
Abstract
A new chiral amplification mechanism based on a stochastic approach is proposed. The mechanism includes five different chemical species, an achiral substrate (A), two chiral forms (L, D), and two intermediary species (LA, DA). The process occurs within a small, semipermeable compartment that can be diffusively coupled with the outside environment. The study considers two alternative primary sources for chiral species within the compartment, one chemical and the other diffusive. As a remarkable fact, the chiral amplification process occurs due to stochastic fluctuations of an intermediary catalytic species (LA, DA) produced in situ, given the interaction of the chiral species with the achiral substrate. The net process includes two different steps: the synthesis of the catalyst (LA and DA) and the catalytic production of new chiral species from the substrate. Stochastic simulations show that proper parameterization can induce a robust chiral state within the compartment regardless of whether the system is open or closed. We also show how an increase in the non-catalytic production of chiral species tends to negatively impact the homochirality degree of the system. By its conception, the proposed mechanism suggests a deeper connection with how most biochemical processes occur in living beings, a fact that could open new avenues for studying this fascinating phenomenon.
Introduction
Homochirality is one of the most distinctive features of terrestrial life at the molecular scale (Blackmond, 2019). Even when most of the functional organic molecules in living beings are chiral and, therefore, can exist in two different enantiomeric forms, living beings show an almost exclusive preference for only one. For instance, whereas amino acids, the key components of the proteins, are left-handed (L-amino acids), energy-rich compounds like sugars are exclusively right-handed (D-sugars), even when they share the same physical-chemical features with their isomer counterpart, excluding the interaction with other chiral species.
Many studies have addressed this issue from very different points of view. They include the chiral biasing associated with parity violations (Bonner, 2000), the influence of the astrophysical environment (Bailey et al., 1998; Bailey, 2001), or the use of much different thermodynamics (Hazen et al., 2001; Viedma et al., 2008; Weissbuch and Lahav, 2011) and kinetic considerations (Sandars, 2003; Gleiser and Walker, 2008, 2012). Regrettably, none of them give a universal solution to this amazing problem. The origin of such asymmetry continues to be one of the greatest puzzles in the study of life. In fact, it is not entirely clear whether homochirality preceded or followed the emergence of life (Laurent et al., 2021). However, homochirality is an indispensable condition for all known forms of contemporary life. Let us bear in mind that all the functioning of the metabolic, hereditary, and reproductive machinery at the cellular level is adjusted to the interaction between specific enantiomers. Without this level of specificity, life could not exist.
In 1953, in his seminal work (Frank, 1953), Frank introduced the first homochiral model based exclusively on kinetics considerations. The success of the Frank model relies on two key components: autocatalysis and chiral cross inhibition, mutual antagonism between L and D enantiomers. Several extensions based on the original Frank model have been proposed in the literature during the last decades. In some cases, autocatalysis is abandoned (Sugimori et al., 2008), while in others, it is the enantiomeric cross-inhibition term (Jafarpour et al., 2015). Another important number of studies focus their attention on the intrinsic fluctuations capable of inducing homochiral development (Saito et al., 2007; Fuss, 2009), the role of catalytic cycles and recycling in the dynamics (Micskei et al., 2008; Stich et al., 2016), and the behavior of open systems where both deterministic (Lente, 2010) and stochastic approaches (Lente and Ditrói, 2009) are used.
In principle, it is difficult to discern which is the most appropriate approach for studying this type of phenomenon. The choice largely depends on the conditions of the particular model to be considered. In general, stochastic models tend to be more challenging to approach mathematically (see for analytical treatment Lente [2004, 2005] Shao and Liu [2007]) and often require computationally extensive simulations. However, unlike the deterministic approach, these have the potential to capture dynamics better when systems are highly dilute (Gillespie, 2007; Sun et al., 2008), a condition most likely in primordial environments.
