
Abstract
Barkol Lake, situated northeast of the Tianshan Mountains, Xinjiang, is a hypersaline lake with abundant sulfate and chloride minerals, which can be a potential analog for microbial saline paleolakes on Mars. The lake water, sediments, and surrounding soils of Barkol Lake were sampled for geochemical analysis and 16S rRNA gene sequencing to investigate the prokaryotic community structure, abundances, interactions, and ecological functions. Results show that (1) prokaryotic community structure differs significantly between biotopes (water, sediment, and soil), with the highest abundances of archaea occurring in water samples and highest prokaryotic diversities in soil samples; (2) archaeal communities are dominated by Halobacterota, Nanoarchaeota, Thermoplasmatota, and Crenarchaeota, while bacterial communities are mainly Proteobacteria, Bacteroidetes, Actinobacteria, Desulfobacterota, Chloroflexi, Gemmatimonadetes, Firmicutes, and Cyanobacteria; (3) the prokaryotic community network for soil is far more complicated and stable than those for water and sediment; (4) soil prokaryotic communities could be significantly affected by environmental factors such as salinity, pH, total sulfur, and Ca2+; (5) archaeal communities may play an important role in the nitrogen cycle, while bacterial communities may mainly participate in the sulfur cycle. This study extends the data set of prokaryotic communities for hypersaline environments, which will provide perspectives into identification of the counterparts and help to understand potential microbial interactions and biogeochemical cycles occurring on Mars.
Introduction
Salt lakes are a kind of commonly found saline or hypersaline environment in semi-arid or arid regions, and the microbes that inhabit them have attracted increasing research interests (Jellison, 2005; Oren, 2014). Halophiles are a group of halophilic and halotolerant microbes that grow and thrive in extreme environments of high salt concentrations, primarily prokaryotes (archaea and bacteria) (Gupta et al., 2014). Salt lakes on Earth have been found to support a surprisingly high diversity of halophiles, such as Halobacteriales (archaea) and Salinibacter (bacterium) (Litchfield, 2002; Oren, 2014), forming unique ecosystems (Tazi et al., 2014; Zaikova et al., 2018). The halophiles in salt lakes may not only contribute to biotechnological and biomedical industries (Zheng et al., 2016) but also inform the search of life on Mars (Cavicchioli, 2002). The current climate of Mars is characterized by extreme aridity and cold temperatures (current average ∼ -60°C), and there are widespread salt deposits of chloride, sulfate, and perchlorate minerals that could be remnants of paleolakes on the martian surface (Squyres et al., 2004; Vaniman et al., 2004; Wordsworth et al., 2017). Chloride minerals can substantially lower the freezing point of water (up to -50°C) (Brass, 1980; Buffo et al., 2022) and reduce water sublimation (Aerts et al., 2019) such that the existence of liquid water is possible on Mars, which is essential to sustain life. Microbes typically account for the oxidation of sulfides to sulfates in terrestrial acid mine drainage sites (Colmer and Hinkle, 1947), which could also explain the origins of sulfate minerals on Mars (Bishop et al., 2005). In particular, sulfate minerals, such as jarosite (KFe3(SO4)2(OH)6), can act as a shelter that preserves amino acids from UV radiation and cold temperature conditions (Martinez-Frias et al., 2006; Santos et al., 2016), while gypsum (CaSO4·2H2O) and the commonly associated halite (NaCl) minerals can serve as a geological niche for microbes by providing significant moisture with fluid inclusions and favoring photosynthesis (Parnell et al., 2004; Edwards et al., 2005; Osterloo et al., 2010). In addition, hypersaline environments can potentially inhibit the decay of organic matter and preserve biological materials over geologically significant timescales (Martinez-Frias et al., 2006). These factors all suggest that paleolakes abundant in chloride and sulfate minerals would be relatively good candidates for the discovery of evidence of life on Mars. The halophilic microbiology in similar salt lakes on Earth could provide important insight into forms of existing or past life that might have occurred on Mars.
Despite fast-moving progress in cultivation-independent molecular biological techniques (Madigan and Oren, 1999), microbiological research of salt lakes is relatively rare compared to that of other terrestrial ecosystems. Even though China is one of a number of countries in the world with an abundance of salt lakes (Zheng et al., 2016), current studies of salt lakes in China are mainly focused on the isolation of halophilic microbes and less so on the prokaryotic community structure, abundances, and ecological functions (e.g., Tao et al., 2020; Yang et al., 2021). The distribution pattern and controlling mechanisms of halophilic microbiology have not been well understood (Gupta et al., 2014). Therefore, it is imperative to advance the comprehensive understanding of halophilic microbiology in salt lakes, which could be pivotal to understanding the evolution and sustainability of extreme life.
Barkol Lake, Xinjiang, is a shrinking hypersaline lake with a cold and dry climate as well as abundant mirabilite (Na2SO4·10H2O), halite, gypsum, and magnesium sulfate (MgSO4·nH2O) minerals (Wei et al., 1998). Barkol Lake can be a potential analog for saline paleolakes on Mars, and its prokaryotic diversity may be of relevance to the search for life on Mars. Since 1998, a large number of halophilic organisms (bacteria, algae, protozoa, brine shrimp, etc.) and their fossils have been found in mirabilite deposits of Barkol Lake (Wei et al., 1998). At present, some Chinese studies with only local importance have explored the prokaryotic communities in Barkol Lake to show high prokaryotic diversities of this extreme environment (e.g., Gu et al., 2007; Xiang et al., 2018; Yang et al., 2019). However, there are few studies that have comprehensively explored the archaeal and bacterial diversities in Barkol Lake and its surroundings. Further, its network stability and ecological functions of the prokaryotic communities have not been investigated. In this study, the structure, abundances, interactions, and ecological functions of the prokaryotic communities in Barkol Lake are discussed based on 16S rRNA gene sequencing, and implications of this study for astrobiological exploration are highlighted as well.
Materials and Methods
Study area and sampling
Barkol Lake (43.61°N, 92.82°E, 1580 m a.s.l.) is situated northeast of the Xinjiang Tianshan Mountains and is a shrinking sulfate-chloride-type hypersaline lake with periodic fluctuations of water surface (40–100 km2) and water depth of 0.7–0.9 m (Lu et al., 2012) (Fig. 1). It is located in the continental arid and semi-arid climate zone, with a mean annual temperature of 1.1°C (extreme minimum: -43.6°C), annual precipitation of 210 mm, and potential evaporation of 2000–2500 mm (Ma et al., 2004; Wei and Wang, 2018).

