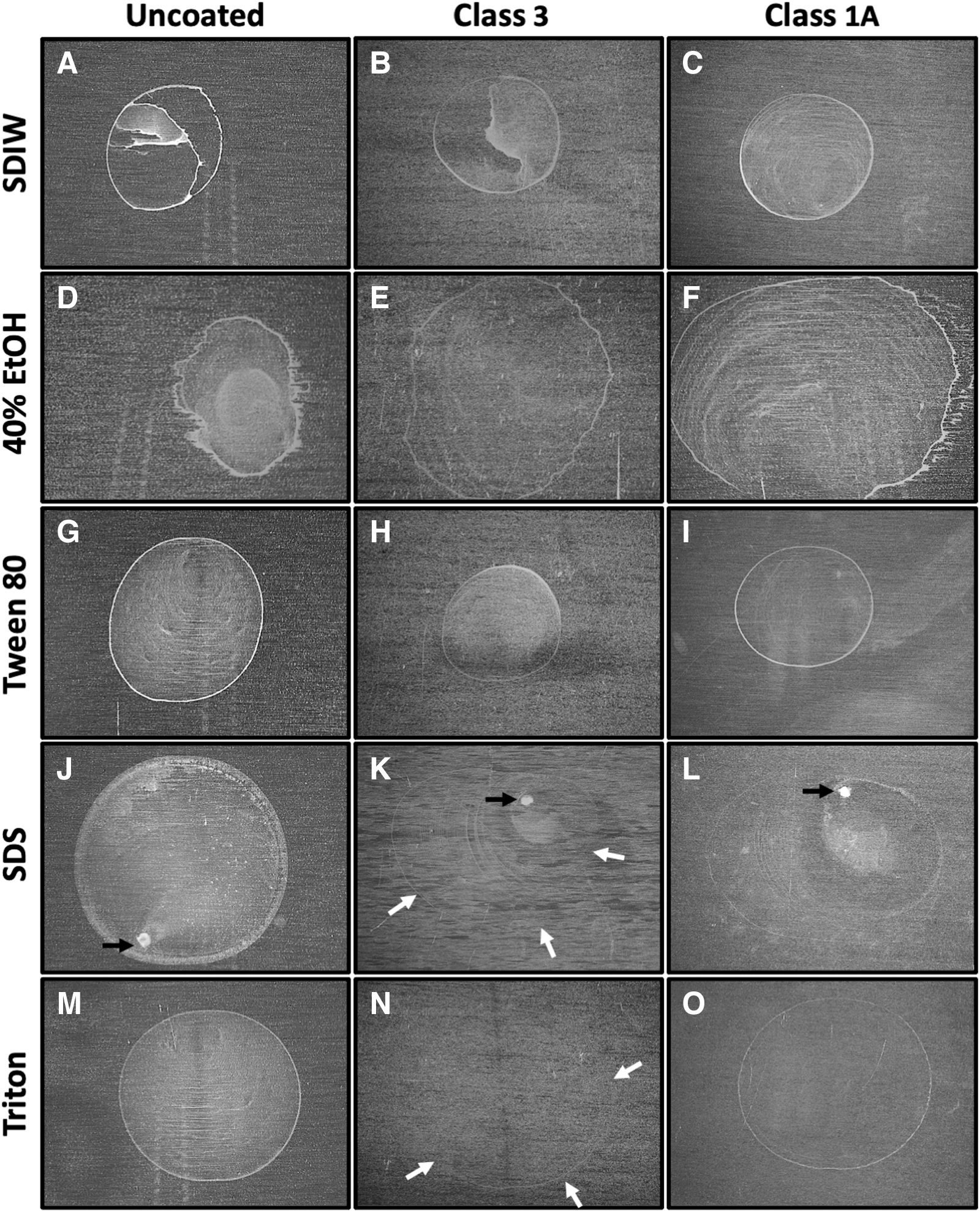
Images of dried Bacillus subtilis HA101 spore monolayers deposited in sterile deionized water (SDIW), 40% ethanol (EtOH), 0.5 critical micelle concentration (CMC) Tween 80, 0.5 CMC sodium dodecyl sulfate (SDS) and 0.5 CMC Triton X-100 on uncoated, Class 3, and Class 1A coupons. Dense concentrations of spores are indicated below by black arrows in spore layers prepared in SDS. Faint edges (often called a “coffee-ring” effect) of spore layers are highlighted by white arrows. Good monolayers were defined as those with minor coffee-ring deposits of spores and with smooth spore deposits within the edges of the deposited monolayers (best examples are panels N and O).
