
Abstract
Since the Viking Labeled Release experiments were carried out on Mars in the 1970s, it has been evident that the martian surface regolith has a strong oxidizing capacity that can convert organic compounds into CO2 and probably water. While H2O2 was suggested originally for being the oxidizing agent responsible for the outcome of the Viking experiments, recent analyses of the martian regolith by the Phoenix lander and by consecutive missions point toward radiation-mediated decomposition products of perchlorate salts as the primary oxidant. In a series of experiments, we have shown that abrasion and triboelectric charging of basalt by simulated saltation could be an additional way of activating regolith. We have also shown that abraded basalt with a chemical composition close to that of martian regolith is toxic to several bacterial species and thus may affect the habitability of the martian surface. In the present study, we investigated the effect of the quantitatively most important minerals (olivine, augite, and plagioclase) and iron oxides (hematite, magnetite, and maghemite) on the survival of bacterial cells to elucidate whether a specific mineral that constitutes basalt is responsible for our observations. We observed that suspensions of iron-containing minerals olivine and augite in phosphate-buffered saline (1 × PBS) significantly reduce the number of surviving cells of our model organism Pseudomonas putida after 24 h of incubation. In contrast, the iron-free mineral plagioclase showed no effect. We also observed that suspending abraded olivine and augite in 1 × PBS led to a dramatic increase in pH compared to the pH of 1 × PBS alone. The sudden increase in pH caused by the presence of these minerals may partly explain the observed cytotoxicity. The cytotoxic effect of augite could be relieved when a strong buffer (20 × PBS) was used. In contrast, olivine, despite the stronger buffer, maintained its cytotoxicity. Iron oxides per se have no negative effect on the survival of our test organism. Overall, our experiments confirm the cytotoxicity of basalt and show that no single constituent mineral of the basalt can account for its toxicity. We could show that abraded iron-containing minerals (olivine and augite) change the pH of water when brought into suspension and thereby could affect the habitability of martian regolith.
Introduction
Since the first photos of Mars taken during fly-bys of the Mariner spacecrafts were transmitted to Earth, it was evident that the martian surface is a harsh environment, and all follow-up missions have provided strong indications that the martian surface is deprived of at least all complex life-forms. Until now no sign of life, neither extant nor extinct, has been found on the surface of the Red Planet. In the meantime, the factors that may affect organisms and/or their remains have been investigated with a primary focus on radiation (Hansen et al., 2005, 2009; Bak et al., 2022; Horne et al., 2022).
The Viking Labeled Release experiments were the first and until now the only in situ experiments that demonstrated that the martian regolith is very reactive (Levin and Straat, 1976; Klein, 1977; Apak, 2008), with a clear oxidizing capacity. As such, no radiation is needed to oxidize simple organic molecules on Mars when liquid water is present (Navarro-González et al., 2010; Bak et al., 2017a; Fox et al., 2019; Lewis et al., 2021). The reactivity was explained by accumulation of H2O2 that can be produced by (1) photochemical reactions in the atmosphere followed by deposition in the upper regolith strata (Encrenaz et al., 2008, 2012), (2) charging of dust during global dust storms and dust devils (Delory et al., 2006; Kok and Renno, 2009; Merrison et al., 2012), and (3) the fracturing of minerals caused by meteorite impacts that results in the formation of reactive surfaces (Fubini and Hubbard, 2003; Hurowitz et al., 2007).
The Phoenix lander mission from 2008 demonstrated the presence of large quantities of perchlorate salts in the regolith (Keller et al., 2006; Hecht et al., 2009; Clark and Kounaves, 2016). These salts have a high oxidizing capacity but only when they decompose and release oxygen at temperatures above 150°C (Glasner and Weidenfeld, 1952; Royle et al., 2017), which is not per se relevant to an explanation of the Viking results. However, Quinn et al. (2013) showed that Ca-perchlorate, when exposed to gamma rays, produces O2 that is trapped in the perchlorate salt, as well is chlorine dioxide (ClO2) and hypochlorite (ClO-). Adding a solution that was containing a mixture of radiolabeled organics equivalent to the mixture applied in the Viking Labeled Release experiment, they were able to reproduce the Viking results. They identified the hypochlorite that reacts with amino acids as the oxidant that would explain the outcome of the Viking Labeled Release experiment. They argue that their results provide good evidence for perchlorates irradiated by cosmic rays and solar energetic particles as the major source of oxidants in the martian regolith.
In a series of laboratory studies, our group has demonstrated that abrasion of minerals during saltation in a simulated martian atmosphere activates minerals by the formation of new surfaces (Bak et al., 2017a, 2022). On Mars, this mechanism could be relevant as the outcome of wind-driven erosion. By abrading islandic basalt with a composition that is comparable to martian basalt, we showed that the process produces precursors of H2O2 and hydroxyl radicals that lead to H2O2 formation after the abraded minerals were suspended in water (Bak et al., 2017a, 2022). We also showed that abraded basalt kills cells of the bacterial species Deinococcus radiodurans and Pseudomonas putida, as well as vegetative cells of the species Bacillus subtilis (Bak et al., 2019). The cytotoxic effect of abraded basalt is removed after washing the abraded basalt with MilliQ water and drying it overnight at 110°C. Hendrix et al. (2019) ground the planetary regolith analogs olivine, pyroxene minerals, including augite and plagioclase feldspars in a ball mill and showed also that the ground minerals produced reactive oxygen species (ROS), including hydroxyl radicals. They argued that their results should be considered when planning manned missions to the Moon and to Mars, as the finely comminuted minerals present a health risk for astronauts. A possible mechanism behind the toxicity of the abraded minerals could be lipid peroxidation of the cell membranes by Fenton-type reactions that are catalyzed by Fe3+ or Fe2+ ions that are present in the basalt, converting hydrogen peroxide into highly toxic hydroxyl free radicals (Haber and Weiss, 1934; Horwell et al. 2007).
In the current study, we investigated the role of iron by a series of experiments with the iron-containing minerals olivine and augite and the iron-free mineral plagioclase, three minerals that are common on Mars (Christensen et al., 2008; Bish et al., 2013). We investigated as well the effect of the iron oxides hematite, magnetite, and maghemite, which are responsible for the red color of the martian surface (Chevrier and Mathé, 2007). We also included quartz and basalt to reproduce the results reported by Bak et al. (2017b). All minerals were abraded for 63 days in a Mars-like atmosphere, and the effect on the survival of bacterial cells was investigated as described by Bak et al. (2019). As a test organism, we used Pseudomonas putida, a rod-shaped, motile, Gram-negative aerobic soil bacterium. We used Ps. putida, but not the more commonly used model microorganism D. radiodurans, because Bak et al. (2017b) showed that Ps. putida was more robust to the abraded basalt and gave results faster than D. radiodurans.