Map and satellite imagery (
Lake water, sediment, and soil samples from across the depth profile and horizontal transect in the desiccated surroundings were collected in July 2018 (Table 1). Lake water samples were collected from various locations of the lake surface (<1 m depth) and were then thoroughly mixed and sequentially filtered through 120 μm (W120μm), 3 μm (W3μm), 0.45 μm (W0.45μm), and 0.22 μm (W0.22μm) diameter membrane filters. After each stage of filtration, membrane filters of the same porosity size were pooled to collect the retained microbes. Sediment samples were collected 20 m (S20m), 60 m (S60m), and 100 m (S100m) away from the shoreline toward the center of the lake. Transect topsoil samples (0–2 cm depth) were collected 5 m (barren surface, T5m), 10 m (near absence of plants, T10m; ∼20% plant coverage, T10mA), 20 m (∼20% plant coverage, T20m), 30 m (∼50% plant coverage, T30m), and 40 m (100% plant coverage, T40m) away from the shoreline (Fig. 1). Profile soil samples were collected at depths of 10 cm (P10cm), 20 cm (P20cm), 30 cm (P30cm), 40 cm (P40cm), and 50 cm (P50cm) in a soil pit that was 20 m away from the shoreline. Three replicates were collected for each sample. All the samples were frozen, transported on ice to the laboratory, and stored at -80°C until analysis.
Summary Statistics of 16S rRNA Gene Amplicon Sequencing
T10mA was sampled under plant coverage that differs from T10m sampled where there is a near absence of plants.
Total DNA of each membrane filter or sediment/soil sample was extracted by using the DNeasy PowerSoil DNA Isolation Kit (MOBIO Laboratories, Carlsbad, CA, USA) according to the manufacturers' instructions. The DNA concentration and purity were then detected with the NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Landsmeer, the Netherlands).
For the PCR amplification, the 515f-806r bacterial/archaeal primer pair F:(5'-GTGYCAGCMGCCGCGGTAA-3') and R:(5'-GGACTACNVGGGTWTCTAAT-3') (Caporaso et al., 2012) were used to amplify the hypervariable V4 region of archaeal and bacterial 16S rRNA gene. Reactions were performed in a 50 μL mixture containing 25 μL Taq DNA MasterMix (TIANGEN Biotech Co. Ltd., Beijing, China), 1 μL 20 ng/μL DNA, 1 μL forward primers, 1 μL reverse primers, and 22 μL RNA-free water. PCR program was set as follows: preheating at 95°C for 5 min; then 28 cycles of 94°C for 30 s, 55°C for 30 s, and 72°C for 1 min; the final extension at 72°C for 5 min. The PCR products were purified with the MiNiBEST DNA Fragment Purification Kit Ver. 4.0 (TaKaRa Co. Ltd, Tokoyo, Japan) and then quality checked through gel electrophoresis. Finally, the purified PCR products were pooled for 250 bp paired-end high-throughput sequencing on an Illumina NovaSeq 6000 platform at the Meige Gene Technology Co., Ltd (Shenzhen, China).
Physiochemical properties analysis
Water samples were directly measured by a PHS-3C pH meter (Shanghai INESA Scientific Instrument Co., Ltd., Shanghai, China) for pH and HQ14d digital conductivity and by salinity meter (Hach Company, Loveland, CO, USA) for salinity, while a split of the soil sample was mixed with Millipore water (>18 MΩ·cm) in the soil to water mass ratio of 1:2.5 and 1:5 for the pH and salinity measurements, respectively. The water content of each soil sample was measured as the weight loss during the drying process of the fresh soil. Total organic carbon (TOC), total nitrogen (TN), and total sulfur (TS) in soil samples were measured by EL cube EA (Elementar Analysensysteme GmbH, Hanau, Germany). Meanwhile, soil samples were extracted by water, and the extracts were then filtered through 0.22 μm membrane filters and analyzed for cation (K+, Na+, Mg2+, and Ca2+) and anion (Cl-, NO3 -, and SO4 2-) concentrations by DX-120 ion chromatography (Dionex Corp., Sunnyvale, CA, USA). Water samples were also measured for anion (Cl- and SO4 2-) concentrations by ion chromatography.
Bioinformatic analysis
With the Deblur method in the Quantitative Insights into Microbial Ecology Version 2 (QIIME2), quality control and filtering were first performed on the obtained sequences to remove low-quality sequences. Operational taxonomic units (OTUs) were clustered to represent sequences with 100% similarity. Taxonomic assignments were then annotated with GTDB databases (version r95) for archaea and bacteria. Based on the taxonomic information of the samples, the alpha diversity analysis was performed by using QIIME2 to obtain Chao1, Shannon and Good's coverage indices.
Principal component analysis (PCA) based clustering was then carried out on OTU level by implementing the built-in function prcomp in R (version 3.6.3) to determine the similarities in prokaryotic community structure between samples. Linear Discriminant Analysis (LDA) Effect Size (LEfSe) analysis was performed by using the Biozeron Cloud Platform (
Results
The prokaryotic community structure and abundances
There are 58,237 to 93,345 high-quality archaeal and bacterial 16S rRNA gene sequences for each sample, and each sample possesses 795–4227 OTUs (100% similarity level) (Table 1). The highest Shannon (prokaryotic diversity) and Chao1 (prokaryotic richness) indices occur in the sample T40m, while the lowest prokaryotic diversity and richness occur in the sample W0.22μm. The prokaryotic diversity and richness of sediment or soil samples are both higher than those of water samples. Good's coverage ranges from 0.9978 to 0.9997, which indicates that the archaeal and bacterial 16S rRNA gene sequences retrieved in this study represent the majority of prokaryotic communities accessible via the 515f-806r primers used (Table 1).