Spores suspended in ethanol and deposited in droplets on coupons had slightly lower recovery at higher concentrations of ethanol regardless of the material (Fig. 3). A significant effect of concentration overall was observed (P = 0.0096), while no overall effect of material was observed (P = 0.1037). The LSmeans test revealed significant differences between recovery of spores deposited in SDIW versus spores deposited in 40% ethanol (P = 0.0101). Although the LSmeans tests gave significant trends, the divergence from the Time = 0 controls (Fig. 3; dashed lines) remained less than one-half of 1 log. The precision of the MPN assay has been estimated to be approximately 0.25 logs (Schuerger et al., 2003, 2006); thus, the differences above were close to the precision limit of the MPN assay.

Recovery of Bacillus subtilis HA101 spores from coupons prepared with diverse ethanol concentrations. Data were analyzed with ANOVA and protected LSmeans tests comparing each data set to the Time = 0 controls (dashed line) (P ≤ 0.05; n = 6; data in Supplementary Table S2). Error bars are standard errors of the means.
Spores suspended in SDIW, Tween 80, and Triton X-100 were similarly recoverable from all tested materials (Fig. 4). Overall, there were no significant effects of surfactants on recoverability of spores (P = 0.1975) (except SDS on uncoated coupons). Conversely, spores deposited in SDS droplets on uncoated coupons were the least recoverable, resulting in a loss of slightly larger than 0.25 logs of spores (P ≤ 0.05).
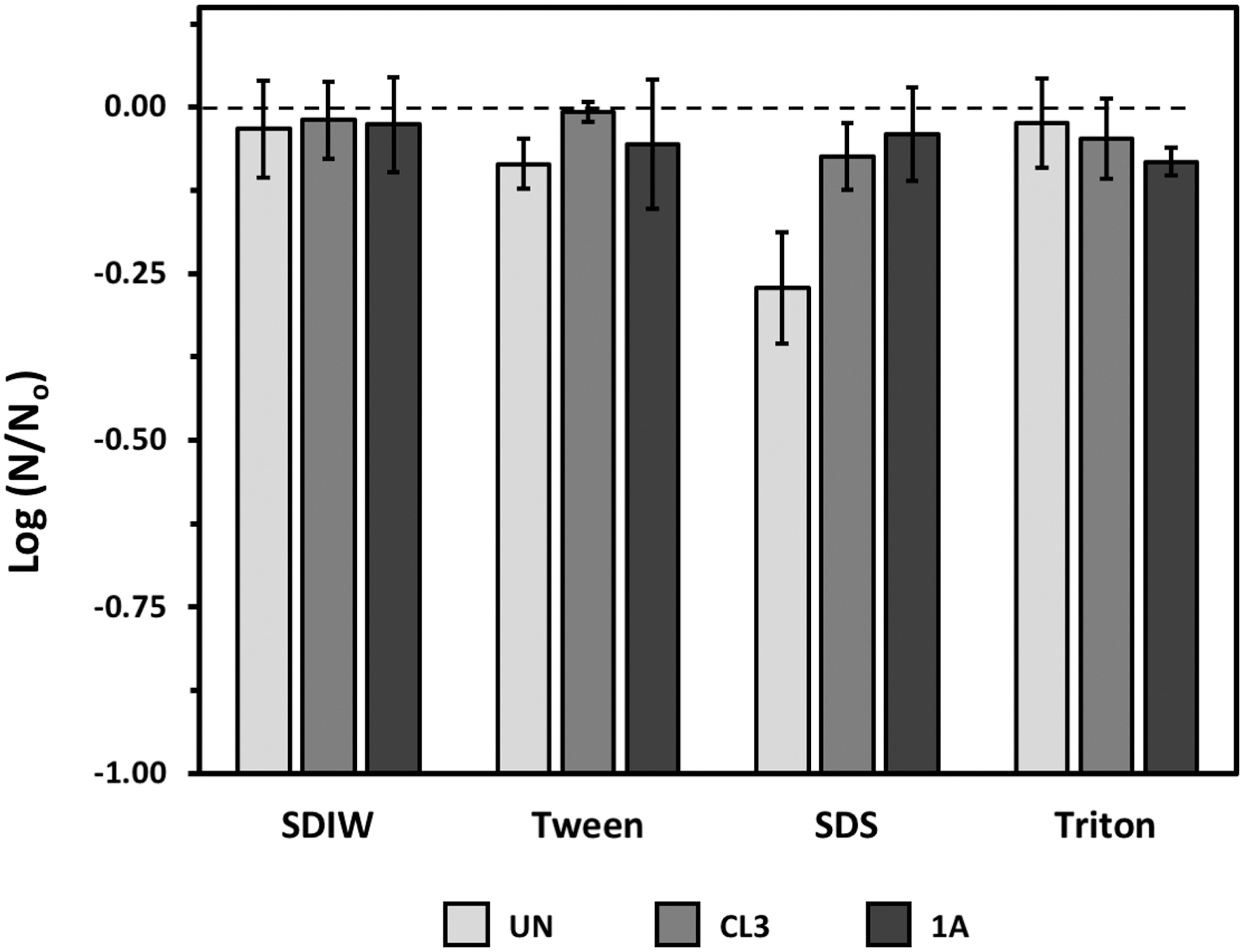
Recovery of Bacillus subtilis HA101 spores from coupons prepared with different surfactants—Tween 80, SDS, and Triton—as the deposition media. Sterile deionized water (SDIW) was used as a control carrier fluid. Recovery of spores are plotted from uncoated coupons (UN; light gray bars), from Class 3–coated coupons (CL3; mid-gray bars), or from Class 1A–coated coupons (1A; black bars). Data were analyzed with ANOVA and protected LSmeans tests comparing each data set (i.e., bar) to the Time = 0 control (dashed line) (P ≤ 0.05; n = 6; data in Supplementary Table S3). Error bars are standard errors of the means.
