
Abstract
The intense debate about the presence of methane in the martian atmosphere has stimulated the study of methanogenic species that are adapted to terrestrial habitats that resemble martian environments. We examined the environmental conditions, energy sources, and ecology of terrestrial methanogens that thrive in deep crystalline fractures, subsea hypersaline lakes, and subglacial water bodies, considered analogs of a hypothetical habitable martian subsurface. We combined this information with recent data on the distribution of buried water/ice and radiogenic elements on Mars, and with models of the subsurface thermal regime of this planet, we identified a 4.3–8.8 km-deep regolith habitat at the midlatitude location of Acidalia Planitia that might fit the requirements for hosting putative martian methanogens analogous to the methanogenic families, Methanosarcinaceae and Methanomicrobiaceae.
Introduction
One of the main aims of astrobiology is the detection of signatures of putative extraterrestrial life (e.g., Seager et al., 2012). Methane is one of the most studied biosignatures on Earth. Its detection, origin, and fate are at the center of intense research, particularly on Mars.
The debate over the presence of methane on Mars is long-standing (Herr and Pimentel, 1969; Sullivan, 1969) and continues to be fueled by the controversy surrounding its in situ detection reported in the Gale Crater (Webster et al., 2015), lack of its detection in the atmosphere using remote sensing techniques (Korablev et al., 2019), and efforts to reconcile the two approaches (Giuranna et al., 2019; Montmessin et al., 2021).
On Earth, archaea are organisms that release biotic methane as the primary metabolite of their metabolism. Eight types of methanogenic metabolism have been distinguished according to the use of different carbon sources and electron donors (Garcia et al., 2022); the three most relevant here are the following: Hydrogenotrophs, which extract energy from CO2, HCO3
−, or HCOO− (e− acceptor) and H2 (e− donor). Acetatotrophs, which extract energy from the degradation of acetate. Methylotrophs, which extract energy from the degradation of C1 organic compounds (formate, methanol, and methylamines, among others; Kurth et al., 2020).
Methylotrophs can use (or not use) H2 as the e− donor. This adds versatility to their metabolic capabilities (see Supplementary Appendix A1 for more details about biotic methanogenesis).
The discovery of a simplified ecosystem dominated by a hydrogenotrophic methanogen from the Methanobacteriaceae family in the hot groundwater of Lidy Hot Springs (Chapelle et al., 2002) has prompted speculation regarding the potential existence of a monospecific ecosystem.
Based on this, the methanogenic hydrogenotrophs have emerged as a candidate “model organism” for studying survival on Mars (Boston et al., 1992; Kral et al., 2010; Mickol et al., 2014) because they feed on simple inorganic molecules (i.e., H2 and CO2) that are expected to be available on the Red Planet. Since these molecules can have abiotic origins, this metabolic pathway could operate without mediation from any other biological metabolism. This conjecture has inspired numerous experiments that aim to test the ability of single methanogenic strains to thrive under the most extreme conditions of temperature, desiccation, salinity, pressure, pH, or radiation (see Supplementary Appendix A2 for an exhaustive list of experimental studies).
Although the findings at Lidy Hot Springs have been criticized (Nealson, 2005; Lin et al., 2006) and, to the best of our knowledge, have not yet been independently validated, it is essential to verify whether the conjecture regarding the existence of methanogenic monospecific communities has solid foundations or, conversely, if it is grounded on an anomalous observation.
The present-day martian surface and shallow subsurface are considered hostile for active Earth-like organisms (Hallsworth, 2021; but for a different opinion see Jones, 2018) due to the impact of high-energy radiation (galactic cosmic rays, solar UV, solar energetic proton radiation) and the extremely dry and cold conditions with extreme day–night temperature oscillations when compared with Earth’s standard environmental conditions (Butturini et al., 2020; Atri et al., 2022).
In contrast, the martian subsurface is believed to be more habitable than its surface (Westall et al., 2013; Tarnas et al., 2021) because it is sheltered from the impact of ionizing UV, X-ray, and particle radiation. In addition, subsurface temperatures are higher and exhibit smaller daily and seasonal fluctuations (Jones et al., 2011), likely increasing the probability of storing liquid water (Michalski et al., 2013). Finally, salts are believed to be relevant in the subsurface of Mars (Burt and Knauth, 2003), which opens up the possibility of liquid water at temperatures below zero. Therefore, the search for a habitable subsurface niche on Mars will involve the detection of anoxic habitats that have circulating liquid water and available energy and carbon sources such as those that support the growth of Earth-like microorganisms.
Three terrestrial subsurface environments have emerged as putative models for this type of a potentially habitable subsurface niche on Mars: Deep crystalline-bedrock continental fractures that host interstitial water (hereafter DCBF; Sherwood Lollar et al., 2007; Fernández-Remolar et al., 2008). Subglacial environments (hereafter SGL) that can host lentic waters and brines (Gaidos et al., 2004; Tung et al., 2005; Mikucki et al., 2015; Forte et al., 2016). Deep-sea hypersaline anoxic basins (hereafter DHAB; La Cono et al., 2019; Fisher et al., 2021).