However, beyond the theoretical aspects, probably the most important limitation for studies on the origin of homochirality lies in the lack of experimental evidence. Unfortunately, reactions like that considered by Soai et al. (1995) have little or no relevance in prebiotic environments (Blackmond, 2019). The controlled synthesis of the chiral octahedral cobalt complex is another of the few known systems capable of inducing a homochiral state under suitable conditions (Asakura et al., 2001). Compared to the Soai reaction, the intrinsic mechanisms behind this system could have significant implications for astrobiological studies. In any case, the study and proposal of new theoretical models more adjusted to the environmental conditions of early Earth will probably continue for a long time.
In this sense, the present work proposes an alternative mechanism to explain the emergence of homochirality at a small scale. Unlike other studies, the chiral amplification mechanism relies on intermediate catalytic species produced in situ due to the interaction of the chiral species with the substrate. The use of intermediate catalytic species in the chiral amplification process suggests a deeper connection with most of the enzymatic reactions and metabolic processes that operate in living beings.
The Model
The model considers the existence of a small and well-mixed compartment surrounded by a semipermeable membrane. All chemical processes are confined to the interior of the compartment. The system includes five different chemical species: an achiral substrate A, two chiral species (L, D), and two intermediate catalytic species (LA, DA). The two catalytic species, LA and DA, are confined to the interior of the compartment. The general model comprises the following three fundamental stages.
Stage 1: The primary non-catalytic source for the chiral species
Includes non-catalytic mechanisms to produce chiral species. In our particular case, two different types of sources are introduced:
2.1a. The existence of an internal chemical source
Chiral species (L, D) are produced from the substrate A through a reversible non-catalytic process according to
where k0 and k_0 are the rate constants for the forward and reverse processes, respectively.
2.1b. The existence of an external diffusive source
The chiral species (L, D) and the substrate A can flow through the semipermeable membrane in correspondence with Fick's law. This characteristic allows diffusive coupling of the system with the external environment. Outside the compartment, we assume that the concentrations of these species are buffered at constant values A0, L0, D0. This process can be represented schematically as
being Fa and Fch, the diffusion coefficients for the achiral and chiral species, respectively. The diffusion treatment considered here is similar to that used by Lente (2010) with the difference that, in our particular case, the concentrations of chiral species outside the compartment are not null. The isolated system appears as a particular case when both coefficients (Fa, Fch) are zero. Although this step is not an essential part of the proposed chiral amplification mechanism, as we will see later, the transport regime can significantly modulate how this process occurs.
Stage 2: The production of the catalytic species LA and DA by a reversible process
where k1 and k_1 are the rate constants for the forward and reverse reactions, respectively. Both species play a fundamental role within the proposed chiral amplification scheme. The possible transit of the system to a homochiral state will depend on how fast they are produced; their stability once formed, and their catalytic potential (stage 3).
Stage 3: The reversible catalytic production of the chiral species L and D
where k2 and k_2 are the corresponding rate constants. The process is assumed to be selective (fidelity = 1); the intermediate species only catalyze the production of chiral species of the same handedness. According to stage 3, the efficiency of the catalytic process depends not only on the rate constant k2 but also on the concentration of the intermediate species in the medium (stage 2). Consequently, we should expect that the chiral amplification mechanism works better for those systems where both constants (k1, k2) reach high values. Similarly, increasing non-catalytic contributions to the process (step 1) should negatively impact the enantiomeric excess.
However, the parameter domain has some important restrictions when the primary chiral species come from a non-catalytic chemical process (stage 1a). For instance, it is necessary to consider that not all the rate constants involved in the model are independent. If we take into account that the free energy variation is independent of whether the process is catalytic or not, we can infer a relationship between the rate constants that govern both processes (see Stich et al. [2016] for a more detailed discussion). Taking as reference the equilibrium state where ΔG
0 = -RT Ln(Keq) and applying it to both catalytic (stage 3) and non-catalytic production (stage 1a), the following relationship is obtained:
which reduces the total number of constants included in the model by one. In particular, we prefer to consider k_2 as a dependent constant during this study. Note that the constants k1 and k_1 are not subject to a relationship of this type since they are not directly involved in the catalytic process but rather in the synthesis of the catalyst.