On the domain level, the relative abundances of archaea are considerably higher in water and sediment samples than in soil samples, and archaea preferably distribute on fine aggregates in water samples (from 19.44% for the sample W120μm to 68.46% for the sample W0.22μm) (Fig. 2). On the phylum level, there are a total of 75 phyla in Barkol Lake, and the structure of bacterial communities is more complicated compared to that of archaeal communities. The dominant archaeal phyla (defined as the phyla with average relative abundances >1%) are Halobacterota for water samples; Nanoarchaeota, Halobacterota, Thermoplasmatota, and Crenarchaeota for sediment and soil samples (Fig. 3A). All samples possess eight dominant bacterial phyla: Proteobacteria, Bacteroidetes, Actinobacteria, Desulfobacterota, Chloroflexi, Gemmatimonadetes, Firmicutes, and Cyanobacteria (Fig. 3B). The dominant archaeal class in water samples is Halobacteria (Halobacterota phylum) with the relative abundances higher than 98%, while the dominant archaeal classes in sediment or soil samples are Nanoarchaeia (Nanoarchaeota phylum), Halobacteria and Thermoplasmata (Thermoplasmatota phylum), as well as Nitrososphaeria (Crenarchaeota phylum) (Fig. 3C). The dominant bacterial classes in all samples are Gammaproteobacteria and Alphaproteobacteria (Proteobacteria phylum), Bacteroidia (Bacteroidete phylum), as well as Actinobacteria and Acidimicrobiia (Actinobacteria phylum) (Fig. 3D). However, we acknowledge that, due to the biases of 515f-806r bacterial/archaeal primer pair, the abundances of Crenarchaeota phylum and SAR11 order (Alphaproteobacteria class within Proteobacteria phylum) could be underestimated to some extent in this study (Walters et al., 2016).
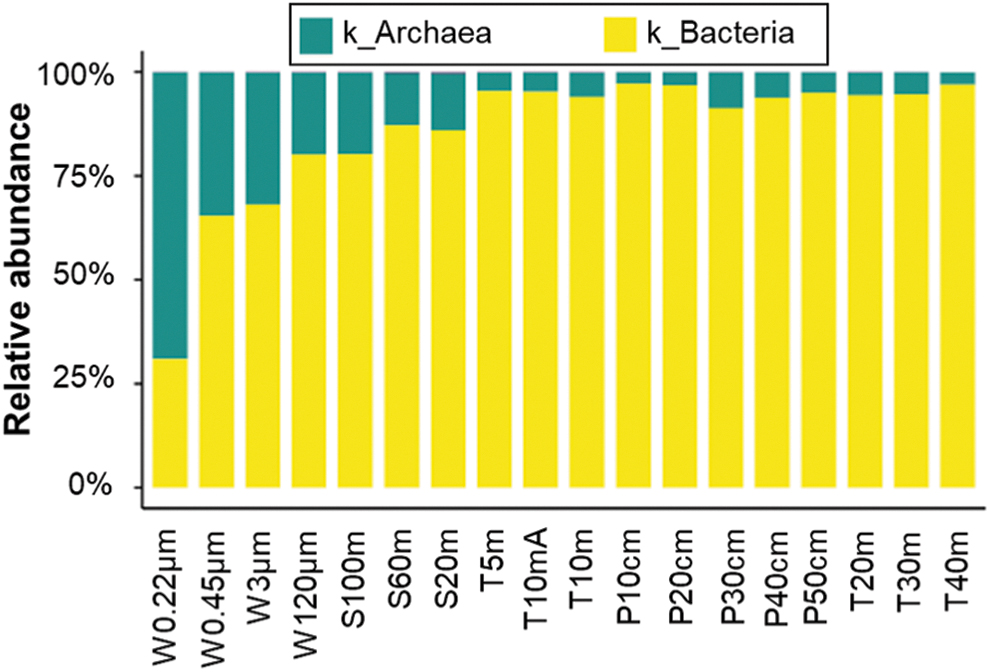
Relative abundances of prokaryotic communities on the domain level. Color images are available online.
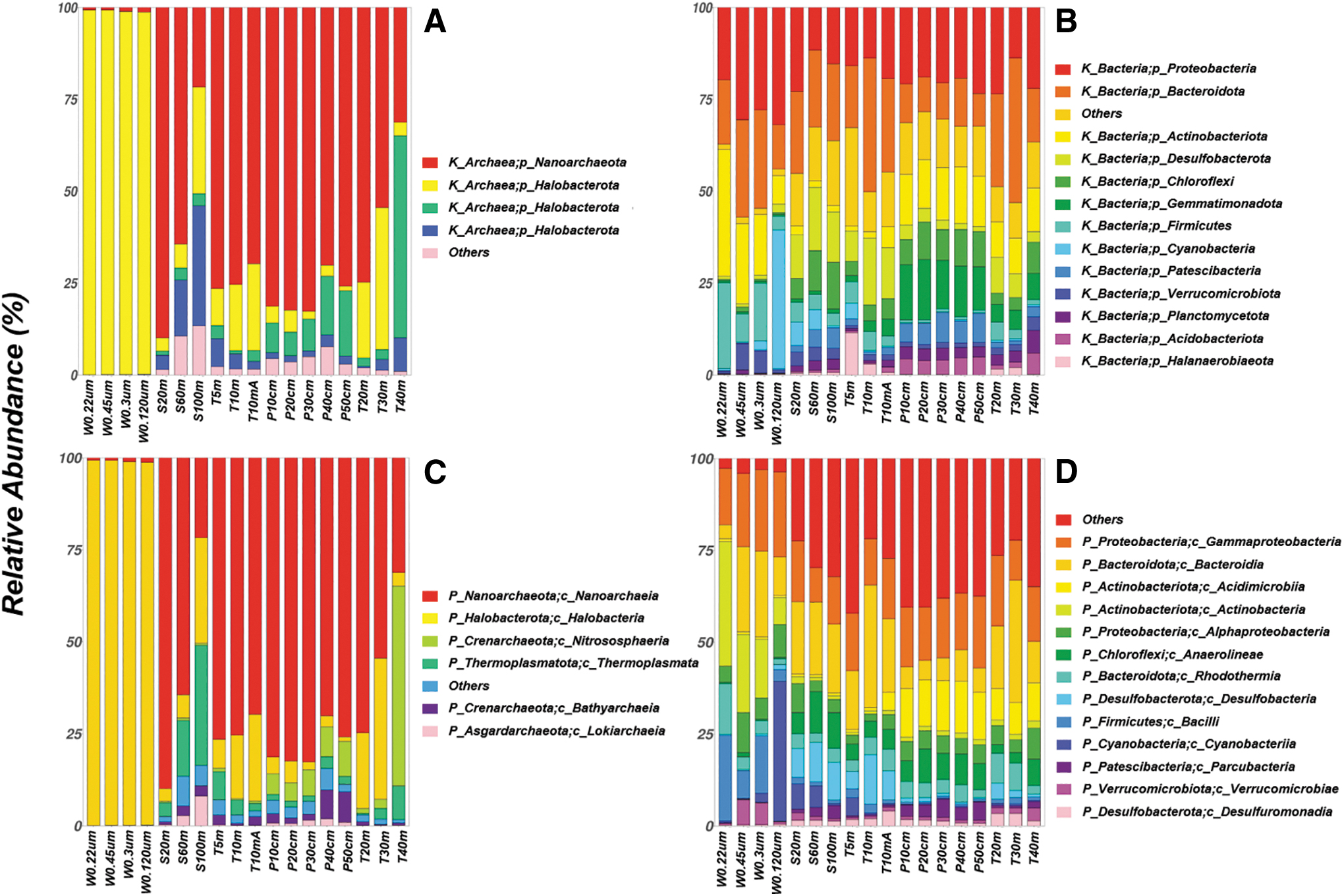
Relative abundances of archaeal communities on the phylum (
Principal component analysis results indicate that samples of the same biotope are similar in both archaeal and bacterial communities on the OTU level (Fig. 4). The total variance (55%) of archaeal communities can be explained by the principal component 1 (PC1, 34%) and principal component 2 (PC2, 21%), while the total variance (60%) of bacterial communities can be explained by the principal component 1 (PC1, 37%) and principal component 2 (PC2, 23%). Water samples are separated away from the other samples. Sediment samples form a denser cluster and are close to the near-shore soil samples (T5m and T10m), while the other soil samples spread significantly.