In Exp-1, survival of spores in uniform monolayers (e.g., SDIW on Class 1A coupons in Fig. 2C) was compared to survival of spores in irregular layers (SDIW droplet on UN coupons in Fig. 2A). After 15 min of UV exposure, the uniform monolayer exhibited a loss of survivors by approximately 4 logs, while the irregular layers exhibited only 2 logs of lost viability (Fig. 5A). By 180 min, both treatments yielded a loss of 1 and 3 additional logs, respectively. Neither treatment was observed to completely inactivate all spores on the coupons at 180 min. The uniform monolayers and the irregular layers resulted in significantly different phase-1 slopes (P < 0.0001) and phase-2 intercepts (P < 0.0001), but the phase-2 slopes were not significantly different (P = 0.6374).
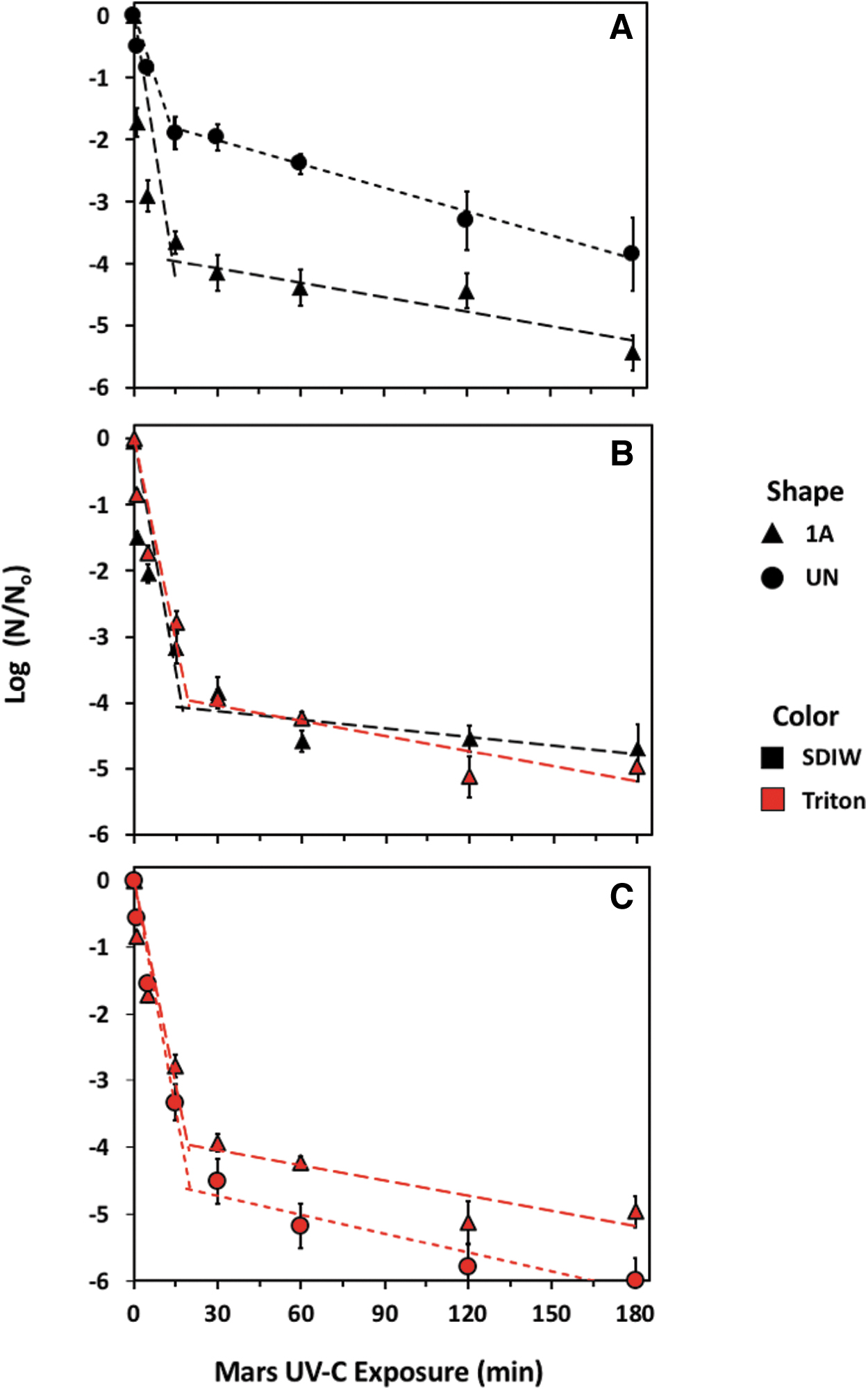
Effects of UV irradiation on monolayers of Bacillus subtilis HA101 spores depicted in Fig. 2. Experiments included (
In Exp-2, the effects of UV on spore layers deposited onto the same materials with different carrier fluids were investigated to determine whether the carrier fluids result in any inherent UV-shielding of spores (Fig. 5B). Spores were deposited onto Class 1A coupons with SDIW and Triton X-100 droplets in which both carrier fluids consistently formed uniform monolayers. In both treatments, viability of spores was reduced by 4 logs in the first 15 min of UV exposure, which was also observed for the uniform monolayer in Fig. 5A. In the remainder of the 180 min experiment, both treatments had decreased viabilities of approximately 5 logs. The viabilities of spores over time for both treatments were similar between phase-1 slopes, phase-2 slope, and spline point intercepts (P > 0.05).