On the other hand, it is important to point out that these restrictions only apply when the primary chiral species are produced totally or partially through an internal non-catalytic chemical reaction (stage 1a). For the same reason, the above restrictions do not apply when the primary chiral species are produced exclusively by an external diffusive source (stage 1b).
During the next sections, we will study the evolution of the system considering the previous arguments in the parameterization. To model the system, the volume of the compartment was set at 5.2 * 10−19 dm3, a value consistent with the existence of a spherical vesicle reactor of 100 nm in diameter. We also consider that there are no chiral species within the compartment initially (L(0) = D(0) = 0). A numerical stochastic approach was used, considering that the amplification process starts with very few molecules. To do this, the model described above was implemented in Python using the benefits of the stochastic simulation code Gillespy2 (Abel et al., 2016). Designed for the modeling of biochemical systems, the code allows efficient generation of exact numerical solutions to the master equation based on Gillespie's stochastic simulation algorithms. Another advantage of Gillespy2 is the possibility of integrating the Python resources for the analysis of the results without resorting to other software. During this work, we used the gillespy2 to compute how the mean values of the enantiomeric excess
Two different scenarios are considered in our analyses. First, the closed system provided with a primary chemical source is considered (stage 1a). Subsequently, the open system is studied considering the existence of an external diffusive source (stage 1b). The results of these procedures are discussed in detail in the following sections.
Results
The closed compartment with an internal non-catalytic chemical source (Fa = Fch = 0)
Here, we explore how the mean values of the enantiomeric excess evolve as a function of the parameters k1 and k2. Figure 1 shows how the mechanism of chiral amplification works as a function of the parameter k1. The other parameters included in the simulation were set to (A = 104, D = L = DA = LA = 0, k0 = 10−3, k_0 = 10−11, k_1 = 1, k2 = 106). The value of k_2 was estimated from the thermodynamic constraints between the rate constants as k_2 = k2 (k_0/k0).
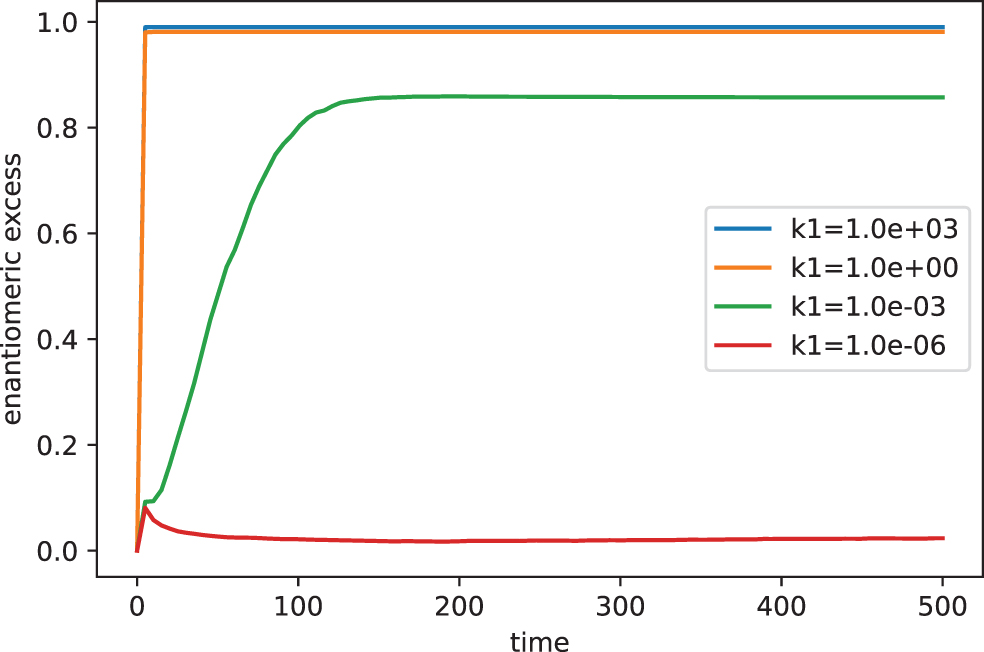
Chiral evolution for different values of the rate constant k1 (105 runs).