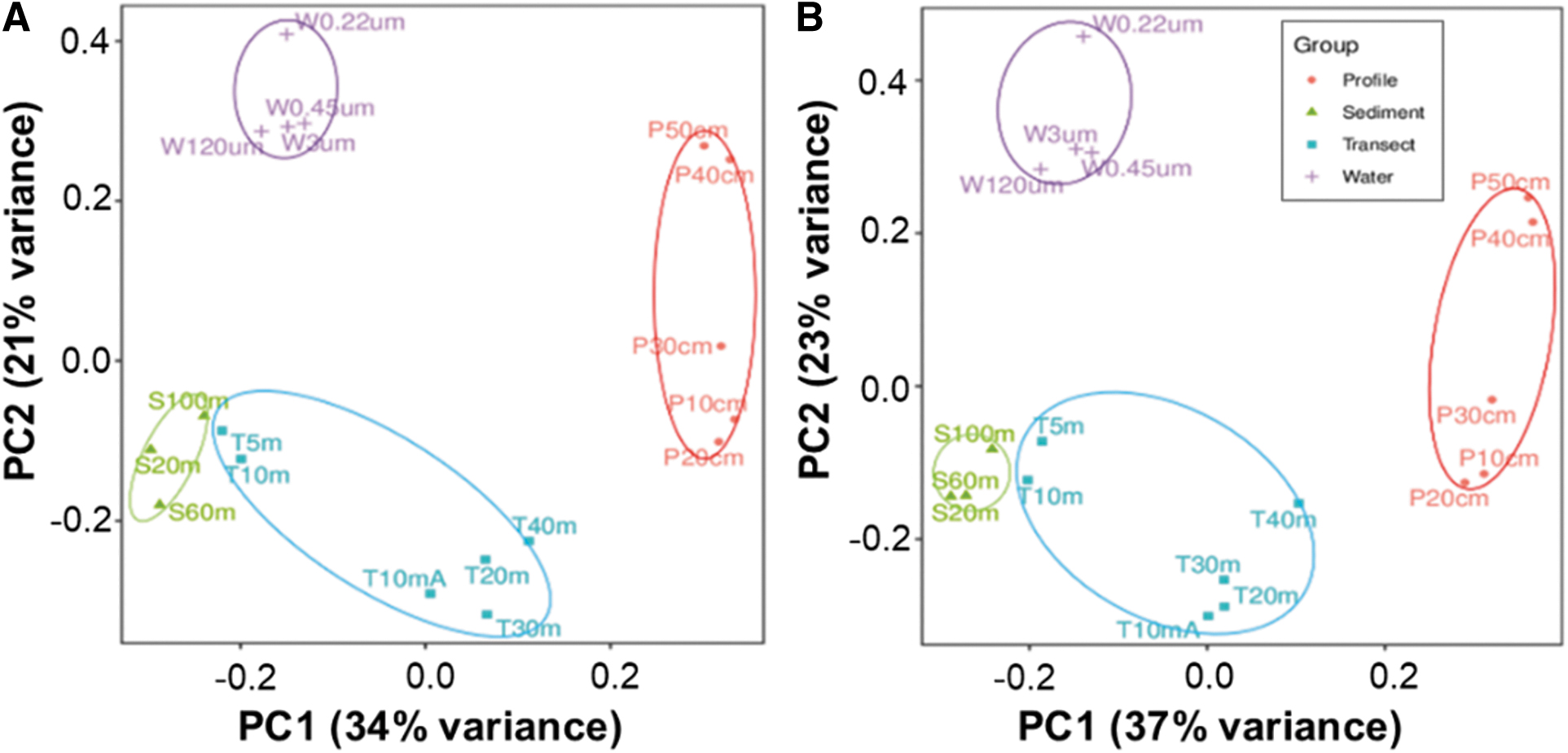
PCA-based clustering on OTU level for archaeal (
LEfSe results indicate that there are 37 taxa groups in archaeal communities and 74 taxa groups in bacterial communities of statistically and biologically consistent differences (LDA score (log 10) > 6) (Fig. 5). Most of them are significantly enriched in water samples, while only one archaeal phylum and three bacterial phyla occur in sediment or soil samples. Specifically, the phyla accounting for the greatest differences between archaeal communities are as follows: Halobacterota (in water) and Thermoplasmata (in sediment and soil) (Fig. 5A). The phyla accounting for the greatest differences between bacterial communities, however, are as follows: Proteobacteria, Bacteroidetes, Actinobacteria, Firmicutes, and Cyanobacteria (in water); Chloroflexi (in sediment); and Gemmatimonadota and Desulfobacterota (in soil) (Fig. 5B). These phyla are specialized and could play important roles in the specific biotopes.
