
Abstract
We report abundant small calcareous mounds associated with fossilized kerogenous microbial mats in tidal-facies sandstones of the predominantly siliciclastic Moodies Group (ca. 3.22 Ga) of the Barberton Greenstone Belt (BGB), South Africa and Eswatini. Most of the bulbous, internally microlaminated mounds are several centimeters in diameter and formed at the sediment-water interface contemporaneously with sedimentation. They originally consisted of Fe-Mg-Mn carbonate, which is now largely silicified; subtle internal compositional laminations are composed of organic matter and sericite. Their presence for >6 km along strike, their restriction to the inferred photic zone, and the internal structure suggest that mineral precipitation was induced by photosynthetic microorganisms. Similar calcareous mounds in this unit also occur within and on top of fluid-escape conduits, suggesting that carbonate precipitation may either have occurred abiogenically or involved chemotrophic metabolism(s) utilizing the oxidation of organic matter, methane, or hydrogen, the latter possibly generated by serpentinization of underlying ultramafic rocks. Alternatively or additionally, carbonate may have precipitated abiotically where heated subsurface fluids, sourced by the intrusion of a major Moodies-age sill, reached the tidal flats. In summary, precipitation mechanisms may have been variable; the calcareous mounds may represent “hybrid carbonates” that may have originated from the small-scale overlap of bioinduced and abiotic processes in space and time. Significantly, the widespread occurrence of these stromatolite-like structures in a fully siliciclastic, high-energy tidal setting broadens search criteria in the search for life on Mars while their possible hybrid origin challenges our ability to unambiguously identify a biogenic component.
Introduction
Many of the earliest geochemical and morphological traces of life on Earth are found in carbonates; thus, high-Mg carbonates are among the prime targets in the search for potential biosignatures, for example, at Jezero Crater on Mars (Bosak et al., 2021). The biosphere's exploitation of geochemical gradients to extract building materials from inorganic precursors fundamentally altered Earth's planetary surface, providing the base for an immense variety of minerals, rock types, and geologic and climatic processes (e.g., Hazen et al., 2008). However, distinguishing between organic and inorganic precipitation, in particular where geochemical data are absent or compromised by weathering or metamorphism (Buick et al., 1981; Grotzinger and Rothman, 1996; Allwood et al., 2010; Hickman-Lewis et al., 2017; Gu et al., 2020), has proven problematic. Where biomediated carbonate precipitation can be established, metabolisms often remain uncertain; while methanogenesis (combining H2 from serpentinization with CO2 from seawater; e.g., Ueno et al., 2006; Sleep and Bird, 2007), sulfate reduction (e.g., Shen and Buick, 2004), and methanotrophy (oxidizing CH4 using, e.g., SO4 or NO3; e.g., Bergmann et al., 2013; Sauterey et al., 2020) are thought to be among the earliest metabolic pathways, it is far less certain when energetically potent, but complex photosynthesis became important and when, how, and where ingredients and products in the evolving metabolic microbial web became dependent on each other (e.g., Nisbet and Sleep, 2001; Sleep and Bird, 2007; Krissansen-Totton et al., 2015; Cardona Lodono, 2019). While laboratory experiments can demonstrate microbial interactions (e.g., Milucka et al., 2015) and genetics can—to some degree—reconstruct evolutionary microbial lineages (Schirrmeister et al., 2015; Betts et al., 2018; Cardona, 2018; Cardona Londono, 2019; Oliver et al., 2021), there is a paucity of “hard” geologic data extracted from suitable outcrops of Paleoarchean (>3.2 Ga) age because they are rare, geologically complex, and lie in highly deformed, pre-plate-tectonic volcano-sedimentary basins, so-called greenstone belts.
Here, we describe previously undocumented calcareous mounds among kerogenous benthic microbial mats in tidal-facies sandstones of the Paleoarchean Moodies Group of the Barberton Greenstone Belt (BGB; Heubeck, 2001, 2009; Noffke et al., 2006, 2022; Gamper et al., 2012; Homann et al., 2015, 2016, 2018; Homann, 2019; Homann and Heubeck, 2021). Their characteristics and occurrence above a cooling sill open a unique paleoenvironmental window on the evolution of shallow-water microbial life.
The Moodies Group
The Barberton Greenstone Belt (Fig. 1) in eastern South Africa and Eswatini, and the Pilbara region in NW Australia constitute the best-preserved regions on Earth to study Paleoarchean sedimentary and volcanic strata (e.g., Hickman, 2012; Byerly et al., 2018; van Kranendonk et al., 2018).