Material and Methods
Preparation of abraded silicate samples
To investigate the effect of abraded minerals on the survival of bacterial cells as a function of the mineral composition in general and the presence of iron in particular, three different minerals were chosen based on their respective abundance in the martian surface regolith (Bish et al., 2013). X-ray diffraction analysis of scooped samples gathered by the Curiosity rover from the Rocknest Aeolian Bedform in Gale Crater (Bish et al., 2013) showed that basaltic material was dominated by plagioclase (40.8%), olivine (22.4%), augite (14.6%), and pigeonite (13.8%). The three most abundant minerals suit perfectly well for testing the effect of iron as they represent two different iron-rich minerals, augite and olivine, and one iron-depleted mineral, plagioclase. In addition, we tested survivability in the presence of three iron oxides—hematite, maghemite, and magnetite—which we mixed into quartz sand that was abraded for 63 days. Martian regolith contains about 20% by weight Fe3+ in the form of iron oxides and iron oxyhydroxides. These are mainly responsible for the planet's red coloring, hence the nickname “the Red Planet.” The main phases observed are in the form of hematite, magnetite, and other minor components such as maghemite, ferrihydrite, and goethite (Chevrier and Mathé, 2007). We also included basalt and quartz with reference to Bak et al. (2017b). We tested the minerals in a non-abraded form as well as abraded minerals that were inactivated by a procedure described by Bak et al. (2017b).
The minerals plagioclase, olivine, and augite were obtained from Ward's Science Inc. (California, USA). Quartz sand was obtained from Merck (Cat. No. 1.07536). The basalt sample stems from Iceland (64°07'29.4″N, 21°43'24.6″W) and is very similar in composition to martian basalt (Bak et al., 2017a). The desired grain-size fraction of 0.125–1 mm was achieved by first crushing the minerals with a hydraulic press, and the acquired mineral powder was then sieved to extract the grain-size fraction. Thereafter, samples of 100 g were suspended in water to remove the very fine-grained material adhering to the larger particles. The water was stirred and removed by suction leaving behind the grains of the desired size. The wash was repeated until newly added water appeared clear after stirring. The samples were dried at 200°C overnight. On the following day, the samples were aliquoted into homogeneous 10 g samples using a Fritsch Rotary cone sample divider. Two samples of each mineral were used for mineralogical characterization by X-ray fluorescence analysis (see the Supplementary Material for data; Table S1).
Eleven samples of each mineral type were eroded by tumbling for 63 days as described by Bak et al. (2017a, 2017b). Briefly, quartz ampoules that are 20 cm long and 3 cm wide were filled with 10 g of sample material. The ampoules were evacuated to <0.12 mbar and filled with 8 mbar of a gas mix composed of 95% CO2 (>99.9% purity, AGA Denmark) and 5% of a custom-made mix (>99% purity, Air liquid, Denmark), giving a final composition similar to that of the martian atmosphere as measured by the Curiosity rover (Mahaffy et al., 2013). The ampoules were then sealed by melting the narrow top and, thereafter, placed in the tumbler and tumbled end over end for 63 days at 30 rpm to simulate wind-driven saltation of minerals on the martian surface (Bak et al. 2017a, 2017b). One ampoule containing each one of the minerals was opened after 63 days, and the content was used for grain-size distribution analysis and comparison of the grain-size distribution with the original material. The size distribution of the particles was measured with a Sympatec laser diffraction instrument with a 632.8 nm HeNe-laser and a Quixel wet dispersion system (see the Supplementary Material for data, Fig. S1). One ampoule of each mineral was inactivated by opening the ampoule, suspending the content in water, and drying it at 60°C following the procedure described by Bak et al. (2017b).
Preparation of Pseudomonas putida cultures
Pseudomonas putida mt-2 (DSM6125) was grown from a glycerol stock culture. The preparation of the cell suspensions and the exposure experiments were carried out according to the procedure described by Bak et al. (2017b) to allow for direct comparison with the results reported by Bak et al. (2017b).
The cultures were grown in 100 mL Erlenmeyer flasks containing 30 mL of liquid medium composed of 80% phosphate-buffered saline (PBS) (8 g NaCl, 0.2 g KCl, 1.44 g Na2HPO4, and 0.24 g KH2PO4 per liter, pH adjusted to 7.4) and 20% liquid lysogeny broth (LB) medium (10 g NaCl, 10 g tryptone (Fluka), and 5 g yeast extract (Merck) per liter, pH adjusted to 7). Lysogeny broth agar plates were made by adding 1.5% w/w of bacteriological agar to LB prior to autoclaving. All chemicals were of analytical grade and acquired from commercial suppliers. MilliQ water was used for all solutions. Cultures were harvested after 120 h of growth in the early stationary phase.
Prior to exposure to the eroded minerals, the cells were washed once to reduce the amounts of growth medium present in the cell suspensions. The cultures were centrifuged at 4700 × g for 5 min in 50 mL sterile Falcon tubes. The supernatant was discarded, and the pellet was resuspended in 30 mL 1 × PBS (pH 7.4).
Exposure-to-minerals experiments
Each of the exposure experiments was carried out as follows. Cell numbers in 1 × PBS cell suspensions were determined using a Neubauer counting chamber, and the cell number was thereafter adjusted to approximately 106 colony-forming units (CFUs) per milliliter by addition of a 1 × PBS solution. The cell suspensions were flushed with N2 (99.9% purity, AGA, Denmark) through sterile 0.2 μm filters to prevent contamination and transferred to a nitrogen gas–filled glovebox. The mineral-containing ampoules were opened in the glovebox, and the content was transferred to a sterile beaker. Three mineral samples of 1 g each were transferred from the beaker to sterile Falcon tubes. Thereafter, 2 mL of the cell suspension was transferred to the mineral-containing Falcon tubes. Cell suspensions in PBS buffer served as references and were used to determine the relative change in CFUs compared to cells that were in contact with minerals. Immediately after, dilution series were prepared from triplicate subsamples of the PBS cell suspension, and 100 μL from each dilution was transferred onto LB plates for later CFU counts. Subsamples of 100 μL from each Falcon tube containing eroded minerals were transferred to Eppendorf tubes at time 0, 1 h, 2.5 h, 5 h, and 24 h. The Eppendorf tubes were removed from the glovebox, and dilution series were prepared and plated immediately after. The plates were incubated at 30°C, and the CFUs on the plates were counted after 24 h. Experiments with inactivated minerals were carried out in the same way.