In Exp-3, the effects of UV on spore monolayers created on different materials with Triton X-100 were investigated to determine whether spores exposed to UV irradiation would yield divergent biocidal kill-curves over 180 min (Fig. 5C). Spores were deposited on UN and Class 1A coupons in droplets containing Triton X-100. After 15 min, viability decreased approximately 4 logs for spores on Class 1A coupons and 4.5 logs for spores on UN coupons. Over the remainder of the 180 min experiment, viable spores on Class 1A coupons decreased another 1 log for a total of 5 logs for the 3-h exposure, while viable spores on UN coupons decreased to 0 at the end of the exposure window (no viable spores were detected). While the endpoint carries biological relevance, no statistically significant differences between the two piece-wise regressions were observed in the phase-1 slope, phase-2 slope, or spline point intercepts (P > 0.05). The modeled regression models and relevant statistics are given in Table 2.
Piece-wise Functions of Log-Transformed Data Serving as Models of UV-Exposed Smooth Versus Uneven Monolayers of Bacillus subtilis HA101 Spores
Models were prepared using LINEST in Excel, while spline points were calculated in
Varying concentrations of spores in Triton X-100 droplets on coupons yielded divergent results with clear differences in monolayer opacity due to different cell densities (Fig. 6). Spore monolayers created with 1.69 × 106 spores of B. subtilis per coupon (Fig. 6A) exhibited smooth monolayers in which the background metal surface could be easily seen. In contrast, >90% of the metal background was obscured by spore layers created with 1.32 × 107 spores/coupons (Fig. 6B). And lastly, spore layers created with 1.16 × 108 spores/coupons exhibited fully opaque multiple layers of spores on aluminum coupons (Fig. 6C).