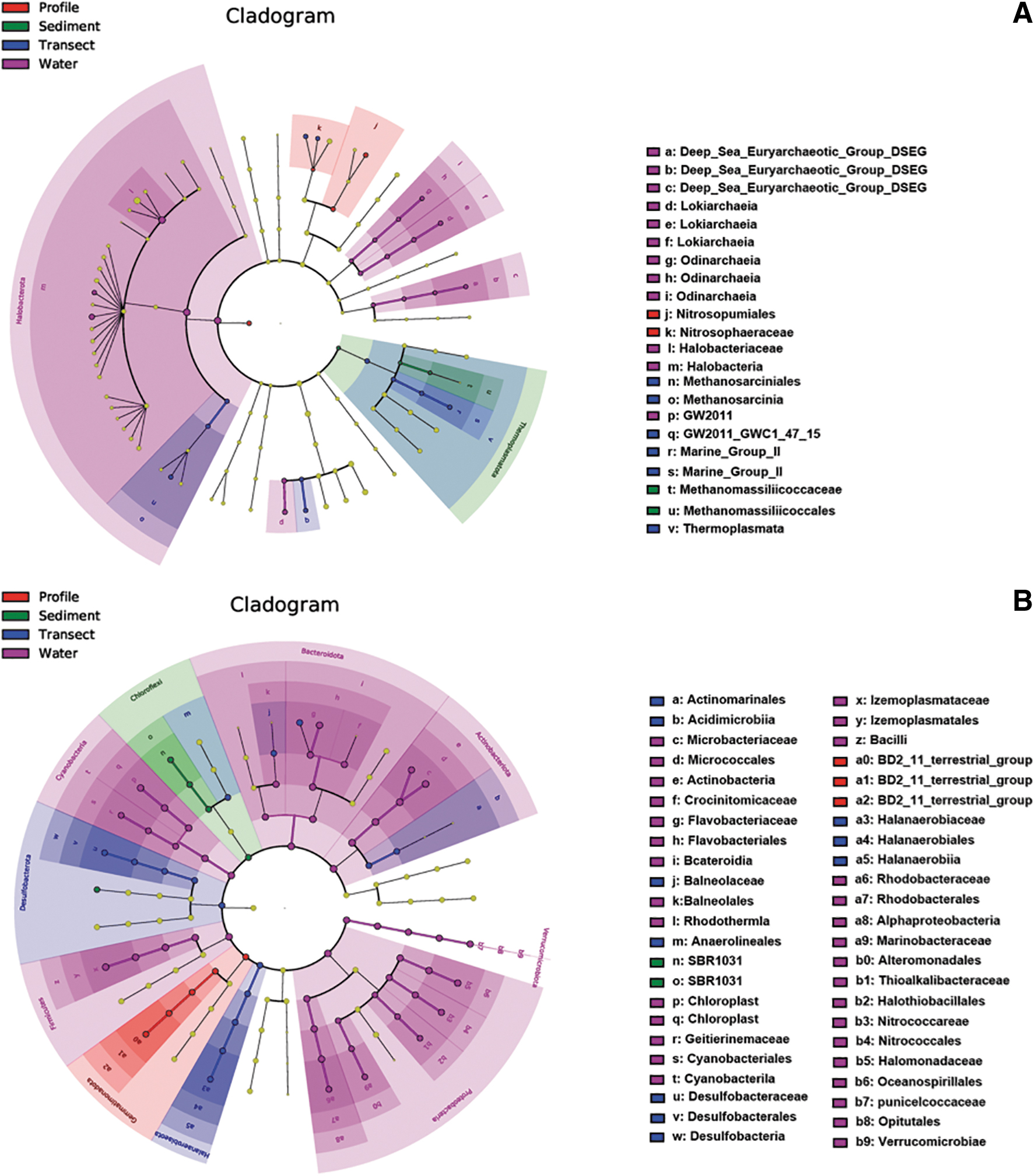
The cladograms of LDA scores based on LEfSe for archaeal (
The constructed networks of prokaryotic community for different biotopes and their calculated topological properties are shown in Fig. 6 and Table 3, respectively. There are 140 nodes (genus with relative abundance >0.2%) in total selected for network construction. Modularity values are all greater than 0.4, indicating the modular structure for each biotope (Banerjee et al., 2016). There are great differences in networks between biotopes. The total nodes (136), total links (4664), average degree (68.588), and average clustering coefficient (0.997) of the soil network are consistently higher than those of the water or sediment networks. Instead, the average path lengths of the water and sediment networks are higher than that of the soil network. The positive correlations (synergistic relationship) between nodes are greater than the negative correlations (antagonistic relationship) in all networks, which indicates that the symbiotic relationship indexes of the three networks are all higher than 50%, in the order of water (75.2%) > sediment (58.8%) > soil (57.8%). Robustness index is 0.43 for the soil network, 0.37 for the sediment network, and 0.33 for the water network, which indicates the order of the network stability (Table 3).
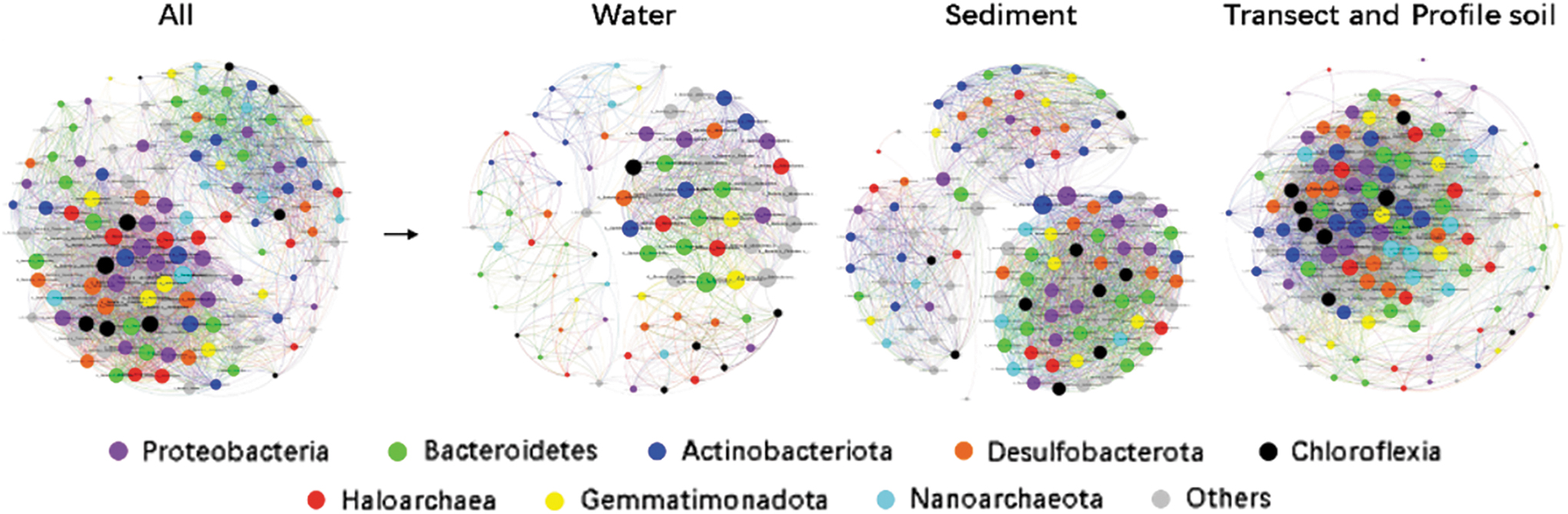
Network analysis for archaeal and bacterial communities of different sample types. Color images are available online.