In addition, separate Falcon tubes with a similar mix of minerals in cell-free 1 × PBS buffer (start pH 7.4) were prepared to measure the change in pH upon addition of the mineral to 1 × PBS. To determine the effect of abrasion on pH, a suspension of non-abraded mineral was included for comparison. The solution remained stagnant after addition of the minerals until the end of the experiment, and the pH in the supernatant was measured 0, 1, 2.5, 5, and 24 h after addition of the mineral.
Exposure of Pseudomonas putida to abraded silicates in 20 × PBS
To determine the effect of the buffer concentration on bacterial survival, we also performed experiments using 20 times more concentrated PBS. This experiment was carried out in the same way as the previous experiment. A 20 × PBS solution without minerals served as control.
Exposure of Pseudomonas putida to abraded quartz mixed with Fe2+/Fe3+ iron oxides
To test the effect of iron oxides on bacterial survival, magnetite, maghemite, and hematite were added at a total 10 wt % to 1 g of quartz and mixed. The three iron oxides have been observed on Mars (Chevrier and Mathé, 2007). The experiments were carried out in a glovebox, and the number of CFUs was determined as described above.
Statistical analysis
In all cases, the average of three samples was calculated to estimate the standard deviation. To determine whether two treatments showed a statistically significant difference, a two-tailed t-test with samples of unequal variance was conducted. A significance value of <0.05 was used.
Results
Survival of Ps. putida cells in suspensions of abraded minerals
Exposure experiment with abraded basalt, olivine, augite, and plagioclase
Pseudomonas putida cells (approx. 106 cells per mL 1 × PBS buffer) were mixed either with abraded basalt, olivine, augite, or plagioclase, and the number of surviving cells was determined as CFUs on agar plates as a function of contact time with the respective mineral.
In the presence of abraded basalt, the percentage of CFUs of Ps. putida cells dropped significantly to 1 ± 0.3% of the starting concentration in suspension with activated basalt after 1 h (Fig. 1) compared to the 1 × PBS (pH 7.4) controls (p-value: 0.010) and the inactivated samples (p-value: 0.008). After 24 h, the percentage of CFUs dropped to 0.03 ± 0.01% of the starting concentration in suspension with activated basalt. There was no significant difference in the percentage of CFUs in the suspension with inactivated basalt compared to the 1 × PBS control at any of the sampling timepoints. After 24 h of incubation, they ended at 404 ± 17% and 311 ± 125% of the starting concentration, respectively, indicating growth during incubation in inactivated basalt and in mineral-free 1 × PBS.
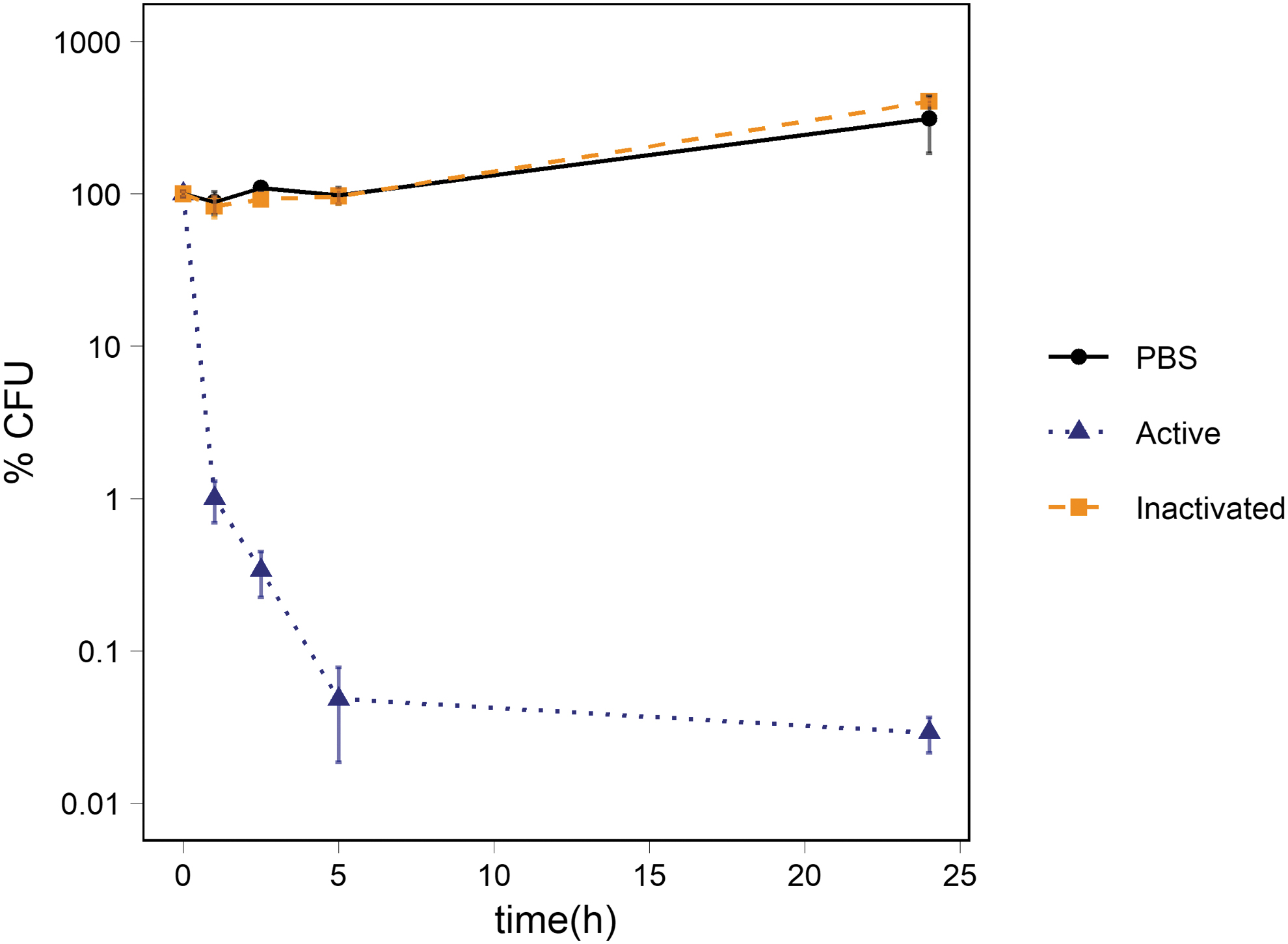
Survival of Ps. putida cells exposed to abraded basalt. The figure shows the percentage of CFUs in suspension of abraded basalt and of inactivated basalt. The average percentage of CFUs in the 1 × PBS control is also depicted. The number of cells was ∼106 CFU/mL at t0. The standard deviation of the three measurements is represented with error bars.