According to the figure, as the value of k1 increases, the system tends to reach a perfect homochiral state. In general, for values of k1 > 1, the enantiomeric excess reaches values very close to unity. On the other hand, the figure also shows that it is possible to reach a practically racemic state for sufficiently small values of the parameter k1. The existence of such trends is clearly confirmed in Fig. 2, where the stationary distributions of the enantiomeric excess are shown as a function of the parameter k1.
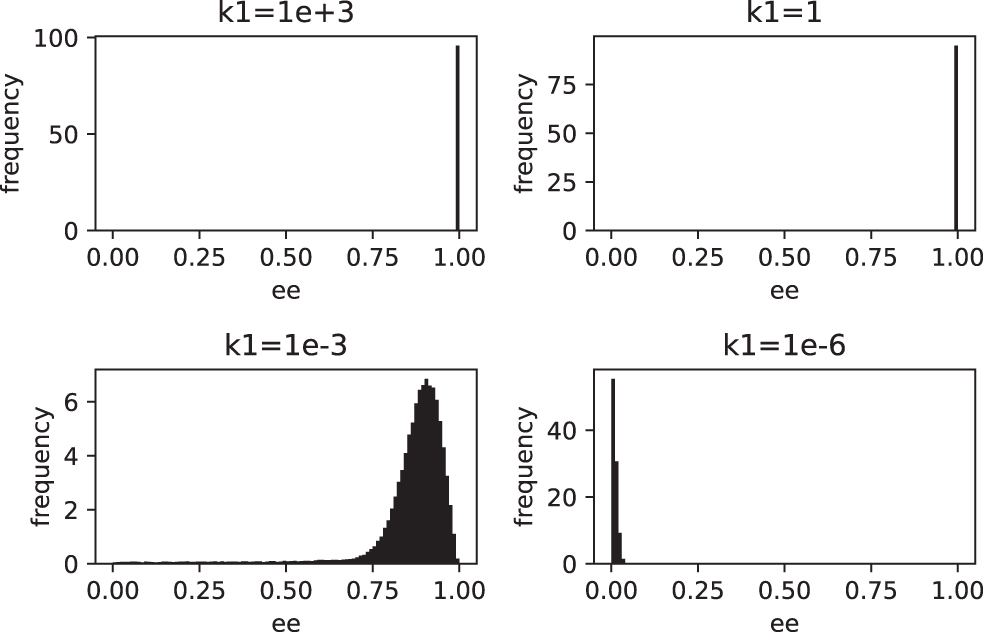
Stationary distributions of the enantiomeric excess for the steady state as a function of the parameter k1 (time = 500, samples = 105).
According to the information shown in Fig. 2, for k1 ≥ 1, the ee values cluster very close to the unity (
Here, we explore how the parameter k2, a measure of the catalytic capacity, impacts on the dynamics of the system. The results of such a procedure are shown in Fig. 3 assuming the following parametrization: (A = 104, D = L = DA = LA = 0, k0 = 10−3, k_0 = 10−11, k1 = 1, k_1 = 1).