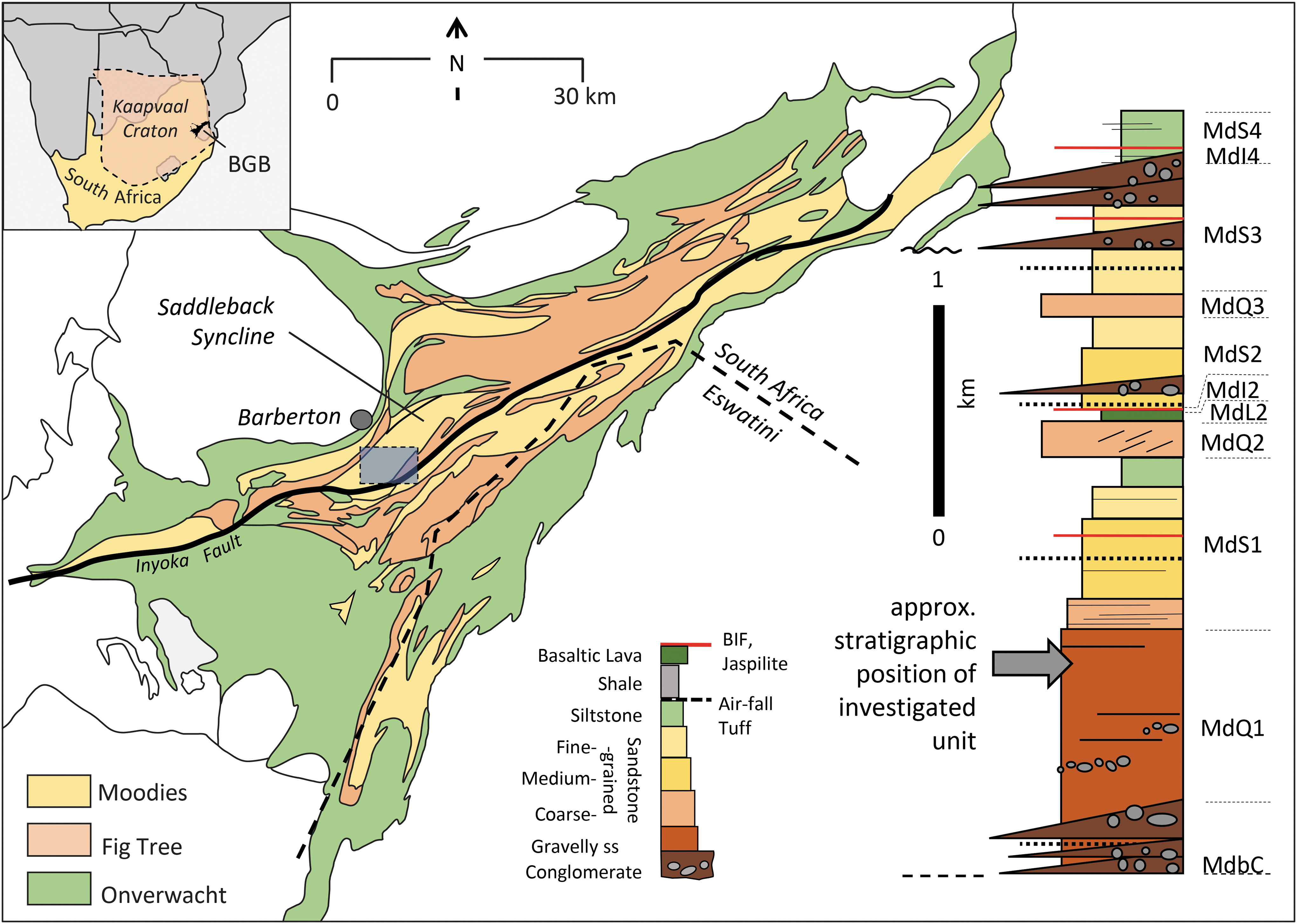
Geological map of the Barberton Greenstone Belt (BGB); gray rectangle shows the area in Fig. 2. Inset to the right shows a generalized stratigraphic column of the Moodies Group in the central BGB north of the Inyoka Fault.