Measurements of pH showed a slight increase in the abraded basalt sample from pH 7.7 to pH ∼8 (Table 1) within 24 h. A negligible increase was observed in suspensions of inactivated basalt, reaching pH 7.7 after 24 h of incubation. There was no significant change in the pH over time in suspensions with the non-abraded basalt.
pH Measurements of Basalt Suspensions in 1 × PBS Buffer (pH 7.4) at Different Time Points
In the presence of abraded olivine, the percentage of CFUs of Ps. putida cells significantly dropped to 0.045% of the starting concentration after 1 h compared to the 1 × PBS control (p-value: 0.001) and to the inactivated samples (p-value: 0.001) (Fig. 2). There were no CFUs detectable at the remaining time points in the suspensions of activated olivine. There was no significant difference in CFUs in the suspensions of inactivated olivine compared to the 1 × PBS control samples at t1 and t2.5. At t5, the suspensions of inactive olivine showed a minor drop compared to the 1 × PBS control (p-value: 0.001), decreasing to 78 ± 10% of the starting concentration, which continued to 41 ± 14% of the starting concentration at t24 compared to the 1 × PBS control. After 24 h, the PBS control contained 153 ± 11% more CFUs relative to the starting concentration (Fig. 2).
Survival of Ps. putida cells exposed to abraded olivine. The figure shows the percentage of CFUs in suspensions of abraded olivine and inactivated olivine. The average percentage of CFUs in the 1 × PBS control is also depicted. The number of cells was ∼106 CFU/mL at t0. The standard deviation of three measurements is represented with error bars.
pH measurements showed a strong increase in pH in suspensions that contained the abraded olivine sample, reaching a final pH of 10.7 after 24 h (Table 2). A large increase in pH was also observed in the inactivated sample. However, starting at a lower value of pH 8.6, this sample only reached a pH of 9.6 after 24 h. The pH of non-abraded olivine increased slowly over the 24 h from pH 7.4 to pH 7.9.
pH Measurements of Olivine Suspensions in 1 × PBS Buffer (pH 7.4) at Different Time Points
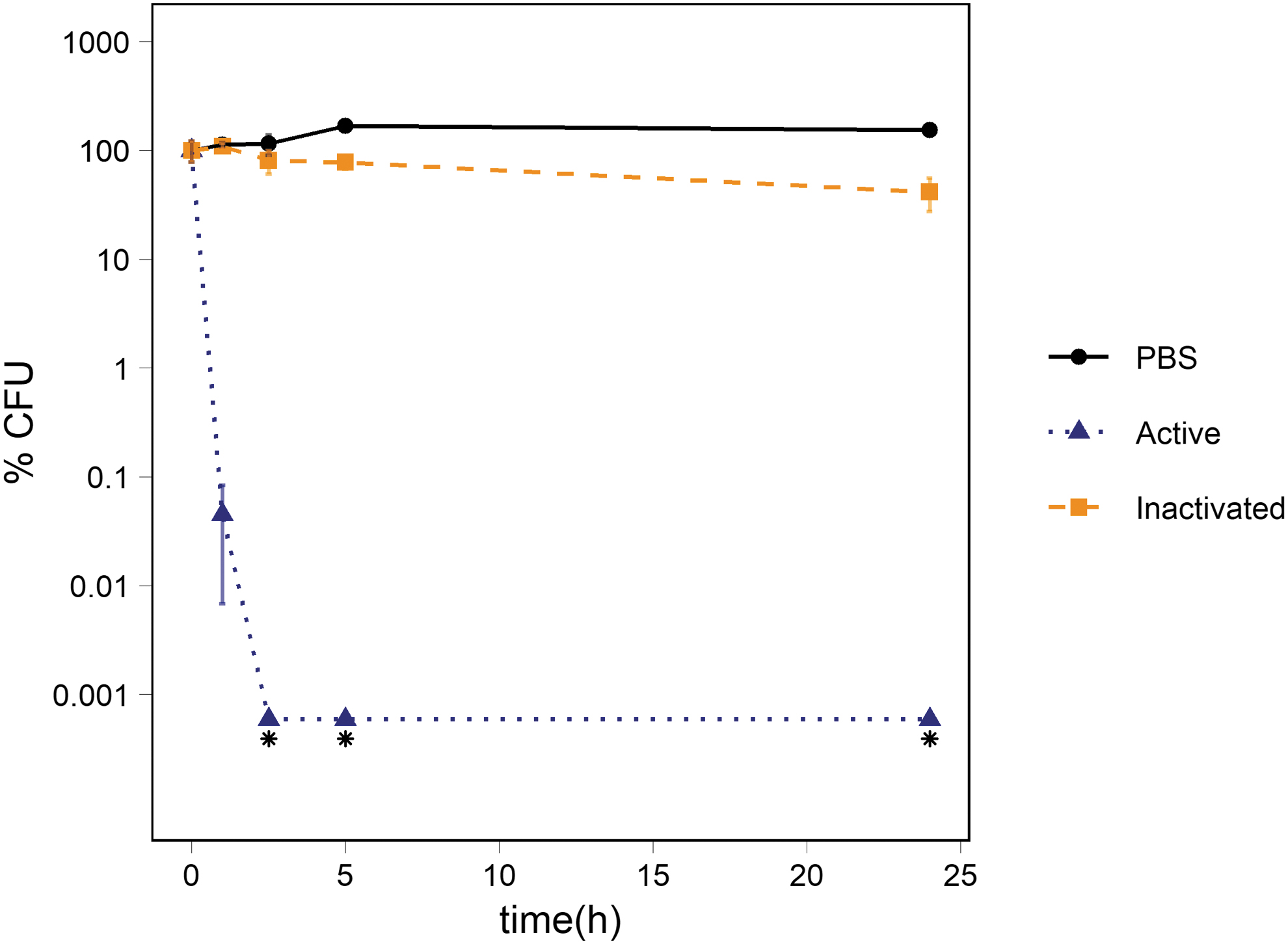
In the presence of abraded augite, there was no significant difference in the percentage of CFUs between the 1 × PBS control and the suspensions of activated and inactivated augite at t1, t2.5, and t5. However, at t5 there was a clear but statistically insignificant decrease in suspensions of activated (p-value: 0.056) and inactivated (p-value: 0.051) augite compared to the 1 × PBS control. The percentage of CFUs in suspension of activated augite dropped significantly between t5 and t24 compared to both the 1 × PBS control (p-value: 0.003) and the suspensions of inactivated augite (p-value: 0.004) reaching 1.2 ± 0.5% of the starting number of CFUs. The percentage of CFUs in 1 × PBS and in suspensions of inactivated augite ended at 166 ± 15% and 158 ± 17% of the starting percentages, respectively (Fig. 3).