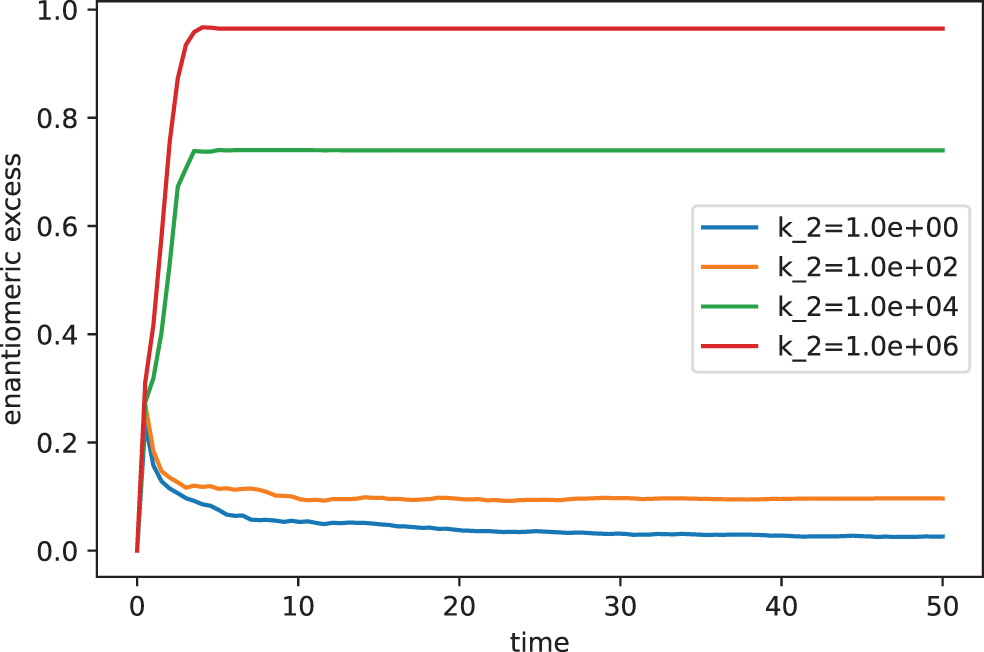
Chiral evolution for different values of the rate constant k2 (105 runs).
As can be seen, Fig. 3 reveals the importance of k2 within the mechanism of chiral amplification. Note that the model evolves in two different ways according to specific range of the parameter k2. While for values of k2 > 10 the enantiomeric excess reaches significant values, for k < 1 the values are remarkably low. Note that, only for enough high values of k2 = 106 the system reaches an almost-homochiral state
Once again, this behavior is consistent with the stationary distributions obtained for each value of the parameter k2 used during the simulation (see Fig. 4).
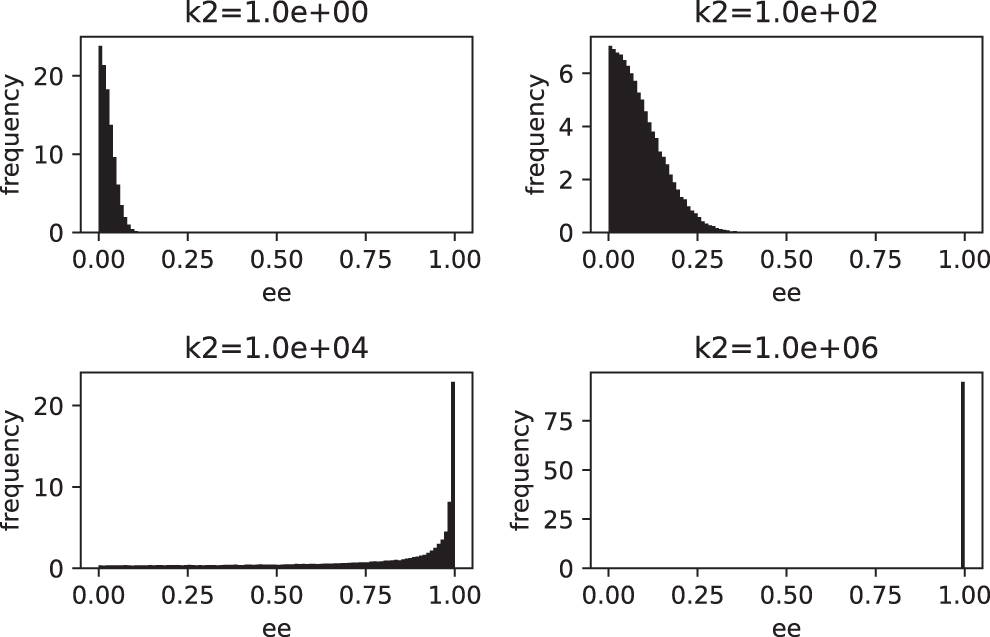
Stationary distributions of the enantiomeric excess for the steady state as a function of the parameter k2 (time = 50, samples = 105).