Of course, DCBF, SGL, and DHAB habitats are coarse analogs of the (still poorly known) martian subsurface. On Earth, the ubiquitous and oxidant biosphere fueled by the Sun’s power has challenged the search for an optimal terrestrial analog of the anoxic martian subsurface. An “ideal” terrestrial analog should harbor a self-sufficient ecosystem with minimal hydrologic, geologic, atmospheric, and biological interactions with Earth’s surface (Lollar et al., 2021). However, only a few DCBF sites are likely to roughly fulfill these requirements for remoteness (Lin et al., 2006; Holland et al., 2013). Furthermore, most SGL and DHAB sites studied to date are not isolated from surface processes (see Supplementary Appendix A3 for more details).
In light of these limitations, SGL sites provide information about microbiomes at temperatures significantly lower than those reported at DCBF sites. On the contrary, microbiomes from DHAB sites (together with some SGL sites) thrive under conditions of extreme salinity. The debated SGL system at the martian south pole, modeled on the basis of radar profiles (Orosei et al., 2018) and anomalies in surface topography (Arnold et al., 2022), is an example of an SGL environment with cold saline fluids (although alternative explanations of radar and topography data exist; Lalich et al., 2024; Sori et al., 2024).
Given the considerations above, the topic of Mars’ subsurface habitability has led to various field studies focused on the DCBF, SGL, and DHAB habitats. By integrating the insights gained from such terrestrial environmental studies with a deeper understanding of the martian subsurface, crucial questions relevant to the potential for life on Mars can be addressed, including the following: Are methanogens significant in DCBF, SGL, and DHAB terrestrial habitats? Do the intrinsic environmental conditions of DCBF, SGL, and DHAB habitats shape the significance of subsurface methanogens and their metabolism? Is the almost “single-species” methanogenic ecosystem reported at Lidy Hot Springs representative of DCBF, SGL, or DHAB systems? How do the answers to the previous questions impact hypotheses regarding the viability of terrestrial methanogens in the subsurface of Mars? What environments in Mars’ subsurface could support Earth-like methanogens? What methanogenic pathways may prevail?
To address these questions, the present study followed two approaches: first, we compiled information about the presence of methanogens in DCBF, SGL, and DHAB systems (Section 3). Then we reviewed recent advances in understanding the geology of the martian subsurface to identify a region and subsurface that could host Earth-like methanogens (Section 4).
Several authors have reviewed separately the significance of microbiota in DCBF, SGL, and DHAB (Kieft, 2016; Achberger et al., 2017; Magnabosco et al., 2018; Merlino et al., 2018; Varrella et al., 2020; Fisher et al., 2021; Soares et al., 2023). However, to date, no known study has compared the composition of microbial communities across these three habitats in the context of Mars’ habitability. Therefore, we specifically review which methanogens have been reported in terrestrial subsurface ecosystems (Section 3.1), whether they coexist with other organisms (Section 3.2), and which energy sources they consume (Section 3.3).
To synthesize recent research advances on the geophysics of Mars’ lithosphere, we focus on (1) information about the availability of subsurface water or ice (Section 4.1); (2) information that provides clues about the possibility of producing abiotic molecular hydrogen, the simplest and most widely used energy source for methanogens (Section 4.2); and (3) information about how subsurface temperature increases with depth until it becomes tolerable for Earth-like methanogens (Section 4.3). Thus, we focus on water, energy sources, and temperature. In contrast, the availability of inorganic carbon, the main electron acceptor for methanogens, is not discussed. Recognizing the limited information about inorganic carbon in Mars’ subsurface (Ehlmann and Edwards, 2014; Wray et al., 2016; Jakosky and Edwards, 2018), we assume in this study that carbon is not a limiting factor.
Combining information from these two approaches provides the basis upon which we identified the location, extent, and depth of a potentially habitable Mars’ subsurface lithosphere that could host Earth-like methanogens and identified which methanogenic metabolism would adapt best to these putative habitats.
Data and Methods
This study combines the information provided by research focused on the microbiological and environmental characteristics of DCBF, DHAB, and SGL habitats as well as the geological, geochemical, and hydrological characterization of Mars’ subsurface. A summary of the methods used to obtain such information is provided below. Additional details are in Supplementary Appendix A4.
Microbial data source
The dataset included microbiological and environmental data from 79 sites (49 DCBF, 11 SGL, and 19 DHAB sites). The criteria for site selection are described in Supplementary Appendix A4. The compiled dataset and bibliographic sources are in Supplementary Appendices A5 and A6.
Salinity proxy
Taking into account the expected relevance of salinity in martian environments (Burt and Knauth, 2003), the molar ionic strength (I) of fluids was calculated. Supplementary Appendix A4 reports the method used to estimate the ionic strength at the selected study sites.
Mars’ subsurface habitats
Information about the presence of water, the availability of energy sources (for putative methanogens), and the geothermal gradients of Mars’ subsurface was collected from the literature.