Survival of Ps. putida cells exposed to abraded augite. The figure shows the percentage of CFUs in suspensions of abraded augite and inactivated augite. The average percentage of CFUs in the 1 × PBS control is also depicted. The number of cells was ∼106 CFU/mL at t0. The standard deviation of the three measurements is represented with error bars.
The pH measurements showed a drastic increase in the abraded augite sample ending at pH 9.8 (Table 3). In the inactivated augite sample, the pH reached 8.7 after 24 h. There were no notable changes in pH observed in the non-abraded sample during the 24 h of incubation.
pH Measurements of Augite Suspensions in 1 × PBS Buffer (pH 7.4) at Different Time Points
In the presence of plagioclase, there were no significant differences in the number of CFUs between the 1 × PBS control and the suspensions of activated and inactivated plagioclase at any time point. The suspensions of activated plagioclase showed a statistically non-significant increase compared to the 1 × PBS control (p-value: 0.08) and a significant increase compared to the suspension of inactivated plagioclase (p-value: 0.017). The percentage of CFUs compared to the starting concentration reached 175 ± 71%, 791 ± 146%, and 329 ± 67% for the 1 × PBS control, the suspensions of activated and inactivated plagioclase, respectively (Fig. 4).
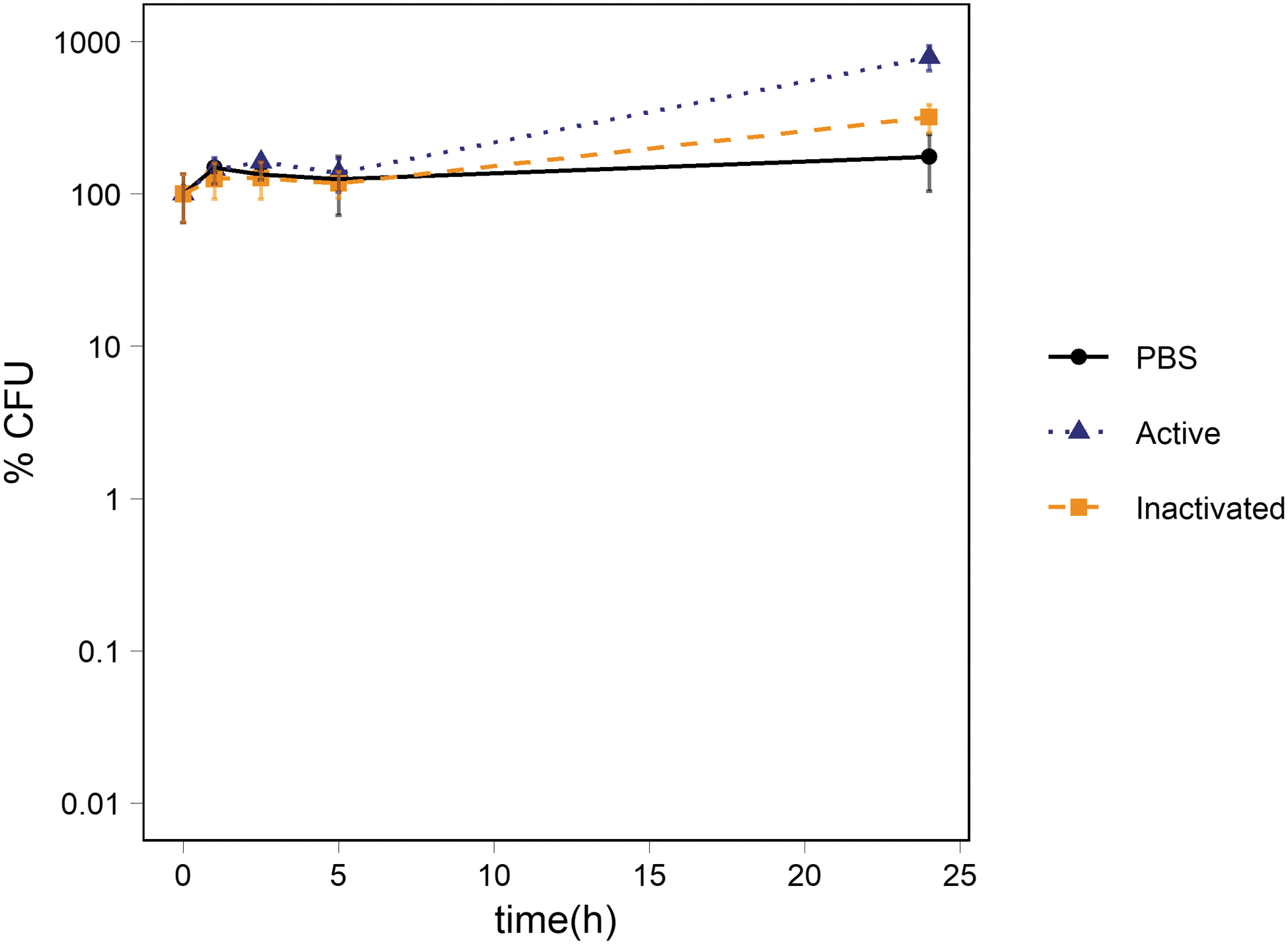
Survival of Ps. putida cells exposed to abraded plagioclase. The figure shows the percentage of CFUs in suspensions of activated and inactivated plagioclase. The average percentage of CFUs in the 1 × PBS control is also depicted. The number of cells was ∼106 CFU/mL at t0. The standard deviation of the three measurements is represented with error bars.
pH measurements showed a minor increase in pH in the suspensions of activated plagioclase, reaching pH 7.7 after 24 h (Table 4). A slight increase was also observed in suspensions of inactivated plagioclase reaching pH 7.5 after 24 h. In contrast, the pH dropped slightly in suspensions of non-abraded sample over the 24 h from pH 7.3 to pH 7.1.
pH Measurements of Plagioclase Suspensions in 1 × PBS Buffer (pH 7.4) at Different Time Points
To disentangle the effect of the mineral composition and pH on the survival of Ps. putida cells, the experiments were repeated in a stronger buffer solution (20 × PBS). To evaluate whether the strong buffer solution per se had an effect of the survival of Ps. putida cells, a 20 × PBS solution without mineral served as a control, and a 1 × PBS solution served as a positive control as it had demonstrated not to affect cell survival.