The water samples have an average pH value of 8.25 and salinity of 77.4‰, and are abundant with SO4 2- (average: 57.7–90.6 g/L) and Cl- (average: 14.5–26.9 g/L). The transect and profile soil samples had pH values ranging from 7.7 to 8.1 and salinities ranging from 3.38‰ to 9.68‰, and TOC and TS contents ranging from 1.77% to 3.32% and 0.34% to 1.13%, respectively, and the major cation and anion Na+ and SO4 2-, respectively (Table 2). For archaeal communities, the first two RDA dimensions explain 63.4% and 27.1% of the total variance, respectively (Fig. 7A). There are strong negative correlations between TS and Halobacterota phylum (p < 0.05, r = -0.70), as well as between pH and Crenarchaeota phylum (p < 0.05, r = -0.76). For bacterial communities, the first two RDA dimensions explained 74.8% and 17.1% of the total variance, respectively (Fig. 7B). Salinity poses the greatest influence on bacterial communities and is significantly correlated with the phyla of Actinobacteriota (p < 0.001, r = -0.91), Desulfobacterota (p < 0.01, r = 0.81), Chloroflexi (p < 0.05, r = -0.67), and Gemmatimonadota (p < 0.01, r = -0.81). TS is significantly correlated with Bacteroidota phylum (p < 0.05, r = -0.73), while Ca2+ is significantly correlated with Gemmatimonadota phylum (p < 0.05, r = 0.67).

RDA ordination plots of the relationships between environmental factors and archaeal (
Physical and Chemical Properties of Profile and Transect Soil Samples
Topological Properties of the Microbial Networks
For a genus.
The correlation between nodes.
The average number of connecting edges for each node.
The degree that nodes tend to gather together.
The average network distance of all nodes.
For the network diagram structure.
Measured as the proportion of positive correlation links to total links.
Measured as the proportion of nodes remained with 50% nodes randomly removed in each network.
Based on the FAPROTAX database, the ecological functions performed by archaeal communities are relatively simple compared to those performed by bacterial communities. A large number of archaeal communities are predicted to operate chemoheterotrophy (averaging 36.6%), and most of the chemoheterotrophs are aerobic (averaging 34.9%) (Fig. 8A). Some archaeal communities are predicted to be related to the nitrogen cycle, mainly nitrification, while others are involved in methanogenesis. Meanwhile, a significant number of bacterial communities are also predicted to operate aerobic chemoheterotrophy (15.2–34.2%) (Fig. 8B). Some bacterial communities are assigned to be predatory or exoparasitic (especially in water samples W0.45μm and W0.3μm) and photoautotrophs. Instead, unlike archaeal communities, bacterial communities are predicted to play an important role in the sulfur cycle, mainly dark oxidation (oxidation under dark conditions) of sulfur compounds (i.e., sulfur, sulfide, and thiosulfate) and sulfate respiration.
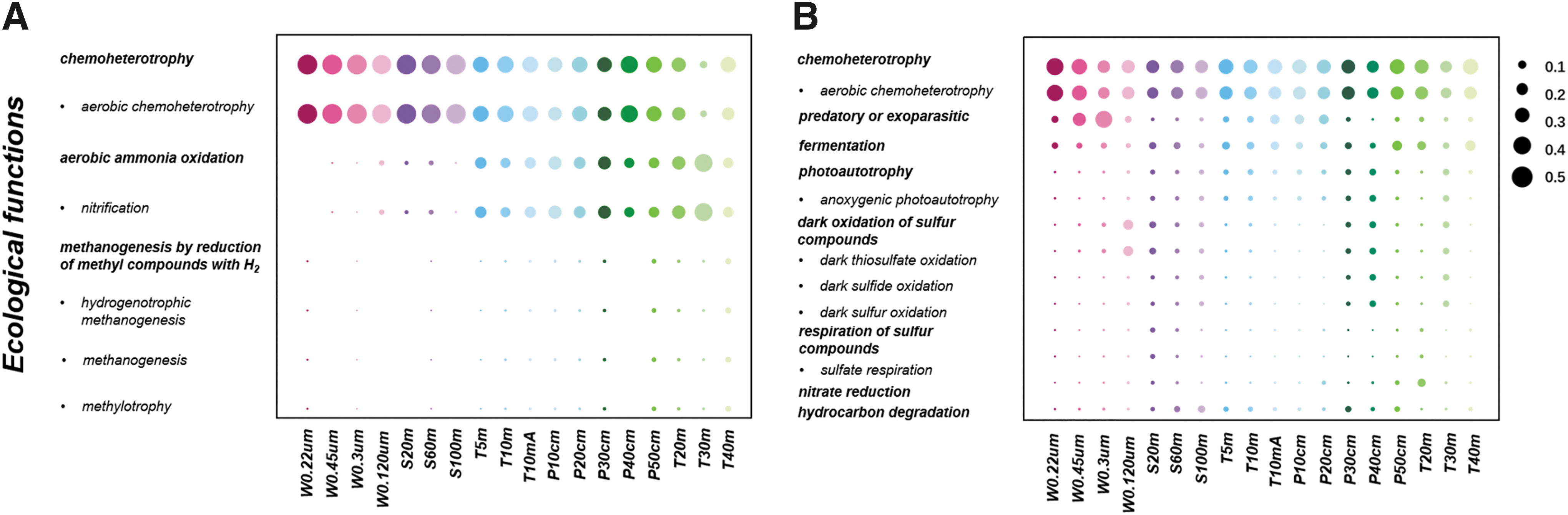
The predicted ecological functions of archaeal (
The structure of prokaryotic communities
The prokaryotic community structures in Barkol Lake differ between biotopes (water, sediment, and soil) and show similarities to those of other salt lakes (Pontefract et al., 2017; Aerts et al., 2019). In detail, the structure of prokaryotic communities in water samples is radically different from those in sediment, transect soil, and profile soil samples, with much higher abundances of archaea in water samples and higher prokaryotic diversities in soil samples (Fig. 3). Archaeal communities are more important in water samples, which is in line with the gradual dominance of archaeal communities in lake water with the increasing salinity observed in the Tibetan Plateau (Zhong et al., 2016). In addition, archaeal communities could be relatively small in size, considering their high abundances on filter membranes between 0.22 and 0.45 μm. The predominant and specialized archaeal phylum in water samples is Halobacterota (98.6–99.2%), specifically Halobacteria class, a well-known group of microorganisms that live in extremely saline or hypersaline environments (Oren, 2014). In sediment and soil samples (with salinities of 3.38–9.68‰), the archaeal phylum Nanoarchaeota accounts for the majority of the whole archaeal communities except in S100m (near the center of the lake) and T40m (farthest from the shoreline), while the archaeal phyla Thermoplasmata (specialized) and Crenarchaeota are also important. Nanoarchaeota phylum has been detected in marine hydrothermal sediments, terrestrial hot springs and hypersaline environments, but little has been known about it so far (Jarett et al., 2018). Thermoplasmata phylum has been widely identified, and some can survive in extreme environments (Golyshina et al., 2016; Y. Yuan et al., 2021). Nitrososphaeria class, the dominant class within Crenarchaeota phylum in Barkol Lake, is widespread across a great variety of ecosystems (e.g., oceans, soils, and freshwater) and could play a significant role in the nitrogen cycle (mainly nitrification) (Ren and Wang, 2022).