Water/ice availability
Information about water or ice on Mars’ subsurface originates from in situ and remote measurements such as radar profiles obtained by the Zhurong rover; seismic data from the upper crust obtained by the InSight lander; images of the surface at the Phoenix and Viking 2 landing sites; and remote-sensing data provided by the Mars Odyssey Mission Gamma Ray Spectrometer (GRS), the Mars Advanced Radar for Subsurface and Ionospheric Sounding instrument (MARSIS), and the Mars Climate Sounder. Morgan et al. (2021) mapped the information obtained from multiple remote-sensing tools into contingency maps (Cv(z, long,lat)) that indicate the likelihood of the presence of ice at the surface (0–1 m depth, Cv(0, long,lat)) and of shallow buried ice (>5 m depth, Cv(5 long,lat)) from 60°S to 60°N.
Energy sources for methanogens
The focus is on the viability of abiotic production of H2 because it is a crucial energy source for terrestrial methanogens. Four main mechanisms are proposed to promote H2 accumulation in the martian subsurface: (1) H2 diffusion from the atmosphere to the subsurface dry regolith; (2) rock crushing after seismic movements; (3) water–mineral interactions such as serpentinization within crustal fractures; and (4) water radiolysis driven by radiogenic heat-producing elements (HPE: 232Th, 40K, and U).
Information related to (1) and (2) originates from theoretical considerations and models. The significance of items (3) and (4) has been explored by combining information provided by the Compact Reconnaissance Imaging Spectrometer (CRISM) and the GRS instruments with modelization. Among the HPEs, 232Th is the most important heat source, and its concentration ([Th](long,lat)) on Mars’ surface has been mapped across all longitudes and from 60°S to 60°N (Hahn et al., 2011).
Geothermal gradients
Temperature gradients in the subsurface martian upper crust can be estimated using the Fourier law and whatever information about average temperature and heat flow at the surface exists, together with clues about the thickness, densities, and thermal conductivities of the geologic structures expected in the martian upper crust. This information was compiled from the literature and is reported in Supplementary Appendix A4. The geothermal gradients were modeled in the region with the highest likelihood of hosting subsurface water and producing H2 together with the highest average surface temperature to locate the shallowest subsurface liquid water datum. At the target site, we estimated the depths at which temperatures are expected to be viable for methanogens according to the information collected from DCBF, SGL, and DHAB sites.
Methanogens from Terrestrial Subsurface Habitats
Methanogens in DHAB, DCBF, and SGL
Methanogens were reported at 66% of sites, and in 21% of cases they were considered a relevant component of the microbiome. Methanogens are ubiquitous in DHABs but were not reported at 45% of DCBF and SGL sites.
Regardless of their ecological significance, methanogens were reported to have a low relative abundance (less than <1% in most cases; Lavalleur and Colwell, 2013; Lopez-Fernandez et al., 2018; Nuppunen-Puputti et al., 2018; Achberger et al., 2016; Kalwasińska et al., 2020).
Methanobacteriaceae and Methanosarcinaceae were the most frequently reported families at DCBF and DHAB sites. At SGL sites, Methanomicrobiaceae and Methanosarcinaceae predominated over other families (Fig. 1a).

The pie charts show the percentage at which each family of methanogens
Hydrogenotrophics and methylotrophics were more frequently reported than acetatotrophics at the DCBF and DHAB sites, respectively. At SGL sites, the three methanogenic pathways were of similar significance (Fig. 1b).
Knowledge on rates of methanogenesis is scarce. Therefore, we know little about the methanogenic activity at the study sites. Despite the small amount of information available, the rate of methanogenesis appears to be higher at DHAB sites (in the range 0.5–85 µM/d) than at DCBF (<2.2 µM/d) and SGL sites (<0.45 µM/d; see Supplementary Appendix A6 for more information).
Methanogens have been reported in the temperature range −17.5 to 82°C (Fig. 2a); the Methanosarcinaceae and Methanomicrobiaceae are the most flexible families and are most frequently reported in cold waters (<15°C). In contrast, Methanobacteriaceae are typically reported in temperate-hot (T >15°C) fluids. The most significant exception is Tarn Flat interglacial organic-rich brine (temperature has not been reported but is likely below 0°C), which hosts a highly diverse methanogenic community, including Methanobacteriaceae (Papale et al., 2019).

Range of environmental conditions reported at the compiled DCBF, SGL, and DHAB sites. Colors indicate the family detected at each site (only the three most frequent families are shown): red dots = Methanobacteriaceae; blue dots = Methanosarcinaceae; green dots = Methanomicrobiaceae; black dots = not detected. The names of a few sites are provided. The data used to generate the plots are in Supplementary Appendix A5.
Methanogens have been reported to be active up to salinities of I∼9 M (Borin et al., 2009; Yakimov et al., 2013; Guan et al., 2015). In contrast, Telling et al. (2018) reported a maximum salinity tolerance threshold for the active microbiome in DCBF of I∼M (Fig. 3).