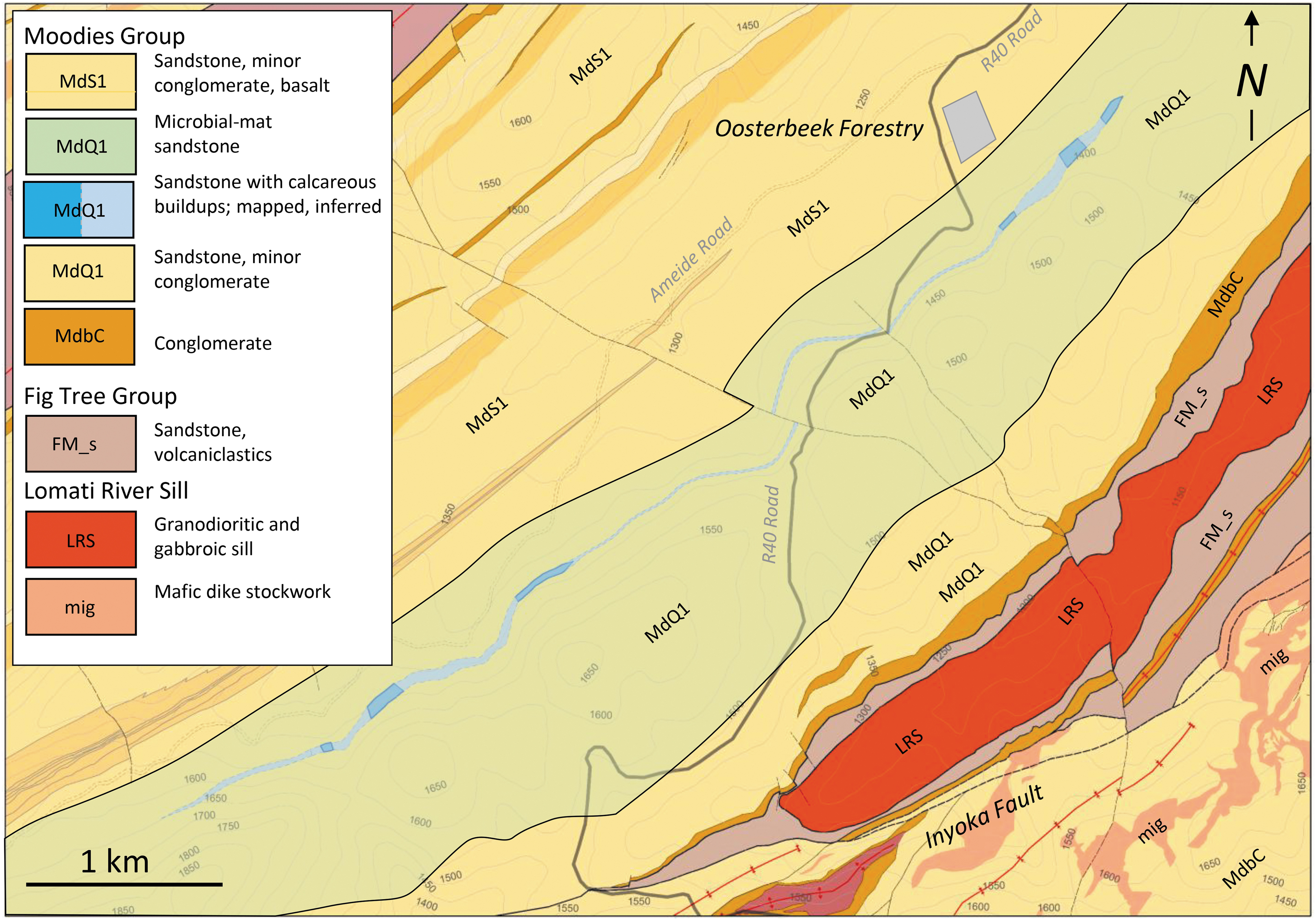
Map of central BGB, emphasizing (in blue) outcrop and mapped extent of the sandstone unit with calcareous mounds in the Saddleback Syncline of the central BGB.
The up to 3.6 km thick Moodies Group (Fig. 1 inset) is the uppermost and youngest of the three principal stratigraphic units making up the ca. 15 km thick Barberton Supergroup, the volcano-sedimentary fill of the BGB (Heubeck, 2019). It is one of the oldest well-preserved quartzose sedimentary units on Earth and one of the oldest siliciclastic shallow-water units. Moodies Group strata predominantly consist of fine- to coarse-grained, compositionally immature to mature and dominantly first-cycle quartzose sandstones deposited in alluvial, fluvial, deltaic, tidal, and shallow-marine environments (Heubeck and Lowe, 1994, 1999; Heubeck et al., 2016). Thick siltstones, subordinate jaspilites, banded iron formations, and shales reflect deposition below wave base, probably in pro-delta or similar settings. Strata generally dip subvertically. Strain is low or negligible in many places so that sedimentary structures are excellently preserved, in part due to widespread early silicification. Key sections of the Moodies Group were drilled in 2021–2022 during the ICDP BASE project.
