
Abstract
Increasing antibiotic resistance (AR) poses dangers of treatment complications and even treatment failure to astronauts. An AR determinant is a gene of resistance carried by bacteria. This article considers the issue of the stability of AR determinants and the influence of manned spaceflight conditions on this characteristic. A phenomenological model has been developed that makes it possible to evaluate the integral value of the stability of determinants of AR in bacteria as a function of time. Based on experimental results obtained during implementation of the SALYUT 7 space program, the stability of determinants of AR in Escherichia coli strains isolated before and after a spaceflight in 16 astronauts was evaluated. In addition, an assessment was made of the integral value of the stability of determinants of AR in bacteria during in vitro experiments, both in spaceflight and terrestrial conditions, after preincubation in space. The calculation using the developed phenomenological model showed that the stability of AR determinants in E. coli bacteria isolated from astronauts before the spaceflight is 33% higher than after the flight. The in vitro experiment carried out on board the International Space Station showed the opposite situation—an increase in the stability of AR determinants by 33% in cultures that have been in space compared with terrestrial control. This indicates an additional influence on the stability of determinants and of the astronaut's immune system, as well as space conditions. The common result in these two types of studies is the experimental fact that the largest number of bacteria, in space conditions, had two determinants of AR. The importance of fighting bacteria with two determinants is that at least three different antibiotics are required to have an effect. This circumstance makes it possible to predict a possible strategy for the use of antibiotics in autonomous spaceflights.
1. Introduction
Increasing antibiotic resistance (AR) poses the danger of antibiotic therapy failure, leading to adverse outcomes of increased treatment duration, increased need for intensive care, surgery, functional decline, and even mortality (Friedman et al., 2016). In particular, the threat to astronauts is higher since there are limited medical resources on board the space shuttle/station and a possibility of compromising long-distance space missions. The adverse outcomes are also economical due to increased medical resource utilization and cost of treatment and possible mission failure.
When analyzing data on AR in bacteria during long-term isolation of a person in a hermetic facility, for example, on an orbital space station, it is necessary to select those situations that are closest to the conditions of an autonomous or interplanetary flight. Under the current operating conditions of the International Space Station (ISS), it is difficult to take it as a model because it does not have classical isolation due to frequent crew changes. Therefore, for us, the most interesting object in this respect is the SALYUT 7 station, on which a series of autonomous flights was carried out.
A retrospective analysis of data on bacterial AR under conditions of a long-term spaceflight (mission—SALYUT 7) showed that bacterial AR decreases with respect to terrestrial conditions (Ilyin et al., 2022). However, the question of the stability of AR determinants under extreme spaceflight conditions remains open since these conditions can affect not only the stability of AR determinants but also to a large extent various mechanisms of horizontal gene transfer (HGT) and carriers that implement HGT. This influence is possible because HGT is largely determined by the environment and external factors (Martínez, 2012).
There are several carriers that implement HGT (Salyers and Amábile-Cuevas, 1997): the self-transmissible plasmid; conjugative transposon; mobilizable plasmid; nonreplicating Bacteroides unit; transposon; gene cassette; integron; and others. Each of these carriers becomes dominant under certain external conditions (for more information, refer to summary of terms below). Under the action of ionizing radiation, weightlessness, and limiting factors of hermetic objects (conditions typical for autonomous manned spaceflights), one should also consider the HGT pathway in AR transmission, in which pathogenic microflora can use the genetic material released as a result of lysis of various cells (Cooper et al., 2017).
Summary of elements and processes that confer antimicrobial resistance in bacteria
In addition, it should be taken into account that in a closed system, the main factor in AR stabilization is the ratio of the biological fitness cost of resistance and the biological fitness cost of adaptability, that is, the ability of the pathogen to reproduce and spread in the host population (Andersson and Hughes, 2011). The presence of polyresistance in Escherichia coli in individuals who have not taken antibiotics for a long time (Raimondi et al., 2019; Bailey et al., 2020) shows a potential risk for astronauts associated with the possible transmission of AR from normal microflora to pathogenic microflora, as well as the possibility of formation of strains similar to nosocomial ones unprovoked by antibiotics under isolation (Solovieva et al., 2008).
The situation is complicated by the fact that in population studies, the dynamics of an increase in incidence of AR in various microorganisms, and in particular in E. coli, was found (Blanquart, 2019). This means that the risk associated with the increase in multiple antibiotic resistance (MAR) of astronaut microflora and its persistence increases every year. The increase in the risk of MAR can be explained by increasing selective pressure caused by human activity (Peterson and Kaur, 2018). It is important to note that selection for AR occurs even at very low concentrations of certain antibiotics (Klümper et al., 2019).