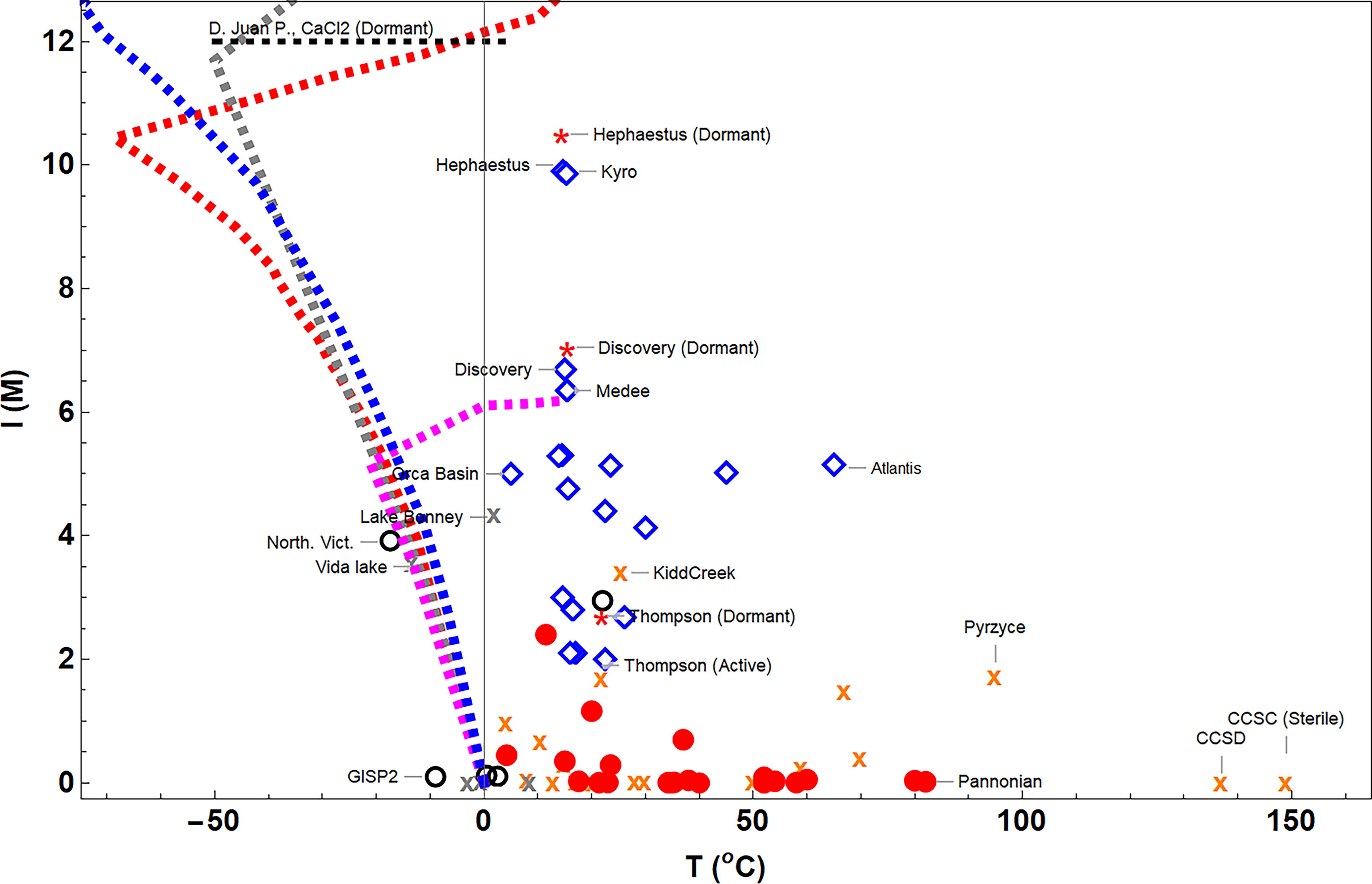
Relationship between salinity (in terms of ionic strength) versus temperature at the study sites. DCBF: red dots; SGL: open black circles; DHAB: blue rhombuses. Crosses indicate sites where the presence of methanogens has not been reported. The names of a few emblematic sites are shown. Dotted lines show the phase diagrams of NaCl (magenta dashed line), Mg(ClO4)2 (red dashed line), Ca(ClO4)2 (blue dashed line), and CaCl2 (gray dashed line). The horizontal segmented black line refers to the range of temperatures reported in the hypersaline Don Juan Pond in Antarctica (a CaCl2 brine apparently devoid of active life in the McMurdo dry valley; Oren, 2013).
DHABs are the most hypersaline subsurface systems, yet brines accumulated at some SGL sites (Lakes Vida, Vanda, and Bonney and endoglacial Antarctic brines of Northern Victoria land and Tarn Flat) and at some DCBF sites (Kidd Creek, Thompson mine, and Soudan iron mine). Methanosarcinaceae is the most salt-tolerant family. Methanobacteriaceae, Methanococcaceae, and Methanotrichaceae are usually linked to fresh and subsaline habitats (i.e., I < 2 M). Methanomicrobiaceae have been reported in brines with an ionic strength up to 4.5 M (Fig. 2b).
Methylotrophs typically predominate in brines with I > 4 M, whereas the hydrogenotrophs preferentially thrive under low salinities (Fig. 2b). Nevertheless, exceptions exist, and the below-zero temperature brines in the Northern Victoria Land endoglacial SGL host a strictly hydrogenotrophic Methanomicrobiaceae (Guglielmin et al., 2023).
The impact of sulfate overlaps with that of salinity: Methanobacteriaceae are typically constrained in water with [SO4 2−] < 2 mM. In contrast, Methanosarcinaceae are much more tolerant and are reported to survive in up to 0.4 M of sulfate (Fig. 2c).