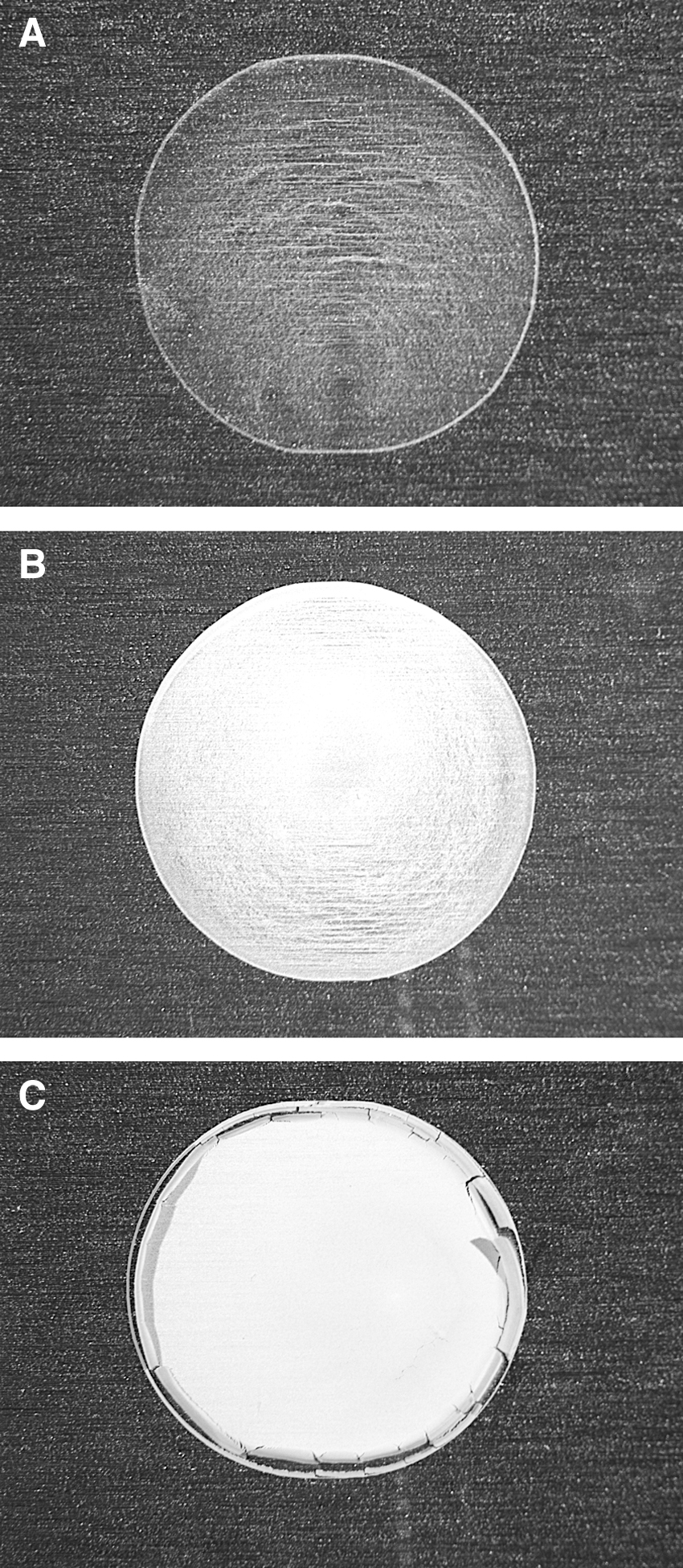
Dried spore layers of Bacillus subtilis HA101 on uncoated aluminum coupons deposited in a spore mixture prepared as 0.5 CMC Triton X-100 and three different spore densities. The average numbers of spores on coupons were (
Subsets of the above cell densities per coupons (i.e., henceforth 106, 107, and 108 per coupons) were imaged with SEM to characterize the spore distribution patterns for each density. Spores on coupons containing both 106 spores and 107 spores were distributed relatively evenly across the coupons (Fig. 7A and 7B, respectively). Stacking of spores was still observed in some subsections of the monolayers with 107 spores/coupon. However, and in sharp contrast to the 106 and 107 doped coupons, spore layers with 108 spores (Fig. 7C) resulted in stacked layers of spores that were measured between 8 layers (Fig. 7D) and 15 layers of B. subtilis spores (data not shown).