Figure 4 shows the gradual transition, from a practically racemic state to an almost perfectly homochiral one, as the parameter k2 grows. Note how the spread of the data grows for the intermediate stages (k2 = 102, 104 ) as a measure of the loss of stability.
Opposed to the closed variant, the model for the open compartment includes the fluxes of the three main species (A, L, D) with the external medium. We are also assuming that there is no primary chemical source within the compartment. Correspondingly, the input of primary chiral species into the compartment comes exclusively from the external environment. For the sake of simplicity, we consider that the populations of these species in the surroundings of the compartment are buffered, setting their values to A0 = 104 and L0 = D0 = 200. Within the compartment, the number of particles was initially set to A(0) = 104, L(0) = D(0) = 0, assuming that substrate A is the only species in equilibrium with the external medium
Here, we explore how the system behaves when the diffusion coefficients for chiral species are changed. Figure 5 shows the chiral evolution of the system as the chiral diffusion coefficient Fch decreases. The other parameters used in the simulation were set to (Fa = k_1 = 1, k1 = 10−2, k2 = 106, k0 = k_0 = k_2 = 0).

The impact of reducing the diffusion coefficient Fch on chiral evolution (105 runs).
According to the figure, as the flux of the chiral species across the boundary is more limited, the possibility of inducing a homochiral state increases. The figure also reveals a markedly nonlinear behavior (initial oscillation) of the system for small values of the parameter Fch (Fch ≤0.1). This kind of behavior evidences the complex interaction between the chiral amplification mechanism and the transport processes across the compartment boundary. In general, the enantiomeric excess reaches values very close to the unity when the parameter Fch is small (Fch <0.1). Also note that, despite the constant input of both enantiomers to the compartment, only one of them is able to grow. The population of the nonfavored isomer remains low, keeping values very similar to those imposed by the external source. Even when we focus our attention on the permeability of chiral species, we also find similar trends increasing the permeability of the substrate Fa or the substrate concentration A0 in the external medium.
Finally, we must point out that the reverse is also true. The existence of relatively high fluxes of chiral species may erase any asymmetry between both enantiomers. For instance, according to Fig. 5, the enantiomeric excess becomes practicably negligible
In the end, we explore how the dynamics of the open system impact the catalytic capacity (k2) assuming an appropriate value of Fch. The results are shown in Fig. 6 considering the following parametrization (Fa = k_1 = 1, Fch = 10−2, k1 = 10−2, k0 = k_0 = k_2 = 0).