The structures of bacterial communities in Barkol Lake are more complicated. The water samples have specialized bacterial phyla of Proteobacteria (19.6–31.8%), Bacteroidetes (12.0–26.2%), Actinobacteria (7.8–34.5%), and Firmicutes (3.8–22.3%). Proteobacteria phylum is the largest phylum in the domain of Bacteria, which usually play a variety of roles in nitrogen fixation, nitrogen removal, desulfurization, organic matter degradation, and so on (Philippot, 2005; Yamamoto and Takai, 2011); and Gammaproteobacteria and Alphaproteobacteria classes are the two major classes in Barkol Lake. Bacteroidetes phylum is abundant in various environments such as marine surface waters (Munoz et al., 2020). The relative abundances of Actinobacteria phylum (mainly Actinobacteria class) in water samples decrease as the porosity size of filter membrane gets larger, which suggests its small particle size in contrast to that of Cyanobacteria phylum (mainly Chloroplast class), which is mainly distributed on the sample W120μm (33.93%). Candidatus Aquiluna, the dominated genus within Actinobacteria class and Actinobacteria phylum, is a group of selenoid-shaped photoheterotrophic microorganisms previously identified in various brackish to saltern waters (Kang et al., 2012; Leoni et al., 2020). Firmicutes phylum is widespread in nature, and many were found to prevail in highly saline sites, such as Bacilli class, which dominates in Barkol Lake (Panosyan et al., 2018).
The sediment and soil samples have the specialized bacterial phyla of Desulfobacterota (1.3–13.9%), Chloroflexi (3.0–12.7%), and Gemmatimonadota (0.9–15.0%). Desulfobacterota phylum has been identified in environments that span wide ranges of salinity, pH, and temperature and could get involved in the sulfur cycle (Murphy et al., 2021). Chloroflexi phylum is metabolically diverse, could operate chemoheterotrophy and anaerobic phototrophy, fix inorganic CO2, and oxidize nitrite (Rao et al., 2022). Gemmatimonadetes phylum has been retrieved from diverse habitats, including saline-alkaline soils and marine sediments (DeBruyn et al., 2011; Pascual et al., 2018). Cyanobacteria phylum is the only prokaryote to operate oxygenic photosynthesis (Sánchez-Baracaldo et al., 2022). Cyanobacteria phylum is only found to be significant in sediment samples (S60m, S20m) and the closest transect soil to the shoreline (T5m), which is in line with the distribution of Cyanobacteria phylum usually in the water column as phytoplanktons.
The controlling mechanism of prokaryotic community structure
Great differences in the prokaryotic community network between biotopes (Fig. 6 and Table 3) should partly account for different prokaryotic community structures of different samples. The symbiotic relationship indexes of the three networks are all higher than 50%, which shows that the prokaryotic communities of Barkol Lake mostly maintain a symbiotic relationship (Wu et al., 2020). Prokaryotic communities of soil samples have the weakest symbiotic relationship, while those of water samples have the strongest. However, with a higher value of Robustness index, the network of soil samples is considered more stable than the water and sediment networks (Sun et al., 2021). From soil to sediment or water, we can consider a process of salt lake development that corresponds to the evolution toward a weaker, though more symbiotic, network with fewer OTUs and lower indices of Chao1 and Shannon.
The distance from the shoreline and soil depth can also influence the prokaryotic community structures, which may be attributed to the differences in environmental factors between samples. For example, the decreases in salinity with the distance from the shoreline and with soil depth could be conducive to bacterial phyla Actinobacteriota, Chloroflexi, and Gemmatimonadota but lower the abundance of bacterial phylum Desulfobacterota. Variations in TS, pH, and Ca2+ could also influence the occurrences of archaeal and bacterial phyla of Crenarchaeota, Halobacterota, Bacteroidota, and Gemmatimonadota. This is consistent with previous studies that have indicated the prokaryotic community structure is possibly related to changes in salinity, pH, moisture, TOC, and TS as well as dissolved oxygen (not measured here) (Zhong et al., 2016; Banda et al., 2020).
The predicted ecological functions
Prokaryotic communities in Barkol Lake have a wide range of ecological functions, and almost half of the prokaryotes in Barkol Lake are attributed to chemoheterotrophy (Fig. 8). The archaeal and bacterial communities both took part in the carbon cycle, such as methanogenesis, photoautotrophy, and hydrocarbon degradation. Meanwhile, the high concentrations of sulfate in lake water, sediment, or soil samples of Barkol Lake can promote the involvement of prokaryotes in oxidation and respiration of sulfur compounds to maintain an active sulfur cycle such as in Mahoney Lake (Canada) and La Hoya del Rincón de Parangueo maar Lake (Mexico) (Klepac-Ceraj et al., 2012; Claudia et al., 2020). Bacterial communities could have played a more important role in the sulfur cycle. For example, the sample W120μm had a high proportion of bacterial communities assigned to dark oxidation of sulfur compounds (13.5%). The Gammaproteobacteria class (Proteobacteria phylum), which is dominant in our sediment samples and accounts for around 0.1% in our transect and profile soil samples, is one of the four known haloalkaliphilic sulfur-oxidizing bacteria that can help with the oxidation of subsulfide (Sorokin and Kuenen 2005); the Desulfobacterota phylum is also known to play a vital role in respiratory processes that utilize sulfate, sulfite, thiosulfate, or elemental sulfur (Murphy et al., 2021). Instead, archaeal communities are more important in the nitrogen cycle, while only a small number of bacterial communities were predicted to participate in the nitrogen cycle, mainly nitrate reduction. The archaeal class Nitrososphaeria (Crenarchaeota phylum) is abundant in Barkol Lake and may be involved in ammonia oxidation.