There was a significant difference between the percentage of CFUs in the 1 × PBS controls compared to the 20 × PBS at t1 (p-value: 0.002) and throughout the rest of the experiment, reaching 14 ± 3% of the original percentage of CFUs. The percentage of CFUs in suspension of activated augite in 20 × PBS followed the same trend as the strong PBS without any mineral added, and there was no significant difference in the percentage of CFUs in the 20 × PBS and the suspension of augite in 20 × PBS at any measured time point. The CFUs in suspensions of activated olivine in 20 × PBS dropped significantly compared to the 1 × PBS buffer at t1 (p-value: 0.003) and continued to decrease until t24 to 0.24 ± 0.17% of the original percentage of CFUs. At t5, the percentage of CFUs in the olivine suspensions dropped below both the percentage of CFUs in the augite suspensions (p-value: 0.002) and the percentage of CFUs in 20 × PBS (p-value: 0.001). The differences continued to increase at t24. The percentage of CFUs in suspension of activated basalt in 20 × PBS showed the steepest decline in percentage of CFUs of all tested suspensions between t0 to t1 (p-value: 0.0004, 1 × PBS) (p-value: 0.001, 20 × PBS) (p-value: 0.001, augite in 20 × PBS) (p-value: 0.001, olivine in 20 × PBS). This continued to increase until t5 to 1.0 ± 0.13%. At t24, the percentage of CFUs was below the detection limit of our method (Fig. 5).
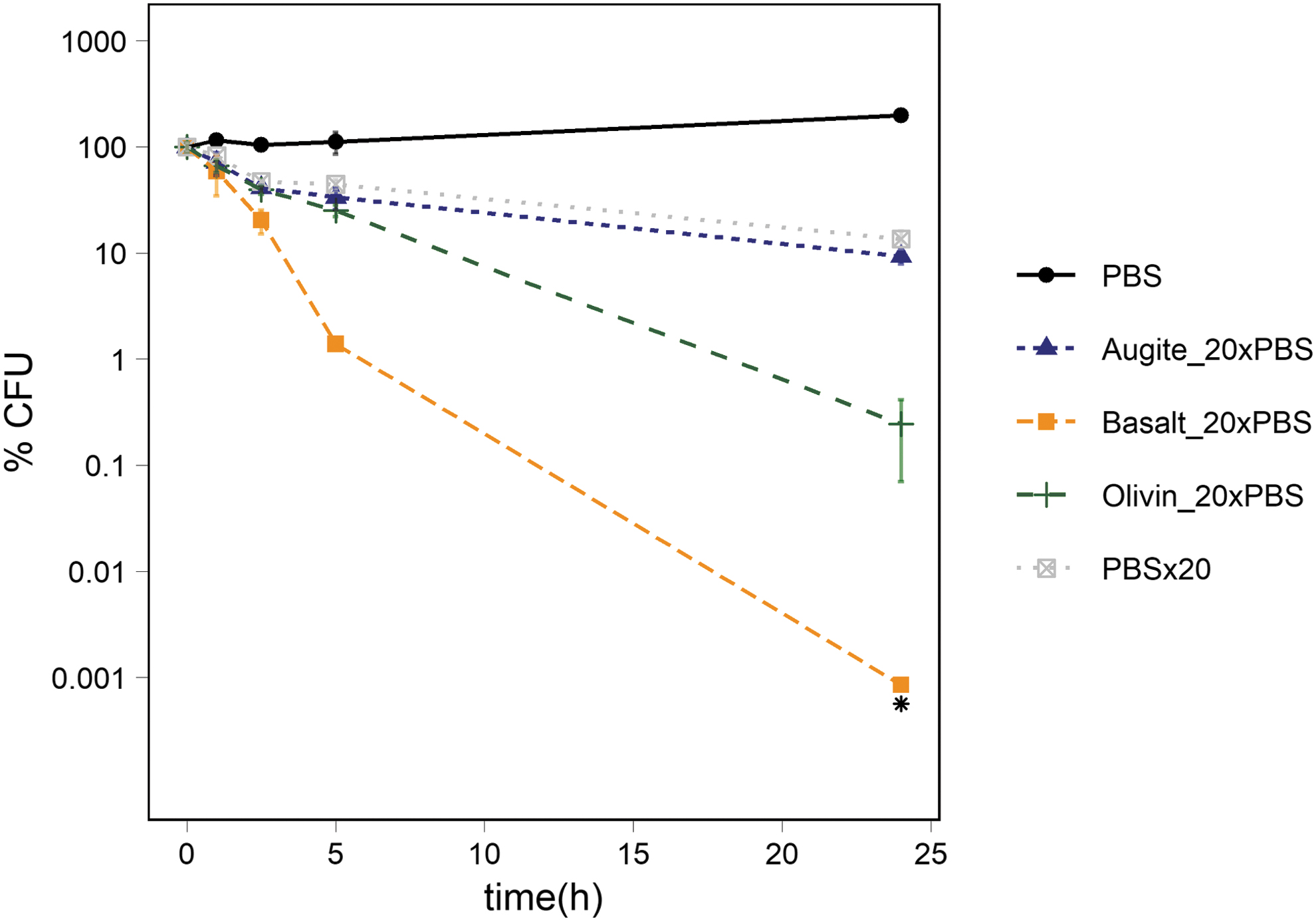
Survival of Ps. putida cells exposed to 20 × PBS alone or in suspension of activated augite, basalt, or olivine in 20 × PBS solution. The figure shows the percentage of CFUs in 1 × PBS, 20 × PBS, activated augite in 20 × PBS, activated olivine in 20 × PBS and activated basalt in 20 × PBS. The number of cells was ∼106 CFU/mL at t0. The standard deviation of the three measurements is represented with error bars.
pH measurements showed a drastic increase in pH in suspensions of activated olivine from pH 6.6 to 8.5 (Table 5). There was a minor increase in pH in the augite and the basalt suspensions in 20 × PBS throughout the experiment.
pH Values in Suspension of Augite, Basalt, and Olivine in 20 × PBS as a Function of Time
To evaluate whether iron oxides affected the survival of Ps. putida cells, 100 mg of either hematite, maghemite, or magnetite was mixed with 1 g of abraded quartz, and the mixture was suspended in 2 mL of a 1 × PBS Ps. putida suspension as described in Material and Methods. The percentage of CFUs in the 1 × PBS controls and iron-free quartz samples followed the same trend until t24, where the CFU percentages diverged significantly from each other (p-value: 0.002). In the quartz samples, the percentage of CFUs dropped to 75 ± 10%, and in the 1 × PBS control the percentage of CFUs increased to 202 ± 20% compared to t0. All iron oxide–containing experiments showed no significant difference compared to each other at any time point. They all showed a significant drop at t1 compared to both the 1 × PBS control and the suspension of iron-free quartz (quartz amended with hematite compared to a PBS control: p-value 0.004; compared to the quartz control: p-value 0.0002), (quartz amended with maghemite compared to a PBS control: p-value 0.01; compared to a quartz control: p-value 0.02), (quartz amended with magnetite compared to a PBS control: p-value 0.004 PBS; compared to a quartz control: p-value 0.03). At t2.5, the percentage of CFUs dropped to 18 ± 3% in suspension of quartz amended with hematite, to 19 ± 33% in suspensions of quartz amended with maghemite, and to 42 ± 16% in suspensions of quartz amended with magnetite compared to the t0 values. In each experiment, the percentage of CFUs increased to a level equal to the iron-free quartz samples and to the 1 × PBS control at t5, then followed the trend of the 1 × PBS control to t24 and reached 255 ± 43% in suspensions of quartz amended with hematite, 340 ± 61 in suspensions of quartz amended with maghemite, and 220 ± 24% in suspensions of quartz amended with magnetite (Fig. 6).