Scanning electron microscope images of spores deposited on uncoated aluminum coupons using Triton X-100 at spore densities of (
The different cell densities under UV exposures resulted in varying degrees of UV inactivation of spores (Fig. 8). First, while some irregularities in distributions were observed in deposits with 106 spores (Figs. 6A and 7A), these results were consistent with the monolayers from the previous experiment exhibiting a 5-log reduction in viability in just 2 h of UV exposure (Fig. 8A). Total losses of viable spores in 106 coupons (i.e., no spores were recovered in the MPN assays) were observed after 24 h of UV exposures but may have occurred sooner had the sampling intervals been shorter.

Exposure of Bacillus subtilis HA101 spore layers at three different densities to Mars-normal UV irradiation. The average numbers of spores on coupons were (
Aggregated layers of B. subtilis with 107 spores were visibly denser than cohorts of coupons with 106 spores (compare Fig. 7A and 7B). In the 107 scenario, approximately 1-log of reduction was observed after 2 h, followed by a gradual reduction of viable spores for the remainder of the 168 h exposure for a total 5-logs lost in viable spores (Fig. 8B). In this case, there were still approximately 102 spores remaining on each coupon after 168 h of exposure. Notably, the slopes of the phase-2 regression models of viable spores for 106 spores and 107 spores were similar (Table 3).
When 108 spores were deposited in Triton X-100 droplets, the edges of the spore layers often curled after evaporation of the carrier fluid and tended to flake off the coupons (Fig. 6C). Although we attempted to use only coupons with intact spore layers, the flaking process may have partially contributed to the loss of viable spores observed for 108 spore-doped coupons. For the 108 spore-doped coupons, a 1-log reduction was observed in samples comparing the 0 and 24 h time-steps (Fig. 8C), but no further losses in viable spores were observed between 24 and 168 h under continuous UV irradiation. All regression models for Fig. 8 data are given in Table 3.
Piece-wise Functions of Log-Transformed Data Serving as Models of UV-Exposed Mono- Versus Multilayers of Bacillus subtilis HA101 Spores
Models were prepared using LINEST in Excel, while spline points were calculated in
Quantitative models for microbial survival in space (e.g., Dillon et al., 1973; Schuerger et al., 2019; Moores and Schuerger, 2020) are predicated on precise kill-kinetics of microbial cells and spores exposed to biocidal factors in robust simulations on (1) how microorganisms reside on spacecraft surfaces and (2) how each surface is exposed to diverse biocidal conditions. Survival rates for external spacecraft surfaces will differ greatly from survival rates in protected niches within the spacecraft (Schuerger et al., 2019). In addition, if extant spacecraft microbiomes prior to launch are best represented by single spores or very small clusters of spores—a missing link in our current understanding of spacecraft microbiomes—then survival rates of multilayered aggregates of spores (e.g., Horneck et al., 2012; Panitz et al., 2015) are likely to yield biased results.
The research presented here is from a series of studies that have attempted to develop and calibrate protocols to prepare uniform spore monolayers via droplets of spore suspensions on simulated spacecraft materials and examine the effects of surface texture, UV irradiation, vacuum, and use of surfactants to assist in the creation of monolayers. Two previous studies in this series have been published (Schuerger, 2022; Schuerger et al., 2023).
First, of the simulated spacecraft materials tested here and by Schuerger (2022), uncoated aluminum 6061 coupons, and Al coupons coated with Chemfilm Class 1A films, proved superior to coupons with Class 3 Chemfilm coatings. The latter Class 3 coating was excessively rough and provided significant protection for Bacillus spores in cracks and overhangs embedded within the chromium oxide film (see Fig. 1 in Schuerger, 2022). Of these two studies (Fig. 2 here; Schuerger, 2022), uncoated Al-coupons with a SDIW carrier fluid or uncoated- or Chemfilm-prepared coupons with Triton X-100 proved to yield consistently uniform and smooth monolayers of spores. Triton X-100 did not deposit a coating on spores, and it did not affect the UV biocidal properties on spore survival (Fig. 5 here). Furthermore, the surfactants tested here failed to provide any stimulation for spore germination or growth, except when the organic food-base—TSB—was added to the media (Fig. 1 here). Thus, we conclude that uniform and reliable monolayers can be prepared with the droplet protocol by using either SDIW + Chemfilm Class 1A or Triton X-100 at 0.5 × CMC with either uncoated or Class 1A coupons.