A model describing the formation of multiple resistance determinants is presented by Lehtinen et al. (2019). This article considers the following factors influencing transmission of MAR: shared resistance mechanisms; genetically determined resistance (associated with particular alleles); genetically determined resistance (associated with the presence of the gene); drug exposure of individual host; resistance status/risk that informs antibiotic choice; and cost epistasis.
In any realization of multiple resistance, it has been found that the emergence of new resistance determinants is most likely in strains with MAR and changes in these factors affecting MAR transmission cannot significantly affect the intensity of transmission (Lehtinen et al., 2019).
However, a recent study (Basra et al., 2018) found evidence of a negative correlation between E. coli growth rate and AR. As the authors note, this relationship is sometimes weak and depends on the environment in which growth rates are observed. This raises the hope that perhaps the impact of space conditions will curb the spread and reduce the stability of AR determinants under manned spaceflight conditions.
The purpose of this work is to build a model that allows us to evaluate the influence of space conditions on the stability of AR determinants.
2. Materials and Methods
Waiver of consent: Elements of consent are eliminated. Data collected on past procedures that are completed.
The analyzed data are presented as results of two space experiments: An experiment to determine the amount of AR determinants in E. coli strains isolated from 16 astronauts (participants of the SALYUT 7 space program) 2 days before and 2 days after the spaceflight. Experiment “Plasmid,” conducted on board the ISS using the equipment “Recomb-K.”
2.1. Determination of AR in E. coli isolated from astronauts
E. coli strains were isolated from 16 astronauts—participants in the SALYUT 7 space program from 1983 to 1986. Fecal samples were taken 2 days before and after the spaceflight.
Sampling was carried out in a sterile container indicating the full name, date, and time of sampling. The sample was not frozen and it was delivered to the laboratory within 2 h.
The study of the qualitative and quantitative composition of the intestinal microflora was carried out in advance using a modified method (Ilyin et al., 2018), which is based on the method by Haenel and Bendig (1975) and modified by Lizko et al. (1979). The essence of the method lies in the dosed inoculation of 10-fold dilutions of precisely weighed material on dense (dish) and semiliquid (test-tube) media. From each sample, five strains of E. coli were isolated.
To determine the level of AR, the method of serial dilutions was used with a medium containing an antibiotic in the following amounts: 4, 16, 32, 64, 128, and 512 μg/mL (standard minimum inhibitory concentration practice). The criterion for the presence of a determinant of AR is the presence of confirmed resistance for a particular antibiotic, which is detected by the method of serial dilutions.
2.2. Space experiment “Plasmid”
The results of the space experiment were compared with ground experiments:
Synchronous experiment
Postflight experiment
Additional information about the experimental conditions can be found in the study by Ilyin et al (2022).
The strains were used in the framework of the Russian–Belgian space experiment “Mobilization,” in which the Catholic University of Louvain and the Belgian Nuclear Research Centre participated, and the strains belong to these institutes (DeBoever et al., 2007).
In the experiments, the equipment, Recomb (Fig. 1), developed by Biotechsys (Russia) was used. The Recomb hybridizer consisted of three series-connected chambers containing a donor, helper, and recipient: the first chamber containing a donor; second chamber containing a helper; and third chamber containing a recipient. Before the start, the chambers were filled with fresh broth cultures of donors, helpers, and recipients.

The Recomb hybridizer consisted of three series-connected chambers containing a donor, helper, and recipient: the first chamber containing a donor (at the top of the hybridizer); second chamber containing a helper (in the central part of the hybridizer); and third chamber containing a recipient (at the bottom of the hybridizer) (DeBoever et al., 2007).
After that the hybridizers were placed in a refrigerated Biokont container and sent into orbit in the Soyuz spacecraft. A day after arrival at the ISS, the equipment was stored at a temperature of +4°C, then 3 days before the end of the flight, donor cultures were mixed with helper cultures, after which the mixture was incubated for a day at a temperature of +37°C. At the end of the incubation, these mixtures were transferred to chamber 3, where they were mixed with the cultures of recipients. These mixtures were incubated at +37°C for a day.
Then, before moving to the Soyuz transport spacecraft, they were kept chilled, and then they were placed in the Soyuz cooled thermocontainer (+4°C) and returned to Earth. In addition, under spaceflight conditions, similar cultures of donors, helpers, and recipients were exposed in separate test tubes equipped with radiation dosimeters to find out how the adaptation of donors, helpers, and recipients to the effects of spaceflight factors can affect the change in the frequency of resistance plasmid mobilization.