Abundant and exquisitely preserved microbial laminations in Moodies tidal-facies sandstones are composed of dark green to black kerogenous matter which show clear Raman-spectroscopic D- and G-peaks of organic carbon (Corg) and fine-grained chloritic and sericitic mica (Noffke et al., 2006; Heubeck, 2009; Homann et al., 2015, 2016, 2018). Primary meso- and microstructures include millimeter-spaced undulatory and wispy laminae, tufts, domal bulbs, trapped gas bubbles, and polylobate textures. Microbial-chip conglomerates indicate desiccation and reworking of brittle microbial mat fragments in shallow, wide tidal channels. Common fluid-escape structures flexed the microbial mats, indicating a supple mechanical behavior at shallow depth. The geometric pattern of kerogenous mats in plane view appears clotty, without preferred directions. Bedding-parallel chert lenses up to 15 cm long and a few millimeters thick associated with microbial mats show internally wispy kerogenous lamination and occasionally remnant palisade-fabric carbonate crystals in thin section (Homann et al., 2016). They are interpreted to have formed as initially gas-filled, then fluid-filled “soft cavities” in wet, coherent shoreline sand laced by microbial filaments which were colonized by biofilms (Homann et al., 2016), subsequently filled by palisade-fabric carbonate, and are now nearly completely silicified. Carbonate precipitation predated compaction.
Geologic mapping of the unit and outcrop photography was complemented by sampling. Slabbing and polishing of hand samples was followed by thin sectioning, petrographic examination, and close-up photography using a Keyence VHX-6000 digital microscope. Elemental maps of selected slabbed structures were generated using a Bruker Tornado M4 Micro X-ray fluorescence (μ-XRF) instrument (50 keV tube voltage; 25 msec/pixel exposure time; 20 μ pixel resolution). Raman analyses were performed using a Renishaw InVia Raman microscope coupled to an Olympus BX61 confocal microscope within the PARI analytical platform at the IPGP. Measurements were made with a 514 nm Ar-ion laser and adjusted to an on-sample intensity of 0.2 mW with a spot size of <2 μm ( × 50 lens).
Results
Calcareous mounds associated with microbial mats
Outcrop- and hand-sample-scale: Detailed geological mapping in the Saddleback Syncline of the central BGB (Figs. 1 and 2) shows that abundant calcareous mounds occur in a ca. 20 m thick unit of medium- to coarse-grained tidal-facies sandstone near the top of unit MdQ1 (following the field terminology of Anhaeusser, 1976). The unit can be traced intermittently for >6 km along strike (Fig. 2) within commercial pine and eucalyptus plantations. It is underlain by up to 500 m thick microbially laminated sandstones and overlain by ca. 50 m of sandstones grading into tuffaceous sandstones of unit MdS1 (Fig. 1 inset; Fig. 2). Identical calcareous mounds also occur at a stratigraphically higher level (upper MdS1) in the Dycedale Syncline, a few kilometers to the north.
Mounds occur within medium-bedded, planar-bedded, and rippled microbially laminated sandstone. This host rock is quartz-rich, medium- to coarse-grained and quartz-carbonate-sericite-cemented; it has undergone little compaction. Mounds weather white and resistant in outcrop. The structures reach 1–5 cm in height and diameter (Figs. 3 and 4) and are randomly but approximately evenly interspersed at millimeter to decimeter distance from each other (Fig. 3A). Most structures are convex-up, bulbous, approximately lobate, and polylobate (Figs. 3 and 4). They are in places laterally linked to form small cumulate domes (Fig. 4); rarely, they are subvertically stacked (Fig. 3E). Crusts (i.e., thin, laterally extensive bedding-parallel structures) were never observed. Internally, many mounds show subparallel convex-up-laminations recognizable in outcrop (Fig. 3); others appear unstructured.

Largely silicified calcareous mounds in outcrop are resistant to weathering and white. (

Silicified mounds in polished hand sample from outcrop. Chert weathers white. (
In thin section, most structures consist of an equigranular mosaic of homogeneous interlocking micro- to mesocrystalline equigranular quartz (chert; Figs. 5A–5F and 6) in which stubby (sub)microscopic dolomite crystals and minor organic matter are evenly dispersed. Rarely, mounds consist entirely of relatively pristine Fe-Mg-Mn carbonates (ankerite; Fig. 6C, 6D) and show subtle internal kerogenous laminations that resemble microstromatolites (Figs. 6 and 7). Locally, palisade fabric of elongate carbonate crystals oriented perpendicular to lamination can be observed. Laminae adjacent to the mounds are defined by (sub)millimeter-thick kerogenous matter, single-grain-thin streaks of siliciclastic sediment enriched in finely dispersed organic matter, concentrations of optically opaque heavy minerals, very-fine-grained quartzofeldspathic sand grains, and sericitic, Corg- or Cr-rich silt-sized mica (Fig. 7); they typically rise and wedge out toward the mounds. Some laminae, however, persist through the core of the mounds and delineate a stacking pattern in cross section (Figs. 4D–4G, 6, and 7). These are best imaged in elemental maps (Fig. 7) where small-scale internal structures are enriched in Al and Cr, representing fuchsitic mica, an alteration product of clay particles derived from the weathering of ultramafic rocks.