Methanogens have been reported in the pH range from 4.5 to 11.3. Methanosarcinaceae and Methanomicrobiaceae are preferentially reported in neutral and acid fluids, whereas Methanobacteriaceae have been reported in water at pH > 6.5 (up to pH∼11.3 at Samail Ophiolite; Rempfert et al., 2017) (Fig. 2d). Hyperalkaline fluids host methanogens at the Samail Ophiolite site (Fones et al., 2019; Kraus et al., 2021; Nothaft et al., 2021) but have not been reported at the Coast Range Ophiolite (Twing et al., 2017) or Cabeço de Vide (Tiago and Veríssimo, 2013) sites.
With the exception of the Lidy Hot Springs site (Chapelle et al., 2002), methanogens, when present, coexist with other organisms. DCBF, SGL, and DHAB host taxonomic and metabolically diverse microbial communities that include bacteria, archaea, viruses (Miettinen et al., 2015; Mikucki et al., 2015; Holmfeldt et al., 2021; Nuppunen-Puputti et al., 2020), and even eukaryotes (Epova et al., 2022; Guglielmin et al., 2023). Bacteria typically predominate over archaea in most habitats (Murray et al., 2012; Guan et al., 2015; Magnabosco et al., 2018). Yet, in addition to Lidy Hot Springs, a notable abundance of archaea (always coexisting with bacteria) have been reported in Olkiluoto (Bomberg et al., 2016), the Tau Tona gold mine (South Africa; Magnabosco et al., 2016), and in some Mediterranean deep-sea brines (Urania, Bannock and Atlante; van der Wielen et al., 2005).
Besides methanogenesis, a plethora of autotrophic and heterotrophic metabolic pathways has been reported (for a list, see Supplementary Appendix A7). The core metabolisms are probably those linked to sulfur cycling (van der Wielen et al., 2005; Telling et al., 2018; Lollar et al., 2019).
Methanogens coexist with organisms that, a priori, compete for the same resources (mainly H2). Thus, methanogenic hydrogenotrophs are not outcompeted by sulfate/sulfur autotrophic reducers (Lin et al., 2006; Plugge et al., 2011; Lavalleur and Colwell, 2013; Magnabosco et al., 2016; Chiriac et al., 2018; Sanz et al., 2021) and do not exclude the bacterial homoacetogens (Kotsyurbenko et al., 2001; Rempfert et al., 2017; Lopez-Fernandez et al., 2018). Finally, syntrophic bacteria-methanogen consortia are frequently reported at DHAB (La Cono et al., 2015), DCBF (Moser et al., 2005; Osburn et al., 2014; Lau et al., 2016; Sheik et al., 2021), and SGL (Papale et al., 2019) sites.
Energetic resources for methanogens
Information about H2 was available for 29 sites. In most cases, the concentration was higher than 1 nM, the minimal reported H2 concentration that enables hydrogenotropic methanogenesis (Heuer et al., 2009). Most of these sites are DCBF sites, except for the two DHAB sites in the Gulf of Mexico and one SGL site (Lake Vida). Methanogens were not reported at 9 of these 29 sites (Supplementary Appendix A4). As well as hydrogen, concentrations of formate and acetate were reported at 22 sites (Supplementary Appendix A6). Methanogens were not reported at 6 of these 22 sites. Therefore, the presence of H2, formate, or acetate does not guarantee the proliferation of methanogens.
At DCBF sites, the presence of H2 is attributed to water–rock interactions (serpentinization and rock friction) and water radiolysis. Information about the availability of H2 is lacking for most DHAB sites. However, its availability is plausible at those DHAB sites in contact with deep gas venting such as in the Gulf of Mexico brines and some Red Sea brines (Merlino et al., 2018). Information about H2 at SGL sites is scarce. However, silicate–water laboratory experiments suggest that abiotic H2 production in glacial habitats might be significant (Samarkin et al., 2010; Telling et al., 2015; Gill Olivas, 2019).
The detection of the hydrogentrophic Methanomicrobiaceae in the SGL endoglacial brines indicates that H2 should be available. The Mg/K ratios in these brines and seawater are identical (Guglielmin et al., 2023), which suggests minimal rock weathering. Thus, the production of abiotic H2 through water–rock fractioning appears improbable. H2 is likely of biotic origin from the fermentation of organic matter and acetate oxidation. Indeed, the concentration of dissolved organic carbon is extremely high at these SGL sites (up to 7 mM; Papale et al., 2019; Fig. 2e).
The occurrence of acetate and formate at deep DCBF sites has been attributed to abiotic processes related to the reduction of dissolved inorganic carbon with H2 previously released after water radiolysis (Lollar et al., 2021) or serpentinization (Fones et al., 2019).
Water radiolysis also generates strong oxidants such as OH•, O2, H2O2, and O−, yet the impact of these agents on the endolithic biota remains unknown. Furthermore, these oxidants can react with reduced elements from minerals (such as pyrite) or with solutes (such as NH4 +) and produce SO4 2−, Fe+3, or NO2 −/NO3 − that can fuel other autotrophic metabolisms (Lefticariu et al., 2010; Silver et al., 2012), which, in turn, might compete with methanogens for H2.