Conclusions and Implications
In the present study, the structure, abundances, interactions, and ecological functions of the prokaryotic communities in a hypersaline lake were thoroughly investigated. Barkol Lake is dominated by archaeal communities of Halobacterota, Nanoarchaeota, Thermoplasmatota, and Crenarchaeota and bacterial communities of Proteobacteria, Bacteroidetes, Actinobacteriota, Desulfobacterota, Chloroflexi, Gemmatimonadota, Firmicutes, and Cyanobacteria. The prokaryotic community structures differ significantly between biotopes (water, sediment, and soil). Halobacterota, Proteobacteria, Bacteroidota, Actinobacteriota, Firmicutes, and Cyanobacteria are the specialized phyla in water, while Thermoplasmatota, Desulfobacterota, Chloroflexi, and Gemmatimonadota are the specialized phyla in sediment and soil samples. The prokaryotic community network for soil is far more complicated and stable than those for water and sediment, while water and sediment samples of prokaryotic communities tend to develop stronger symbiotic relationships. Soil prokaryotic communities may be significantly affected by pH, salinity, TS, and Ca2+. In such a unique environment, archaeal communities tend to play an important role in the nitrogen cycle, while bacterial communities mainly participate in the sulfur cycle.
With the high salt concentrations as well as the arid and cold climate, Barkol Lake can be an ideal analog for ancient saline lakes on Mars and a potential Mars-like shelter for extreme life. Barkol Lake harbors brines with considerable Na+, Ca2+, Mg2+, SO4 2-, and Cl- ions along with a mineral assemblage of mirabilite-halite-gypsum-magnesium sulfate similar to brines and mineral compositions predicted to have occurred on Mars (Vaniman et al., 2004; Pontefract et al., 2017). Chloride minerals can significantly lower the freezing temperature (up to -50°C) of water due, in all likelihood, to supercooling processes (Brass, 1980; Buffo et al., 2022) that may act to preserve liquid brines during extremely low-temperature events (historic minimum: -43.6°C) in Barkol Lake (Wei and Wang, 2018). This may also be an important mechanism with the potential to acclimate life to the daily severe temperature changes on Mars. In addition, an abundance of sulfate and chloride minerals is known to host massive halophiles in Barkol Lake (Wei et al., 1998), and it is possible that such conditions could act, on Mars, as a refuge for martian counterparts by abating the effects of UV radiation and low temperatures and providing significant moisture (Parnell et al., 2004; Edwards et al., 2005; Osterloo et al., 2010). Although a number of Mars-related studies have considered the potential for acidic sulfate-rich lakes on Mars, it would seem that Barkol Lake, which is slightly neutral to alkaline, is potentially analogous to a number of martian sites, such as subsurface environments, given that acidification usually occurs at the surface (Hurowitz et al., 2010; Pontefract et al., 2017; Aerts et al., 2019; Buffo et al., 2022). Widespread, desiccated salt lakes on Mars are candidate sites as well for the search for evidence of martian life (Pontefract et al., 2017).
Further investigation of prokaryotic communities that are adaptive to a hypersaline environment, such as that of Barkol Lake, will help extend the data set of prokaryotic community structure for hypersaline environments and provide insightful perspectives into their counterparts, microbial interactions, and biogeochemical cycles on Mars (Oren, 2014). For example, the halophilic prokaryotic communities in Barkol Lake (e.g., Halobacterota, Thermoplasmatota, Crenarchaeota, Actinobacteriota, and Firmicutes phyla) may theoretically be similar to those that survive in liquid brines or desiccated salt lakes that are (or once were) present on Mars, which is essential for martian life search and identification. The differences in the prokaryotic community network between biotopes in Barkol Lake suggest that microbial communities tend to form simpler, weaker, though more symbiotic relationships, during aridification and desiccation processes that likely occur on Mars. As observed in Barkol Lake, there could be large varieties in prokaryotic community structures, should they exist, that span changing environmental conditions on Mars. The predicted ecological functions of only nitrification and nitrate reduction mainly by archaeal communities (e.g., Nitrososphaeria class) suggest that the nitrogen cycle in Mars-like environments is not as dynamic as typical terrestrial systems, which is in line with recent studies in hyperarid desert environments (see Shen et al., 2022 and references therein). Rather, the predicted ecological functions of dark oxidation and respiration of various sulfur compounds, mainly by bacterial communities (e.g., Gammaproteobacteria class and Desulfobacterota phylum), suggest the potential for an active sulfur cycle and could inform with regard to important mechanisms of sulfate mineral formations in Mars-like environments.
Footnotes
Acknowledgments
We thank Liyu Zhang and Xuejiao Qiao at Peking University Shenzhen Graduate School as well as Ying Wang from Environmental Ecology laboratory at Sun Yat-sen University for assistance in the lab and field.
Author Contributions
Wenqi Cai conducted the geochemical analysis and bioinformatic analysis, and wrote the original manuscript.
Ke Yu participated in the sample collection, supervised the 16S rRNA gene sequencing and bioinformatic analysis, and wrote the original manuscript.
Wanting Yang helped with the geochemical analysis and bioinformatic analysis.
Rong Mu performed DNA extraction, coordinated 16S rRNA gene sequencing, and contributed to manuscript editing.
Chunyang Lian helped with network analysis and contributed to manuscript editing.
Luhua Xie contributed to sample collection, geochemical analysis, and manuscript editing.
Yan Yan contributed to sample collection, geochemical analysis, and manuscript editing.
Shibin Liao contributed to sample collection and manuscript editing.
Fan Wang designed the study, performed the sample collection, oversaw the study plan, and led the manuscript writing and editing.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Ke Yu was financially supported by the National Natural Science Foundation of China (51939009). Fan Wang was financially supported by the Fundamental Research Funds for the Central Universities from Sun Yat-sen University (2021qntd29).
Associate Editor: Don Cowan