Close-up photographs of calcareous mounds in polished slabs, demonstrating domal growth and silica replacement of remnant carbonate.
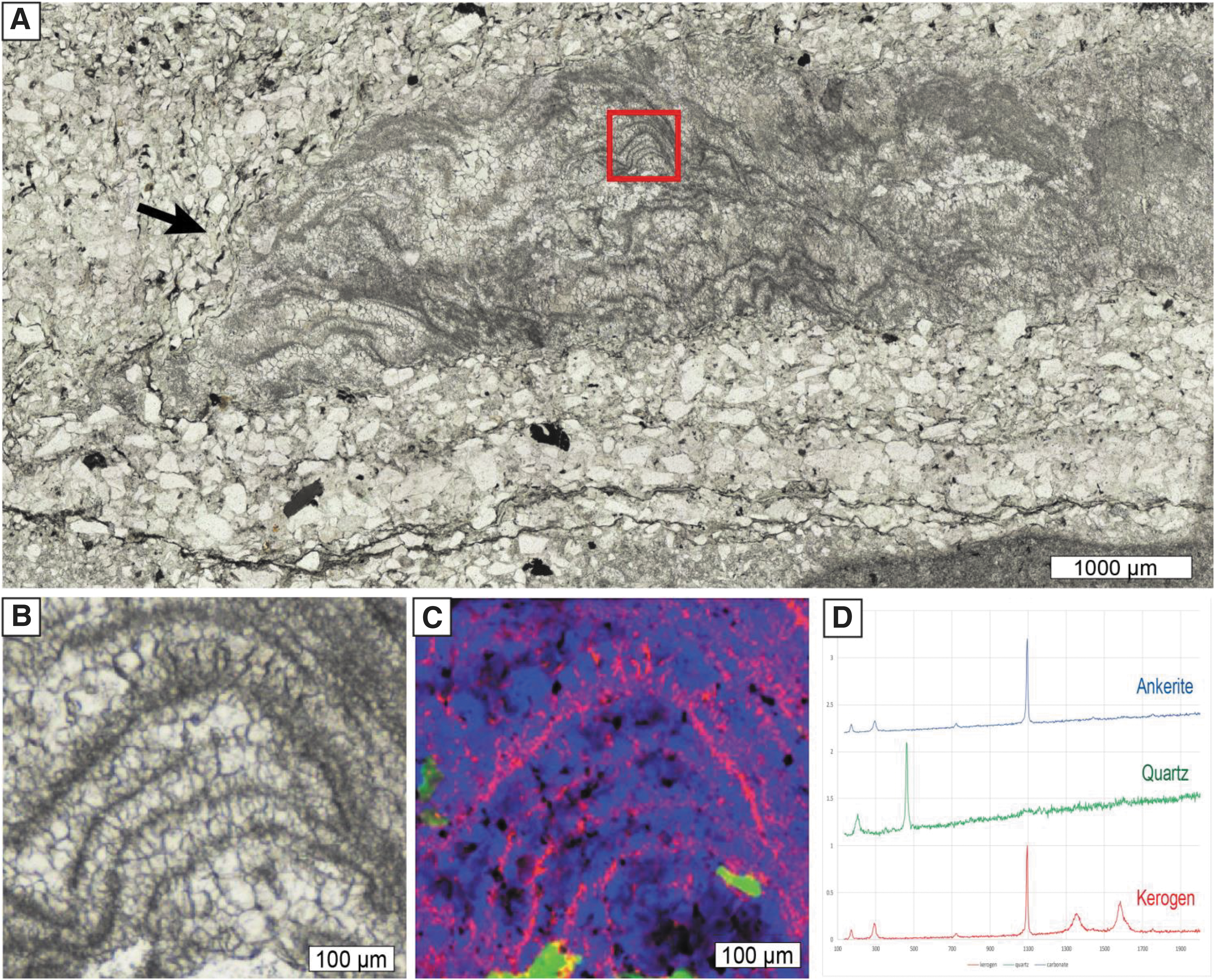
Thin-section photomicrographs (
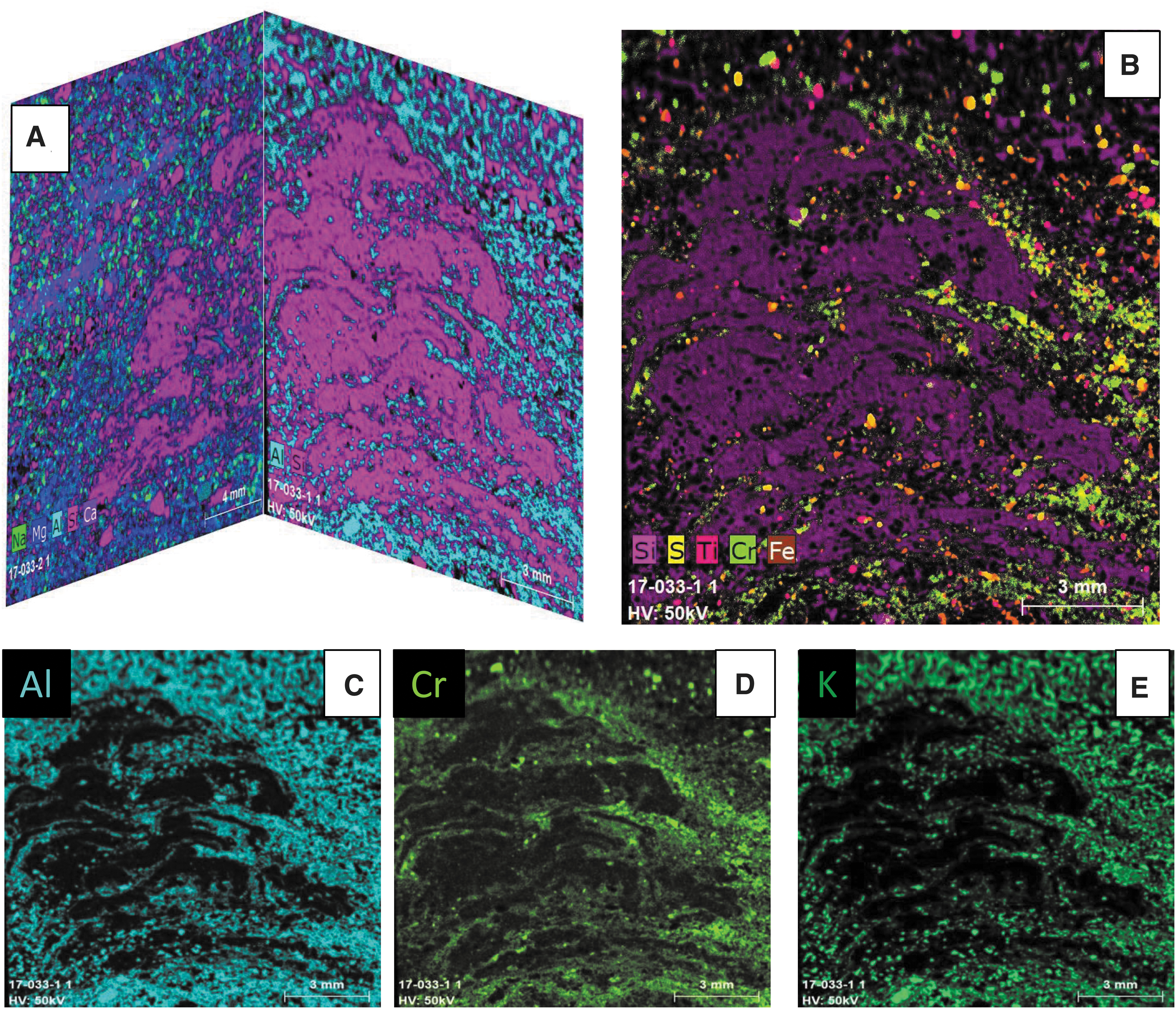
Micro X-ray fluorescence (μ-XRF) elemental maps of carbonate mound (sample 17-033). (
Silicified calcareous structures also occur in fluid-escape (dewatering) conduits of the same unit (Fig. 3E, 3F); these are common in microbially laminated sandstone in the BGB (Homann et al., 2015, 2018). In the proximal parts of the lateral feeders to the conduit, mounds form mostly stratiform, distally tapering, upward-curved masses 1 to 2 cm thick and several centimeters long, whereas they form less structured, lensoid, inverse-conical or spheroidal, evenly dispersed or loosely concatenated masses of similar dimensions in the main vertical dewatering conduit (Fig. 3F).