Potential Habitats in Mars’ Subsurface
Water/ice availability
Phoenix and Viking 2 revealed that a very shallow (few centimeters in depth) subsurface ice is widespread and stable at latitudes higher than 45°N of the northern hemisphere (Mellon et al., 2009 and Mellon & Sizemore, 2022) that likely extends to several meters in depth (Putzig et al., 2014). In contrast, Zhurong and InSight indicated that, at low latitudes (<25°N) of the lowlands, the shallow and deep crustal regolith is apparently dry with weak evidence of the presence of ice or liquid water (Manga and Wright, 2021; Li et al., 2022; Wright et al., 2022). Maps of the likelihood of ice (Morgan et al., 2021) show the widespread presence of near-surface ice at latitudes higher than 60°. However, buried cryosphere (>5 m depth) is relatively likely even at latitudes down to 30°N in the northern hemisphere (Putzig et al., 2023; Fig. 3a). Similarly, Piqueux et al. (2019) estimated that shallow buried ice (at less than 1 m depth) is widespread at latitudes as low as 40°N. Finally, interpretation of MARSIS echoes has revealed the location of putative SGL-layered deep brine deposits at the south pole (Lauro et al., 2022; but for a different interpretation of MARSIS echoes see Lalich et al., 2024) and putative deep ice-rich deposits in the Medusae Fossae Formation, at the martian equator, below a 300–600-m-thick dry upper regolith (Watters et al., 2024). This latter discovery suggests that there might be subsurface ice at the present-day martian equator; this diverges somewhat from conclusions based on InSight data.
Availability of energy sources
In the present-day martian atmosphere, molecular hydrogen can be released after photochemical reactions. Sholes et al. (2019) modeled the possibility of passive diffusion of atmospheric H2 through the regolith and the possibility that it could sustain hydrogenotrophic methanogenesis in a subsurface connected to the atmosphere. A priori, this mechanism should be effective at low latitudes (<30°) where the upper subsurface is highly likely to be dry (i.e., ice-free, Fig. 4a), which allows the atmospheric gases to diffuse through the regolith interstices. Despite the existence of this theoretical approach, empirical evidence that provides clues as to the relevance of this process remains elusive, to the best of our knowledge. This leads to the conclusion that, even if the diffusion of photo-produced H2 in the atmosphere turns out to be possible, it would occur only in subsurface habitats that are intrinsically uninhabitable because they do not contain water. On Earth, friction, fracturing, and crushing of hydrated minerals or the reaction of silicon radicals with water (Kita et al., 1982) as a consequence of seismic movements (or meteoritic impacts) promote the liberation of seismogenic hydrogen. McMahon et al. (2016) hypothesized that seismogenic hydrogen in the martian upper crust could sustain hydrogenotrophic methanogenesis. This mechanism is expected to be activated by seismic events. The InSight mission demonstrated that Mars is seismically active, especially at Cerberus Fossae and along or north of the dichotomy between the southern highlands and northern lowlands, especially at the Tharsis Plateau (Ceylan et al., 2023). However, to date, there is a large uncertainty about the presence of subsurface water/ice in these regions (Burr et al, 2002; Morgan et al., 2021; Fig. 3a). Consequently, the current information does not strengthen the feasibility of H2 production from martian present-day seismic activity.
Serpentinization is a low-temperature water–rock process that produces hydrogen on Earth’s crust. Amador et al. (2018) concluded that clues that would indicate serpentine systems are rare on Mars’ surface. Indeed, it is only in the eastern portion of Nili Fossae that the detection of serpentine together with other related minerals (talc and saponite) clearly coincides with the detection of a high concentration of olivine-rich basalt outcrops to make past serpentinization in this area plausible. Although water–rock processes in the present-day martian crust cannot be ruled out (Vance and Melwani Daswani, 2020), information provided by CRISM does not support globally widespread serpentinization in early Mars when the planet was geologically more active and does not provide any indication of active serpentinization on present-day Mars. In sum, the available information suggests that a significant H2 yield from serpentinization in present-day Mars’ subsurface is unlikely.
In a water/ice-rich subsurface, radiogenic elements (HPE) fuel water radiolysis with the release of H2 (among other molecules) that might support a hydrogenotrophic community (Dzaugis et al., 2018). According to information provided by GRS (Hahn et al., 2011), the concentration of Th on Mars’ surface is approximately one-tenth of that reported in the terrestrial crust (Lin et al., 2005) but is in the same range as that reported in subsea basalts (Dzaugis et al., 2016).
The HPE concentration in the subsurface is unknown, but seismic data from InSight suggest that the martian crust is enriched in radiogenic elements (Knapmeyer-Endrun et al., 2021). Assuming that the abundance of radiogenic elements in the subsurface is the same as that detected at the surface (Hahn et al., 2011), the radiogenic heat production of the present-day martian crust would range between 2.5 and 7.5 × 10−5 µW/kg and would correlate with the abundance of Th, the largest heat source (Hahn et al., 2011). If interstitial water is in contact with such a radiogenic flux, H2 release is plausible (Dzaugis et al., 2018).