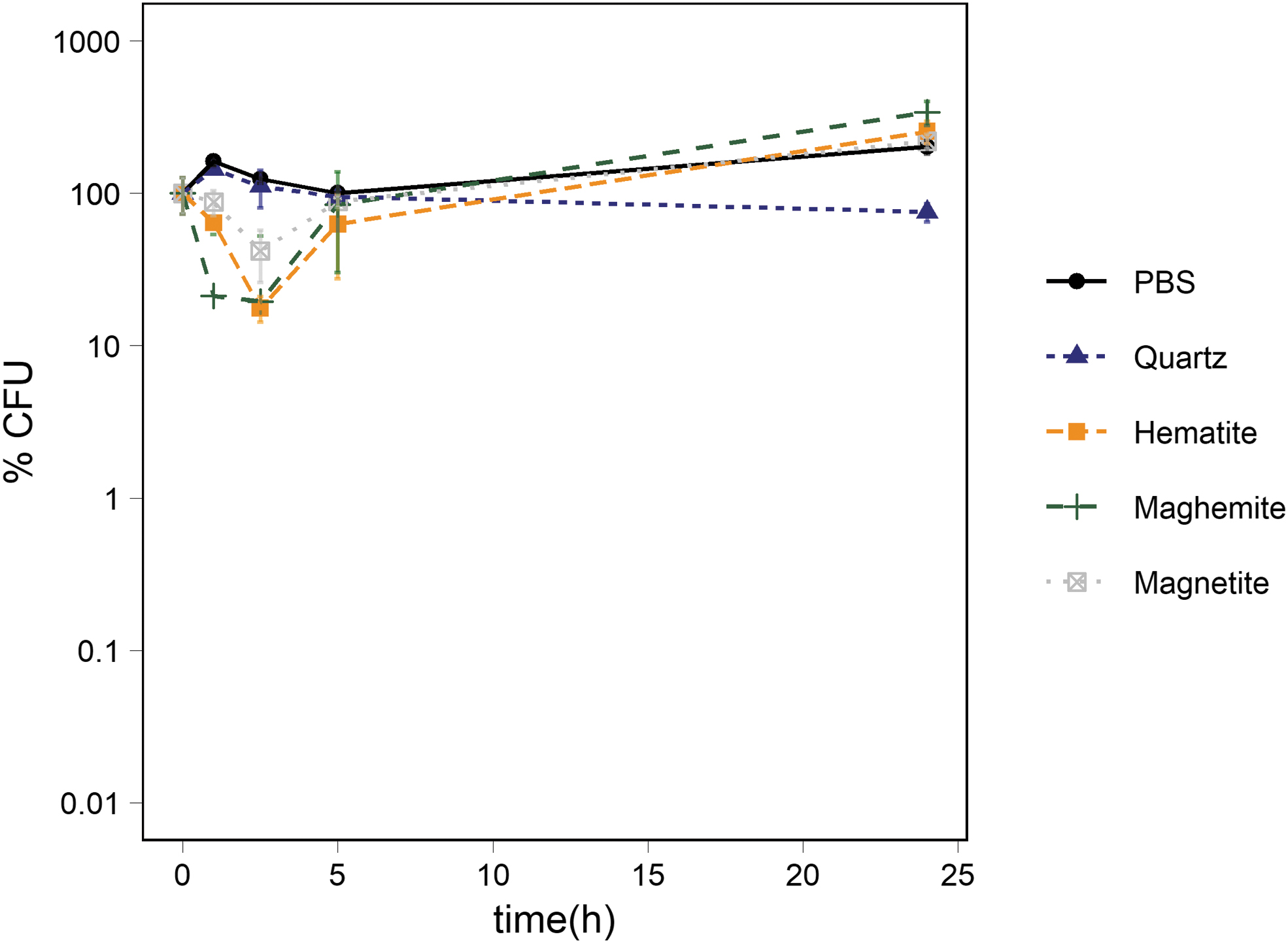
Survival of Ps. putida cells exposed to abraded quartz amended with 10 w/w % of the iron oxide hematite, maghemite, and magnetite, respectively. The figure shows the percentage of CFUs in suspensions of quartz, quartz-amended hematite, quartz amended with maghemite, and quartz abraded with magnetite. The average percentage of CFUs in the PBS control is also depicted. The number of cells was ∼106 CFU/mL at t0. The standard deviation of the three measurements is represented with error bars.
The pH was only measured at t0 (pH 7.3) and at the end of this experiment (t24). In all suspensions containing quartz and one of the iron oxides the pH decreased from pH 7.3 at t0 to ∼pH 6.8 at t24. In the suspensions of the iron-free quartz the pH only dropped slightly to pH 7.2 at t24.