Discussion
Original composition and depositional environment
The location of precipitation, external and internal geometry, and the petrographic fabric and composition of the mounds suggest that fine-grained (micritic) calcite or ankerite precipitated from fluid locally oversaturated with respect to calcium carbonate at the sediment-water contact and within active fluid-escape structures while the siliciclastic sediment was unconsolidated and water-saturated. The onlap structures suggest that vertical growth occurred at the sediment-water interface and competed with sedimentation (Fig. 4A–4C; Fig. 7B, 7D, 7E). Early-diagenetic, pre-compaction silicification replacing carbonate cement is widely observed in the BGB, in particular in strata of the Moodies Group in the central BGB, where it was interpreted to be due to hydrothermal mobilization of Si above the cooling Lomati River Sill of Moodies age, which underlies the unit under discussion (Heubeck and Lowe, 1994; Fugmann, 2017; Homann and Heubeck, 2021; Reimann et al., 2021). Additional but largely syn- or post-compaction silica sources may have become available from surficial paleoweathering, smectite-to-illite diagenesis, pressure solution of chert and quartz grains during compaction, and post-orogenic dike swarms cross-cutting the BGB (Olsson et al., 2010; Heubeck, 2019). Some of the best-preserved mounds are composed of Fe-Mg-Mn carbonates. Although ankerite and ferroan dolomite are often regarded as secondary in origin, a microbially mediated formation of these minerals cannot entirely be excluded.
The lithology, array of sedimentary structures (including common shale rip-up clasts, mudcracks, bidirectional paleocurrent indicators, shallow scouring), and depositional architecture are, in their combination, diagnostic of a tidal setting, mostly of a shallow-subtidal facies (Heubeck and Lowe, 1994; Homann et al., 2015; Heubeck et al., 2016). Paleoarchean tidal environments were probably poorly delineated and graded extensively into adjacent terrestrial and marine realms due to a lack of rooted vegetation, high tidal forcing, and their pre-plate tectonic setting. Aside from a clear southwestern provenance of most units, the lower 2000 m of Moodies strata in the studied Saddleback Syncline show no consistent stratigraphic lateral and vertical changes in depositional setting, partially owed to poor outcrop conditions (Heubeck and Lowe, 1994; Homann et al., 2015; Stutenbecker et al., 2019) but potentially also due to high sedimentation rates resulting in vertical stacking of facies.
Abiogenic or biogenic origin of the carbonate?
The regional context of the mound-bearing unit within the zone of hydrothermal silicification above a major Moodies-age mafic sill provides a strong argument for abiogenic mineral reactions associated with mound growth (Reimann et al., 2021). Abundant carbonate was likely transferred into solution by hydrothermal activity and metasomatism in the contact-metamorphic halo of the Lomati River Sill; it would readily have precipitated by CO2-degassing from pore waters upon reaching the sediment-water interface, in particular where flow was fast (e.g., Lowe, 1994; Grotzinger and Rothman, 1996; Pope and Grotzinger, 2000; Tobler et al., 2008).
To evaluate cases of proposed biogenicity, protocols (Buick et al., 1981; Cady et al., 2003; Brasier et al., 2004, 2006; Noffke, 2009; Schopf and Kudryatsev, 2012; McLoughlin, 2022) are commonly used because neither sedimentary context, paleoenvironment, mode of preservation, molecular-structural characteristics, carbonaceous composition, carbon isotopic composition, nor three-dimensional morphology alone are reliable indicators of biogenicity. The photic-zone facies, original carbonate composition, morphological complexity and variation, bulbous convex-up geometry, microscale internal lamination, lateral centimeter-spacing, apparent preferred vertical growth, and increasing thickness of laminae toward the top of the mounds are all features known from stromatolites (Lowe, 1994; Awramik and Grey, 2005; Awramik, 2006; Schopf et al., 2007; Altermann, 2008; Allwood et al., 2009; Petroff et al., 2010; Riding, 2011) and comparable to those described from the ca. 3.43 Ga Strelley Pool Formation (Allwood et al., 2009, 2010; Wacey, 2010) and the ca. 3.48 Ga Dresser Formation of Western Australia (Dunlop et al., 1978; Walter et al., 1980; Hickman-Lewis et al., 2023). The existence of abundant kerogenous laminations resembling modern cyanobacterial mats in over- and underlying sandstones makes the involvement of biogenic processes in the carbonate mounds plausible. The structures described here fall within some (abiogenic) definitions of stromatolites (e.g., Semikhatov et al., 1979; Hofmann, 1973 “an attached, laminated, lithified sedimentary growth structure, accretionary away from a point or limited surface of initiation”); in comparison to the bulk of stromatolitic structures (e.g., classifications of Riding, 1991, 2011; Grey and Awramik, 2020, Altermann, 2008), they are comparatively small and more irregular.