The size of the void volume in sediments and rock fractures influences the efficiency of water radiolysis. Indeed, H2 production is expected to be higher in low-porosity sediments (<5%) and small fractures (<1 µM width; Dzaugis et al., 2018). It is expected that the porosity of the first 4 km of the martian crust could range between 10% and 63%, with the highest values corresponding to the upper centimeters of the regolith (Clifford et al., 2010; Goossens et al., 2017; Michalski et al., 2018; Grott et al., 2021; Kilburn et al., 2022). Thus, although the porosity of the martian regolith of the upper crust is currently poorly constrained, radiolytic H2 production is likely to be more efficient in deeper environments with lower porosity and fracturing compared with the shallow subsurface.
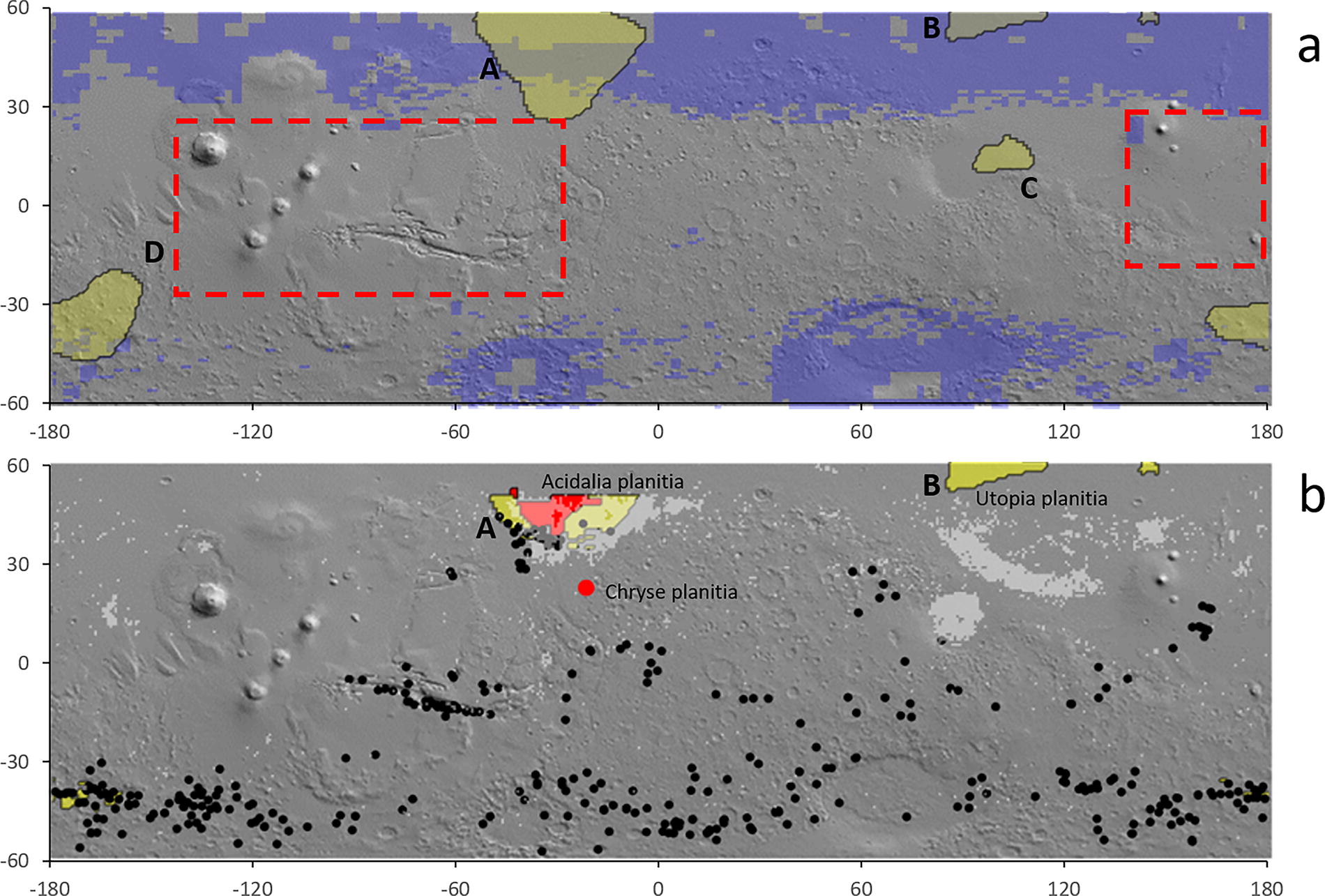
According to Hahn et al. (2011), there are four regions on Mars’ surface with a Th abundance higher than the 95th percentile (Fig. 4a, labeled A to D). Areas A (the largest one) and B are at midhigh latitudes (from 30°N to 60°N of the northern hemisphere). Area C (Hephaestus Rupes) is at a lower latitude (20°N) close to the Zhurong landing site. Finally, area D coincides with the Eridania Region, a zone that could have hosted a radiogenic hydrothermal system in the Noachian (Ojha et al., 2021).