In this case, conjugation was carried out under ground conditions (postflight experiment). On the day of landing, the contents of all Recomb chambers were removed and the biomaterial was inoculated on counterselective nutrient media to isolate transconjugants.
Part of the contents of the test tubes was also extracted, and this material was used in studies on mobilization of plasmids at the State Scientific Center of the Russian Federation, IMBP RAS. The rest of the biomaterial was immediately sent to the Belgian Nuclear Research Centre SCK-CEN, the Free University of Brussels, the Catholic University of Louvain, and the University of Ghent, where studies began a day later.
A synchronous experiment was organized as a control, in which all manipulations were carried out synchronously with the flight ones on similar cultures and similar equipment.
A month after the space experiment, a control deferred experiment (postflight experiment) was carried out, in which small temperature and time errors found after the flight were taken into account.
2.3. Experimental cultures
Gram-negative conjugation of three cultures was carried out: helper: E. coli CM 140 (contains nonconjugative plasmid RP4: IncP1 (60 kb); donor: E. coli CM 1962 pMOL222: IncQ (27 kb) with ncc and nre genes; and recipient: Ralstonia metallidurans AE815, plasmid free, RifR.
Fresh broth cultures of donors and recipients at a concentration of 107 CFU/mL each were loaded into chambers 1, 2, and 3 of the Recomb-K equipment (two hybridizers) 18 h before the start. The hybridizers were placed in a refrigerated container.
During the experiment, the crew performed a series of operations that are described in detail by Ilyin et al. (2022). Ground synchronous control was carried out. Postflight studies included the following operations:
Seeding material on counterselective media and counting colonies of transconjugants.
Isolating donor and recipient colonies to study plasmid mobilization after the flight.
Conducting studies on plasmid mobilization after the flight.
Examining the linkage of genes transferred with the studied plasmid.
To do this, three colonies of transconjugants were isolated from each dish and their sensitivity to antibiotics included in the plasmid was studied using the disk method.
The characteristics of the cultures used in the experiments are summarized in Tables 1 and 2. For a number of conjugation pairs, two gene transfer processes were studied—donation (when the helper plasmid is mobilized by the donor plasmid in the donor cell) and conduction (when the helper plasmid is mobilized by the donor plasmid in the cell in which the helper plasmid is deposited).
Experimental Design with Gram-Negative Bacterial Cultures (Strains Were Exposed in Space, but Conjugation Took Place on Earth)
Experimental Design with Gram-Negative Bacterial Strains (Conjugation Took Place in Space)
3. Results
In this article, we consider two experimental approaches: in vivo, an experiment with a bacterium isolated from astronauts, and in vitro, an experiment conducted under controlled conditions. This approach made it possible to determine how strongly cosmic conditions affect the stability of AR determinants in in vivo and in vitro conditions. Obviously, the astronaut's body is an additional factor influencing the stability of AR determinants in bacteria living inside and on the surface of the astronaut's body. An autonomous in vitro experiment made it possible to find out the direct role of cosmic conditions in the formation of the stability of AR determinants.
To successfully solve the problem, it was necessary to create a mathematical model that allows us to use two experimental points (the first point—before the spaceflight, and the second point—after the spaceflight) to assess the stability of AR determinants as integral (in time) values.
3.1. Mathematical model of stability of AR determinants
For the statistical description of the stability of AR determinants, we chose an adapted version of the classical Luria–Delbrück equation, namely a solution based on the Poisson distribution (Haddix et al., 2000). The Poisson distribution, in the study of AR, is widely used in various modifications of the Poisson equation (Shapiro et al., 2020; Megraud et al., 2021; Tartof et al., 2021). However, as the authors note (Card et al., 2021), the Poisson distribution is an imperfect null model in that case because it assumes that mutations occur with uniform probability across all genes.
Below we present our phenomenological model in which this limitation can be lifted. The definition of Poisson distribution describes a discrete distribution of a stream of rare events that have a certain fixed average intensity and do not affect each other. The Poisson distribution can be written as follows:
where
It is logical to assume that in time T, sufficient for implementation of all possible events n, the integral of the probability
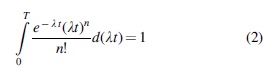
By transforming Formula 2, we can obtain the formula known as the Hermite integral (provided that
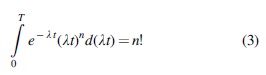
where
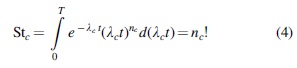
where the subscript c indicates the specified external conditions.