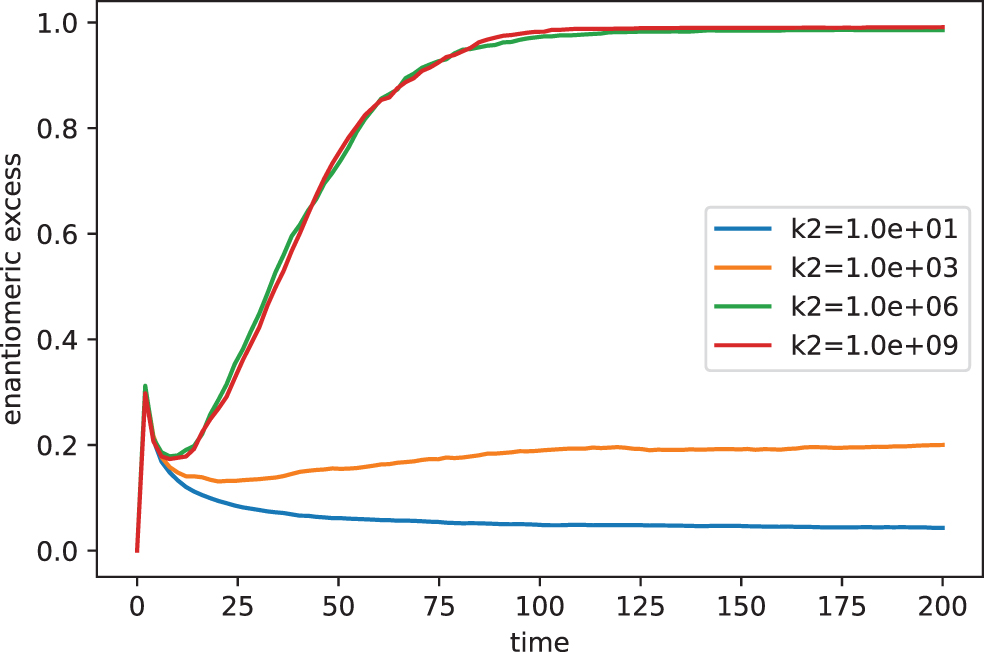
Evolution of chirality for the open system for different values of the catalytic efficiency parameter k2.
In general, the results shown in Fig. 6 are in agreement with those derivate previously in Fig. 3. Once again, the increment of the catalytic capacity of the intermediate species impacts positively in the chiral amplification process. Similar to Fig. 4, the behavior is practically split by k2 in two well-defined regions, a low homochirality degree region ee ≤0.18 for k2 ≤ 103 and a high homochirality degree region ee ≥0.98 for k2 ≥ 106.
The results obtained in the previous section show the fundamental role of the parameters k1 and k2 within the proposed chiral amplification mechanism. In this sense, the dependence of two kinetic parameters marks a notable difference from previous mechanisms where the catalytic process usually involves only one stage. The analysis of the curves shows, both in open and closed systems, how the efficiency of the process increases when both parameters increase. The analysis also shows a gradual reduction in the time required to reach the homochiral state, a dependence that is more marked for the parameter k1 (see Fig. 1), a behavior consistent with the structure of the proposed mechanism. Note that the amplification process only starts with the formation of the first catalyst molecules (LA or DA), a process controlled by the rate constant k1.
It is also important to highlight the similarities between the open and closed system behavior as a function of the parameter k2, as shown in Figs. 3 and 6. Accordingly, the chiral evolution of the system is practically independent of the way in which the primary chiral species are generated, either chemically or diffusively. In principle, regardless of the source type used, the chiral amplification process begins with the production of a first chiral molecule and the subsequent formation of an intermediate species (LA or DA). For example, consider that a molecule of the LA type is first formed. Then, in correspondence with the proposed scheme, the LA species reacts with another A substrate molecule to transform it into an L species. This process can be repeated several times for the same LA molecule if we assume that the intermediate species formed is sufficiently stable and that the catalytic reaction occurs at a high rate compared to the non-catalytic one. In turn, each new L molecule formed reacts with another substrate molecule to again form another LA molecule in a continuous loop. In consonance, the concentrations of LA and L species grow quickly, whereas the amount of available substrate in the medium decreases noticeably. At this moment, if we assume the creation of a second intermediary molecule of the opposite type DA, it must compete for a depleted substrate against a much higher population of LA molecules. Consequently, the DA molecule will have a greater possibility of dissociating again into its components, releasing a new substrate molecule into the medium that will probably be rapidly transformed into an L species through a chiral LA chain.
It is important to reaffirm that all these results are obtained assuming that, initially, there are no chiral species within the compartment. However, these trends could vary if there is an appreciable concentration of these species within the compartment at the beginning of the process. In these cases, the possibility of reaching a homochiral state increases as the value of k1 decreases. Although apparently paradoxical, this behavior does not contradict previous results. For example, if we start from an appreciable concentration of both enantiomers within the compartment, the possibility of inducing catalytic chains for both isomers through the intermediate species LA and DA is high if the value of k1 is large. This fact reduces the chances of generating an appreciable enantiomeric excess within the system. The effective way to reduce this possibility is to decrease k1 so that the formation of these species (LA or DA) is a highly improbable event in the early stages of the process. Once one of them is formed, the catalytic production process is activated, favoring one of the two isomers, as described above.