It remains unclear whether biogenic processes were also involved in the precipitation of carbonate on top of and within the fluid-escape structures. Microbial mounds growing on top of fluid-escape structures may have simply exploited an elevated position above a turbid boundary layer to conduct more efficient photosynthesis; alternatively, their formation may have been boosted by a thermally induced flux of dissolved carbonate and CO2 through fluid-escape conduits. Carbonate plugs now located along the fluid-escape conduits may have originally formed at the surface and were subsequently buried. Alternatively, if the mounds grew within the fluid-escape structures, either an abiogenic origin or chemosynthetic processes may have been involved. This is a wide field, including methanotrophy (possibly using waste O2 from nearby photosynthesizing microbial mats or using sulfate reduction coupled to anaerobic methane oxidation; Himmler et al., 2018), anoxygenic photosynthesis using H2S in coastal hydrothermal fields (Allen, 2016), fermentation, denitrification, and other processes; see Bosak (2011) for a review. An important unresolved question, however, is which commonly available oxidant could react with decaying organic matter or CH4 to precipitate carbonate because sea water sulfate, in the Phanerozoic widely used in the anaerobic oxidation of methane, is thought to have been low in Archean oceans. Bulk carbon isotope measurements on carbonate remnants yielded relatively homogenous δ13Ccarb values of +0.2‰ and δ18Ocarb values of -15.4‰ (n = 16; Homann et al., 2018). These are common values for Archean carbonates that rule out significant secondary alteration but do not provide strong evidence for a direct link between microbial activity and carbonate formation. In the absence of conclusive isotopic data, comparing process-simulating models of mound morphology, spacing and internal textures (Bosak et al., 2013) to the field evidence may be a promising avenue to constrain biological controls and possible metabolisms.
Implications for possible life on Mars
The documentation of numerous small, well-preserved, weathering-resistant, stromatolite-like carbonate mounds in a Paleoarchean, fully siliciclastic, medium- to high-energy tidal setting broadens the depositional facies to be targeted in the search for life on Mars (Cady et al., 2003; Hickman-Lewis et al., 2023). Carbonates have been shown to exist on Mars at the Phoenix and Perseverance landing sites (Boynton et al., 2009; Clavé et al., 2022); they are thought to have formed in a variety of settings (Scheller et al., 2021). The mounds described here are comparable in age, setting, and perhaps taphonomy to those expected on Mars in once-habitable Noachian strata; their centimeter-scale macro- and millimeter-scale microfabrics would likely be readily observable by rover instruments (Tarnas et al., 2021; Clavé et al., 2022) and be key targets for drilling and sampling (Scheller et al., 2021; Hickman-Lewis et al., 2023).
Conclusions
Field and geochemical evidence preserved in small calcareous mounds in the 3.2 Ga Moodies Group of the central BGB is sufficiently affected by alteration and metamorphism to prevent the unambiguous identification of abiogenic or biogenic processes responsible for their formation. Although microbial life flourished during the deposition of Moodies sediments, abiogenesis currently seems the more parsimonious hypothesis in the absence of positive evidence for biogenic precipitation, although these irregularly and delicately laminated fabrics do not closely resemble known inorganic precipitates. The structures may be “hybrid carbonates” (Riding, 2008; Riding and Virgone, 2020) that originated from the small-scale overlap of biological and abiological processes in space and time, presently commonly observed in evaporitic and hot-springs environments conducive to in situ mineralization. The occurrence of these stromatolite-like structures in a fully siliciclastic tidal setting widens search criteria in the search for life on Mars. Detailed geochemical research on unweathered material and on the alteration processes affecting the host rock, detailed field mapping to constrain depositional facies and processes, and high-precision age dating would advance our understanding of this Paleoarchean depositional and ecological setting and help disentangle abiogenic and biogenic processes contributing to the origin of these mounds.
Footnotes
Acknowledgments
Research was partially financed by DFG funding to C.H. (He 2418/22-1 and 2418/26-1). We thank Mark van Zuilen for the Raman map, Dora Paprika for assistance with field work, and Wlady Altermann and Robert Riding for advice in the field. SAPPI is thanked for permitting access to private forest roads, the Mpumalanga Tourism and Parks Agency for research permits in the Barberton-Makhonjwa Mountains World Heritage Site, and the Barberton townspeople for their hospitality. Comments and corrections by Robert Riding and two anonymous reviewers significantly improved the manuscript.