The integrand  in Equation 3 is a function that describes the stability of AR determinants over time (Fig. 2). Function
in Equation 3 is a function that describes the stability of AR determinants over time (Fig. 2). Function 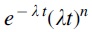 shows the rate of change of stability of AR determinants.
shows the rate of change of stability of AR determinants.

Evolution of the stability of AR determinants across time. An experiment to determine the number of AR determinants in Escherichia coli strains isolated from 16 astronauts (participants of the SALYUT 7 space program): nBefore—2 days before the spaceflight; and nAfter—2 days after the spaceflight. Experiment “Plasmid,” conducted on board the ISS using the equipment “Recomb-K”: n1—experiment in spaceflight conditions; n2—synchronous experiment in terrestrial laboratory conditions—control; n3—delayed experiment in terrestrial laboratory conditions with strains preincubated in space—experiment; and n4—control for the delayed experiment. AR = antibiotic resistance; ISS = International Space Station.
The same function can be obtained by considering two processes that are mutually opposite in time: a power-law growth (the process of cointegration) of AR features, 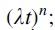 and an exponential process of decay of AR features,
and an exponential process of decay of AR features,
Thus, by describing the phenomenon of the stability of AR determinants by two mutually opposite processes, we obtained a model [functional time dependence—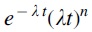 ] of the stability in time of AR determinants. In essence, the integrand describes the evolution of AR over time. First, there is a sharp increase in the number of AR determinants, then a maximum and a gradual decrease in the number of AR determinants.
] of the stability in time of AR determinants. In essence, the integrand describes the evolution of AR over time. First, there is a sharp increase in the number of AR determinants, then a maximum and a gradual decrease in the number of AR determinants.
By integrating this function (calculating Eq. 3), we obtain a value (the area of the integrand) reflecting the stability of the AR determinants. By analyzing the obtained result, we arrive at a method for comparing the stability of AR determinants under various conditions.
Therefore, if we have a number of AR determinants under terrestrial conditions equal to nE
, and under space conditions in the same bacteria equal to nS
, then the stability ratio of AR determinants will be expressed by the following formula:
Formula 4 is predictive in nature since it takes into account significant periods of rise and subsequent decrease in the stability function of the AR determinants.
In addition, Formula 5 makes it possible to make prognostic estimates of the difference in the stability of AR determinants under different conditions.
3.2. The result of the experiment to determine the number of AR determinants in E. coli isolated from astronauts
Of the 80 E. coli cultures isolated before the flight, 30 cultures with 2 AR determinants were found. After the spaceflight, the number of cultures with 2 AR determinants decreased to 12 (Table 3). After the spaceflight, the number of cultures that do not have AR determinants increased from 30 to 64. Cultures with three or more AR determinants have undergone even greater changes after the spaceflight.
Changes in Sensitivity to Antibiotics of Escherichia coli Isolated from Astronauts 2 Days Before and After a Spaceflight
AR = antibiotic resistance.
In general, the average number of AR determinants before the flight was 1.325 per culture, and after the spaceflight, this value decreased to 0.400. We tested the statistical significance of the difference between the two groups (preflight and postflight) using Fisher's exact test (p < 0.001).
Using Formula 5, we can evaluate the influence of cosmic conditions on the stability of the AR determinants in E. coli cultures. Thus, the integral value of the stability function of the AR determinants in E. coli cultures before the flight was 
This suggests that before the spaceflight, the stability of AR determinants in E. coli cultures was 33.5% higher than after the spaceflight. Thus, space conditions reduce the stability of E. coli AR determinants in astronauts' bodies by 33% relative to terrestrial conditions. The reason for this may be additional influences on the part of the body of astronauts—their immunity, physiological changes, diet, work, and rest (Crucian et al., 2018).
A more recent experiment carried out in vitro on the ISS made it possible to directly assess the stability of AR determinants in gram-negative bacteria.
3.3. The result of the in vitro experiment
Of scientific interest is the question of the stability of AR determinants preincubated in space conditions and developing in terrestrial conditions. This question and the question posed above are answered in Table 4. By analogy with the previous calculation, we calculate the changes in the stability of AR determinants under the conditions of the in vitro experiment. In this case, Formula 5 will use the following values:
Change in Sensitivity to Antibiotics of Gram-Negative Bacteria: an In Vitro Experiment Carried Out on Board the International Space Station and in Terrestrial Conditions
We get the following ratio: 
According to the results of the in vitro experiment and using Formula 5, it was found that in space conditions, the stability of AR determinants for gram-negative bacteria increased by 33%. In space-preincubated strains of gram-negative bacteria that were then cultured on Earth, the stability of AR determinants decreased by 8% relative to the control.