In creating spore monolayers of various Bacillus spp., some have claimed that significant coffee-ring effect rimes can form multilayered deposits on the outer edges of the droplets (Levy et al., 2011; Noell et al., 2013; Raguse et al., 2016; Schuerger, 2022). The coffee-ring effect rimes have been attributed to the shape of particles in which ellipsoidal particles are able to dry in a more dispersed pattern than spherical particles (Yunker et al., 2011). In the data presented here (Figs. 2 and 6), the coffee-ring effect was not eliminated by Triton X-100 but was greatly subdued. The coffee-ring rimes of drying spores may create multilayered aggregates that might explain the tailing effects observed in UV-irradiation kill-curves (see Figs. 5 and 8 here; Cerf, 1977; Schuerger et al., 2003, 2006; Fayolle et al., 2020).
Second, recovery efficiency of spores from coupons is a critical component in calibrating biocidal assay data intended for populating microbial survival models for spacecraft. Schuerger (2022) demonstrated that the addition of 1 g of pre-sterilized silica sand to 50 cc conical tubes used to remove spores from aluminum coupons boosted recovery rates by 1 log. In assays in which only a liquid carrier fluid was used (e.g., Shuster et al., 2019; Schuerger, 2022), recovered spores from surfaces were generally 1–1.5 logs lower than the starting cell densities in liquid cultures used to create monolayers. To enhance the recovery of spores, a polyvinyl alcohol (PVA) assay has been developed (e.g., Lindberg and Horneck, 1991; Xue and Nicholson, 1996). In a direct comparison between the PVA and standard MPN assays, Schuerger et al. (2002) demonstrated that both assays were on par with each other and yielded close to 100% recovery of Bacillus spp. spores.
Third, we recommend that lower doses of spores (∼2 × 106 spores per coupon) are more appropriate in biocidal assays for spacecraft survival models than higher doses of spores (∼2 × 108 spores per coupon)—especially for UV-irradiation assays—because at the higher range, spores develop thick multilayers in which the upper spores will attenuate UV photons, protect deeper layers of spores (Fig. 8 here), and thus bias the results toward higher survival rates. For example, with low-density (∼2 × 106) spore monolayers, Schuerger et al. (2006) demonstrated that survival dropped below the detection limit of one viable spore per coupon for seven Bacillus spp. exposed to only 180 min of simulated equatorial martian UVC flux. In contrast, high-density spore multilayers of B. subtilis 168 and B. pumilus SAFR-032 (both at ∼3.6 × 108 spores per coupon) in the EXPOSE-E (Horneck et al., 2012) and EXPOSE-R (Panitz et al., 2015) experiments exhibited significant survivors after 1.5 and 2 years in space, respectively. The survivors in the Horneck et al. (2012) and Panitz et al. (2015) studies were likely deeply embedded spores surrounded and protected by upper spore layers.
The data presented here in Figs. 6, 7, and 8 support the conclusion that UV kill-curves with monolayers at ∼2 × 106 spores per coupon yield significantly more realistic biocidal inactivation rates for spacecraft surfaces than multilayered spore survival rates from thick ∼108 spores per coupon because it is much more likely that spacecraft microbiomes are composed of individual spores or small clusters of spores. The 108 spore multilayers were estimated to be composed of up to 10 layers of B. pumilus SAFR-032 (Panitz et al., 2015) and 15 layers here for B. subtilis HA 101 (9 layers shown in Fig. 7 here, but other counts were as high as 15 layers on a coupon, data not shown). Based on the data presented here and by Schuerger (2022), we propose an upper limit of 5 × 106 spores per coupon (≤ 5 × 106 spores per cm2) for UV-irradiation experiments in which the results are intended to populate microbial survival models for interplanetary or planetary spacecraft. Higher densities of spores are likely to bias results to unusual high survival rates for Bacillus spores on spacecraft surfaces.
And lastly, reviewing the data in Fig. 8 yields several subtle conclusions that should be examined: first, survival kill-curves for the 106, 107, or 108 spore densities form biphasic kill-curves in which phase-1 is steep over short time steps, followed by refractory phase-2 models over longer time steps. For the 106 (Fig. 8A) and 107 (Fig. 8B) spore densities, we propose that the tailings are due to stacking of spores in small aggregates, in cracks or pits, or multilayers in the coffee-ring rimes. As the density increases, so does the stacking—followed by a shallowing of the phase-1 and phase-2 kill-curves. However, even at 108 spores per coupon, a significant 1-log decrease in spore survival was observed over the first 24 h of UV exposure, suggesting that most of the upper 1–2 layers of spores were inactivated by UV irradiation. The phase-2 linear model decreased slightly between 24 and 168 h (Fig. 8C); thus, it is plausible that deeper layers of spores were being inactivated over time.