Exposure experiments
Potential effect of iron oxides through Fenton-type chemistry on the survival of Ps. putida cells
We have previously observed that suspensions of abraded basalt can kill bacterial cells (Bak et al., 2017b). In that study, we hypothesized that Fenton-type chemistry producing ROS is responsible for the observed cytotoxicity. In this study, we investigated whether the individual minerals that constitute the basalt have cytotoxic effects per se. We examined the effect of abraded olivine, augite, and plagioclase which dominate the basaltic material on Mars (Bish et al., 2013). Of the tested minerals, olivine and augite contain iron while plagioclase does not (for details see the Supplementary Material Table S1). In addition, we investigated quartz supplemented with 10% w/w of magnetite, hematite, and maghemite, respectively, to test whether iron oxides mixed in a mineral matrix expressed cytotoxicity. In accordance with Bak et al. (2017b), we hypothesized that iron, both in the form of Fe2+ and Fe3+, could explain the cytotoxicity of basalt. Therefore, we assumed that olivine, augite, as well as iron oxide–amended quartz would kill cells as reported for basalt, while plagioclase would not. Our immediate observations agreed with this hypothesis, and we thus concluded that a Fenton-type mechanism could be responsible for the observed cytotoxicity. In agreement with our hypothesis, we observed that the relative number of CFUs decreases in the suspensions of hematite-, maghemite-, and magnetite-amended quartz but only during the first 2.5 h of the experiment. To our surprise, the number of CFUs then increased toward the end of the experiment and reached the same relative numbers as the 1 × PBS control. Apparently, Ps. putida cells that survive the initial phase of being in contact with iron oxide can proliferate even in a 1 × PBS solution without nutrients and energy sources. A similar observation was reported by Xu et al. (2012), studying the effects of several minerals including quartz on the survival of cells of Ps. aeruginosa strains. They explained the observation with adoption of a defense mechanism that protects the cells against the amended particles. They suggested that extracellular polymeric substance may shield the cells against the toxic minerals. This does not, however, explain why the percentage of CFUs increased again toward the end of their experiments. We suggest that the increase in CFUs in our study, as well as in the study by Xu et al. (2012), could be explained by a mechanism involving the use of polyhyroxyalkanoates (PHA). The ability to produce PHA has been reported in many different strains of Ps. putida including strain mt-2 and in Ps. aeruginosa (Rojas-Rosas et al., 2007; Follonier et al., 2011; Shahid et al., 2013). PHA functions as a storage molecule for carbon, and the synthesis of PHA is induced by shortage of either carbon or nitrogen in the surrounding environment (Shahid et al., 2013). In some cases, the dry weight of the cell contained up to 77% PHA (Shahid et al., 2013), and this could be the energy and carbon source for the observed cell growth in the 1 × PBS control and in some of the other suspensions, including the iron oxide–containing quartz suspension. The shift from dying to growing at t5 in the iron oxide–amended suspensions could be due to the production of siderophores (Schalk et al., 2020) by the surviving Ps. putia cells which complex the iron ions inside and outside the cell and thereby reduce their toxicity.
Potential effect of pH on the survival of Ps. putida cells
Measuring the pH in the mineral suspensions indicated that high pH in addition to ROS may have caused the observed cytotoxicity.
In suspensions of basalt in 1 × PBS, we only observed a slight increase in pH when comparing abraded material (pH 7.5–8.0) with material that was not abraded (pH 7.4). On the other hand, the pH in suspensions of abraded olivine increased dramatically and reached pH values between 10.1 and 10.7 when activated and between 8.6 and 9.6 when inactivated. The pH in suspensions of non-abraded olivine varied between pH 7.4 and 7.9 (Table 2). These observations indicate that Ps. putida has a significant pH tolerance, at least in the short term (24 h), and an upper limit for survival at around pH 10. This was supported by Reva et al. (2006), who reported growth of a related Ps. putida strain (Ps. putida KT2440) at pH 10. The strong increase in pH is probably due to an exchange of divalent ions such as Mg2+ and Ca2+ with protons at the mineral surfaces. While magnesium ions dominate in olivine (42% w/w of MgO), calcium ions are most prevalent in augite (23% w/w CaO). Plagioclase contains almost no MgO and only a minor amount of CaO (4% w/w of CaO). In plagioclase suspensions, we only observed minor changes in pH, even though it contains the largest amount of Na2O (10% w/w), which is a strongly basic oxide. We saw no effect on bacterial survival. Interestingly, basalt shows only a minor increase on pH even though it contains considerable amounts of Mg2+ (9% w/w of MgO) and Ca2+ (11% w/w of CaO) as well as Na2O (2% w/w). This may be explained by the presence of large amounts of Al2O3 (14% w/w) that, due to its amphoteric nature, may counteract the effect of the basic oxides. An even higher Al2O3 content (22% w/w) is found in plagioclase, which may counteract the basic effect of Na2O and consequently result in a milder increase in pH compared to olivine. The less pronounced increase in pH was observed in suspensions with non-abraded as well as inactivated minerals. Concerning the inactivated minerals, the reduced change in pH may be due to the inactivation procedure (Bak et al., 2017b). Here, the abraded material was washed and dried before it was added to the cell suspension. This pre-treatment may have removed some of the mineral's alkalinity and, thus, removed fewer protons from solution which resulted in a less pronounced increase in pH. Due to the strong pH effect, we were not able to evaluate whether ROS production could account for the cytotoxicity of the mineral in the 1 × PBS suspension. Therefore, we carried out experiments in 20 × PBS, which allowed us to stabilize the pH between 6.6 and 8.6, a pH that was tolerated by Ps. putida (Reva et al., 2006). We saw that the 20 × PBS affected the survival negatively per se compared to the 1 × PBS control. However, the cytotoxicity of augite in 1 × PBS was identical to the cytotoxicity of 20 × PBS, indicating that the decreased percentage of CFUs was most likely due to the increase in pH and not due to the iron-based Fenton-type chemistry. In contrast, olivine in 20 × PBS maintained its cytotoxicity compared to the 20 × PBS control, but the decline in the percentage of CFUs was less dramatic compared to olivine suspensions in 1 × PBS. This indicates that the cytotoxic effect of activated olivine could be due to a combination of increased pH and ROS production through Fenton-type chemistry. The cytotoxicity of basalt was slightly damped by a stronger buffered PBS solution. However, after 24 h no CFUs were seen on the agar plates inoculated with cells from suspensions of basalt in 20 × PBS.
5. Conclusions and Perspective s
Our study shows that basalt, activated through abrasion, expresses the strongest cytotoxic effect of the tested minerals. In contrast to its constituents, olivine, augite, and plagioclase, this effect is not due to pH effects but more likely to Fenton-type chemistry. In contrast to the iron oxides hematite, magnetite, and maghemite, cells of Ps. putida were not able to cope with the presence of activated basalt. The possible role of siderophores produced by Ps. putida in moderating the effect of iron oxides needs to be investigated. Future studies should include human cells or even animal models to investigate the possible health-threating effects of abraded and activated basalt on astronauts.
Funding
The research was financially supported by the Danish Council for Independent Research, Natural Sciences (ref. 09-066733) and the Villum Foundation (Villum Experiment grant No. 36213). Funding for the Stellar Astrophysics Centre is provided by The Danish National Research Foundation (Grant agreement no.: DNRF106).
Footnotes
Author Contributions
ENB, MB, KF, PN, JT, and SJKJ form a group dedicated to exploring triboelectric processes, f. ex. saltation on planetary bodies, like Mars. The current research is a spin-off of this endeavor. This study was designed by MKKS, ENB, KF, PN, and SJKJ. MKKS carried out the experiments. The manuscript was written by MKKS and KF with input from ENB, SJKJ, JT, MB, and PN.
Conflict of Interest
There is no conflict of interest.
Supplementary Material
Supplementary Information
Supplementary Table S1
Supplementary Figure S1
Associate Editor: Radu Popa
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.