4. Discussion
The mathematical model we developed allows us to make predictions of the integral value of the stability of AR determinants. This takes into account the time dependence of the stability of AR determinants with a characteristic period of an increase in AR intensity and its subsequent decrease. This model made it possible to evaluate the stability of AR determinants both in the in vivo experiment and in the in vitro experiment. In the in vivo experiment, we found a decrease in stability, by 1/3, under spaceflight conditions relative to the preflight level, and in the in vitro experiment, we found an increase in stability in gram-negative bacteria, by 1/3, relative to the control under terrestrial conditions.
Apparently, this difference is caused by the indirect action of the processes of adaptation of the astronauts' immunity to space conditions. Attention is drawn to the fact that in both experimental conditions (in vivo and in vitro), we observe the same change in stability of AR determinants—by 1/3, but with a different direction. We can assume that if this is not a coincidence, then perhaps this is due to activation or deactivation of one of the mechanisms for stabilizing AR determinants in bacteria.
Differences in AR plasmid stability could be introduced by varying environmental conditions, such as intestinal surface with immune cells, as opposed to cytokine-free broth cultures. Decreased AR plasmid stability in astronauts may be linked to the higher growth rate of plasmid-free bacterial cells. Considering that AR genes are usually found on larger plasmids in E. coli (Wein et al., 2020), the bigger plasmids lead to higher fitness cost and reduced segregation stability (Wójcik et al., 1993).
The increase in stability of AR determinants in in vitro cultures may be due to a sufficient AR plasmid copy number for effective segregation between daughter cells. Additional aspects that could determine the direction of AR plasmid stability are sequences of plasmids, particularly the origin of replication genes, mechanisms of replication, and capability of recombination of the plasmid and the particular host strain (Friehs, 2004).
Comparison of data on the stability of resistance determinants in the in vitro experiment and data on AR in a real flight indicates the correctness of the chosen in vitro experiment and explains the reason for these changes. After all, in a spaceflight (in vivo experiment), the number of microorganisms resistant to two antibiotics was the majority (Table 3), even despite the total decrease in multidrug resistance, presumably due to the predominance of AR plasmid segregation processes over cointegration processes.
It can be assumed that biresistant cultures in a real flight are products of segregation of plasmids containing a greater number of resistance determinants. Meanwhile, the number of monoresistant cultures, which can also be products of such segregation, is relatively small compared with biresistant cultures. Moreover, in the in vitro experiment, an increased tendency toward the formation of precisely biresistant clones was demonstrated (Table 4). If biresistant determinants are stable in bacterial genomes, it can be assumed that they will exist in human microflora populations for a long time in the spaceflight and can create problems in choosing an antibiotic therapy strategy if indicated.
5. Conclusions
Long-term spaceflights, missions to the Moon and Mars, are associated with significant risks such as the reaction of the astronaut's microflora to space conditions (Tesei et al., 2022). The question of the stability of AR determinants under spaceflight conditions remains open. In this article, we have proposed a model that allows us to compare the stability of AR determinants in spaceflight conditions and in laboratory conditions on Earth.
The presented model showed changes of the same magnitude (33.5%), but opposite in direction, of the AR determinants' stability of bacteria isolated from astronauts and bacteria under the conditions of the in vitro experiment. One can only suggest that the similarity of stability values, 1/3, in different experiments indicates that there is a mechanism maintaining the stability of AR determinants, which is turned on or off depending on the conditions (in vitro or in vivo).
An important fact was also discovered—the predominance of biresistant forms of bacteria in space conditions. This fact imposes certain limits in the formation of a strategy for antibacterial treatment in space conditions.
Footnotes
Acknowledgments
This work was performed in the framework of a research program of the Institute for Biomedical Problems, Russian Academy of Sciences 64.2.
Authors' Contributions
V.I. and O.O. supervised the study and provided financial support. V.I. and A.A. contributed to conception of the study. A.A., M.S., and A.K. carried out the computational work. A.A., E.P., and A.K. performed the calculations and statistical analysis. V.I., E.P., D.M., M.S., and A.A. wrote the draft of the manuscript. All authors took part in revision and approved the manuscript.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The authors did not receive financial support from any organization for the submitted work.
Abbreviations Used
Associate Editor: Don Cowan