Consequently, it is obvious that the required values of k1 and k2 to induce a chiral amplification mechanism vary depending on the productivity of the primary sources and the possible accumulation of chiral species initially. In general, for the cases considered in our study (L(0) = D(0) = 0), the chiral amplification process is favored when both parameters increase. The same principles apply in an open system when considering two primary sources, one external and one internal. An increase in the net rate of enantiomer production by any non-catalytic route will always tend to decrease the degree of homochirality within the system. However, we must consider that including an internal chemical source, even partially, imposes additional thermodynamic restrictions on the rate constants involved in catalytic and non-catalytic processes.
Finally, we cannot forget that k1 and k2, as second order rate constants, are subject to important theoretical restrictions (Bar-Even et al., 2011). Beyond these theoretical limits (108 to 109 mol−1 dm3 s−1) it is not possible to reach realistic conclusions. However, the theoretical margin seems to be wide enough to obtain a reproducible homochiral state in most real situations.
Conclusions
Homochirality is perhaps one of the most fascinating and tangible examples of broken symmetry in nature. Paradoxically, and despite the multiple efforts made in this direction, the origin of homochirality in living beings is far from being a well-understood topic. In this way, we propose a rather alternative theoretical mechanism capable of explaining some key elements of this interesting phenomenon. In our case, the chiral amplification process is intrinsically linked to the dynamics of the intermediate catalytic species, where stochastic fluctuations play a fundamental role. In this sense, introducing a chiral intermediate species within the autocatalysis mechanism opens new possibilities for studying this type of system. Unlike the classical mechanism, where the same chiral species acts as a catalyst, introducing an intermediate species favors the chiral breakdown of the system. These potentialities are due to the fact that the system must reach a previous stage (the formation of LA or DA) before establishing an effective autocatalytic chain to produce a specific enantiomer (L or D). In the most remarkable cases, the system can reach a practically homochiral state with values of ee very close to unity. It is important to highlight that this behavior persists for the open system when moderate constant flows of both enantiomeric forms are imposed from the outside. Furthermore, the behavior for the open version is also evidence of the noticeable role of the membranes and the selective transport as modulators of the whole process. For this particular case, homochirality's rise could depend critically on these factors. It is important to note that encapsulation is considered a crucial element for the development and subsequent evolution of prebiotic systems (de Souza et al., 2014; Shirt-Ediss et al., 2015; Piedrafita et al., 2017; Martín et al., 2021).
On the other hand, the model of chiral amplification proposed here could have certain points of contact with the theory of metabolic replacement (M,R) systems (Piedrafita et al., 2010). Accordingly, the catalysts for each metabolic step must be produced in situ, a condition that ensures metabolic closure. Likely, this condition was already present in the most incipient forms of life during early Earth. In this sense, the proposed mechanism approaches, at least formally, this typical structure. Perhaps the origin of homochirality could be more linked to the architecture of these large networks (Strogatz, 2001) than the result of specific interaction mechanisms. In this sense, the present work can contribute positively to understanding this fascinating phenomenon that permeates, without exception, the essence of life on our planet.
Footnotes
Acknowledgments
Y.L. acknowledges Dirección de Investigación, Postgrado y Transferencia Tecnológica de la Universidad de Tarapacá for financial support through Proyecto UTA Mayor N° 4724-22.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Author Contribution Statement
O.M. conceptualized the model and wrote the original draft, Y.L. funding acquisition, formal analysis, review and editing. J.S., Y.P., and Y.M. contributed through multiple rounds of discussion to the final version of the manuscript.
Associate Editor: Christopher McKay