Conclusions
Creating uniform monolayers of microbial vegetative cells and spores is a critical step in developing robust microbial survival models for interplanetary spacecraft. The data presented here are derived from a series of protocols papers (see also Schuerger, 2022; Schuerger et al., 2023) that have sought to develop methods for creating uniform monolayers, exposing Bacillus spp. spores to simulated biocidal space conditions, and recovering close to 100% of the spores applied to aluminum coupons. These data suggest that low-density spore monolayers (i.e., 2–5 × 106 spores per coupon) yield reliable predictions of inactivation kinetics for UV irradiation assays because the spores do not aggregate into clumps. Protocols that either cause aggregation (e.g., surface hydrophobicity that enhances clumping; Fayolle et al., 2020; Schuerger et al., 2005), enhance layering at high spore densities (e.g., data in Fig. 8 here; Horneck et al., 2012; Panitz et al., 2015), or use materials with rough surface textures (e.g., Edmonds et al., 2009; Schuerger, 2022; Schuerger et al., 2023) will bias results to suggest that spore survival rates after UV exposures are higher than would be predicated on actual spacecraft surfaces in which single cells/spores or very small clusters of cells/spores are the norm.
Based on the data presented here—and by Schuerger (2022) and Schuerger et al. (2022)—overall recommendations for spore-survival assays are as follows: (1) Use low-density (2–5 × 106 spores per cm2) monolayers of spores on smooth surfaces for UV irradiation experiments; (2) Do not use monolayers that are more than 3 weeks old (Schuerger, 2022); (3) Surfactants like Triton X-100 can be used to enhance the creation of smooth monolayers (Fig. 5 here); (4) UV exposures from deuterium, Hg-line, or xenon-arc lamps appear to yield comparable results for inactivation kinetics (Schuerger, 2022); (5) PVA and the standard MPN assays with 1 g of silica sand added to extraction tubes are comparable to each other and yield similar extractions close to 100% (Schuerger et al., 2023); (6) Investigators should be cautious of biocidal volatile organic carbon (i.e., VOCs) in assays—especially in sealed containers (Schuerger et al., 2023)—that can act to artificially increase kill-rates for Bacillus spp.
And finally, the protocols discussed above are not final. A key objective here (and in Schuerger, 2022) is to stimulate a discussion in the spacecraft processing and planetary protection communities to standardize protocols to such an extent that data generated by divergent labs can be directly compared. The long-term goal should be to develop robust microbial survival models for interplanetary and planetary spacecraft that will quantitatively estimate bioburden survival rates when such data are critical for mission success. These data are presented as a series of steps in that direction.
Footnotes
Author Contributions
ACS and ELH envisioned the research, jointly prepared and edited the manuscript, conducted the statistics, and created the figures. ELH conducted the experiments presented here.
Acknowledgments
We thank Karen Kelly at the Interdisciplinary Center for Biotechnology Research SEM labs on the main campus of the University of Florida (Gainesville, FL) for assistance in the SEM imaging.
Author Disclosure Statement
The authors have no financial, commercial, or other relationship conflict of interest with any organization listed in the text.
Funding
Funding support for the creation of this work was provided by the National Aeronautics and Space Administration (NASA), Planetary Protection Office, NASA Headquarters, Washington, DC, USA (grants #NNX12AJ84G; 80NSSC17K0263; 80NSSC20K0745).
Data Management Plan
Raw data for Figures 1, 3, 4, ![]() , and 8 are given in the attached supplementary files. In addition, raw data from the current study are available in a University of Florida Institutional Repository (UFIR) for ACS at the link
, and 8 are given in the attached supplementary files. In addition, raw data from the current study are available in a University of Florida Institutional Repository (UFIR) for ACS at the link
Supplementary Material
Supplementary Table S1: Fig 1 Raw Data
Supplementary Table S2: Fig 3 Raw Data
Supplementary Table S3: Fig 4 Raw Data
Supplementary Table S4: Fig 5 Raw Data
Supplementary Table S5: Fig 8 Raw Data
Abbreviations Used
Associate Editor: Petra Rettberg
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.