We intersected the map of the ice contingency at a depth >5 m Cv(5,long.lat) (Morgan et al., 2021) with that of the Th abundance ([Th](long,lat)) to identify regions with the highest chance of promoting water radiolysis in the present-day martian subsurface. Therefore, this intersection helps identify regions with the highest likelihood of hosting subsurface Earth-like methanogens. Locations at which Cv(5 long,lat) and [Th](long,lat) values are higher than the 95th percentile are selected. This intersection reveals that the areas labeled as A and B in Fig. 4 have the highest probability to store a subsurface buried/shallow water/ice layer together with a high availability of HPE. In contrast, areas C and D were removed from the model because evidence of the presence of subsurface water/ice is currently weak (Fig. 4b). It is noteworthy that, according to planetary-scale crustal models (Parro et al., 2017), areas A and B are likely to experience low heat flow values, both at the surface and the base of the crust, which implies a deeper depth for the 0°C isotherm.
The last step involved calculating the geothermal gradient at one location within the selected areas A and B to find the depth at which the minimum temperature viable for terrestrial methanogens (i.e., −17.5°C) and the melting temperature of ice are exceeded.
Within the two areas, the most robust location is in the southern Acidalia Planitia (35°N, 30°W, red area at Fig. 4b), in contact with Chryse Planitia, with an annual average temperature of −58°C (according to the Mars climate database). This average temperature is higher than that estimated at the rest of the sites located inside areas A and B, because it is at the lowest latitude.
According to Parro et al. (2017), the heat flow at the surface of this location is approximately 0.016 W/m2. To constrain the subsurface geology and thermal conductivities, we adopted the information derived by the Zhurong rover and especially by the InSight lander. The Zhurong landing site and the Acidalia Planitia are within the limits of a putative ancient ocean in the northern hemisphere (Di Achille and Hynek, 2010), in the same geological unit (Late Hesperian lowlands unit, Tanaka et al., 2014), with a low dielectric constant that indicates the presence of low-density sediments or deposits of ground ice or both (Mouginot et al., 2012).
Radar reflections detected by Zhurong allow the identification of three main structural sedimentary layers below the landing site (Li et al., 2022): a regolith top layer probably of fine-grained sediments of approximately 10 m in depth; a second layer that extends up to 30 m in depth, mainly composed of unevenly distributed small boulders/cobbles; and a third layer that extends down to 80 m below the surface where large, clast-supported boulders are distributed more evenly. The deeper subsurface layers remain unconstrained at the Zhurong site. However, the InSight seismograph detected structural discontinuities at 2, 8–11, and 20 km below the surface (Knapmeyer-Endrun et al., 2021; Shi et al., 2023). The 2-km-deep discontinuity has been interpreted as a structural transition between a highly fractured basaltic layer (called a “megaregolith”) and a more coherent and less fractured layer (Carrasco et al., 2023; Shi et al., 2023). The discontinuity detected at 8–11 km deep has been attributed to the presence of rigid flood basalt (Xu et al., 2023) putatively constituted by an unfractured plagioclase feldspar rock (Kilburn et al., 2022). The boundary between the crust and the uppermost mantle is between 20 and 39 km (Knapmeyer-Endrun et al., 2021; Schimmel et al., 2021). Importantly, the InSight seismograph indicated the existence of a poorly cemented upper crust (Wright et al., 2022), which suggests that the significance of salts at subsurface at low- and midlatitudes of northern lowlands is doubtful. Notably, salts also seem to be in low abundance in Jezero Crater (Wiens et al., 2022), which is also located at low latitudes in the northern hemisphere (18.4°N; 77.6°E).
The thermal conductivity of the dry martian upper low-density regolith is constrained between 0.085 and 0.039 W/(m·K) (Zent et al., 2010; Grott et al., 2021). Li et al. (2022) modeled the temperature profile down to a depth of 140 m at the Zhurong site using a constant thermal conductivity of 0.8 W/(m·K) (as suggested by Egea-Gonzalez et al., 2021) regardless of the lithological vertical heterogeneity described above. Instead, to model the steady-state temperature profiles in the Acidalia Planitia subsurface, we changed the substrate’s thermal conductivity according to its saturation level (dry or saturated), using values from the literature tabulated in Table ST4 in Supplementary Appendix A4. This was modeled using the Fourier law, assuming a subsurface with two main layers: an upper 2-km-thick layer of highly fractured megaregolith with moderate density (1700 kg/m3; Parro et al., 2017) and a bottom 7-km-thick denser layer (2850 kg/m3; Parro et al., 2017) of fractured basalt ice-saturated upper crust. A thin, shallow upper layer of less than 10 m consisting of high porosity and low thermal conductivity substrate was not included in the calculations because its small thickness makes it irrelevant to the temperature results.
Temperature profiles stop at a depth of 9 km where seismic data from InSight suggest a structural discontinuity that could reflect the presence of an unfractured (zero-porosity) flood basalt (Kilburn et al., 2022; Xu et al., 2023). Figure 5 shows the three subsurface temperature profiles estimated based on the water/ice boundary being at z = 150, 1000, and 2000 m in depth. In the first case, T = 0°C is reached at a depth of 8.8 km, while in the second and third scenarios, ice melts at a 6.8 and 4.3 km depth, respectively. The melting depths would decrease to 5.0, 3.2, and 1.8 km, respectively, if water is NaCl-saturated. However, as mentioned before, the low abundance of cement minerals inferred by InSight makes this scenario unlikely or perhaps constrained to small and isolated salty lenses.
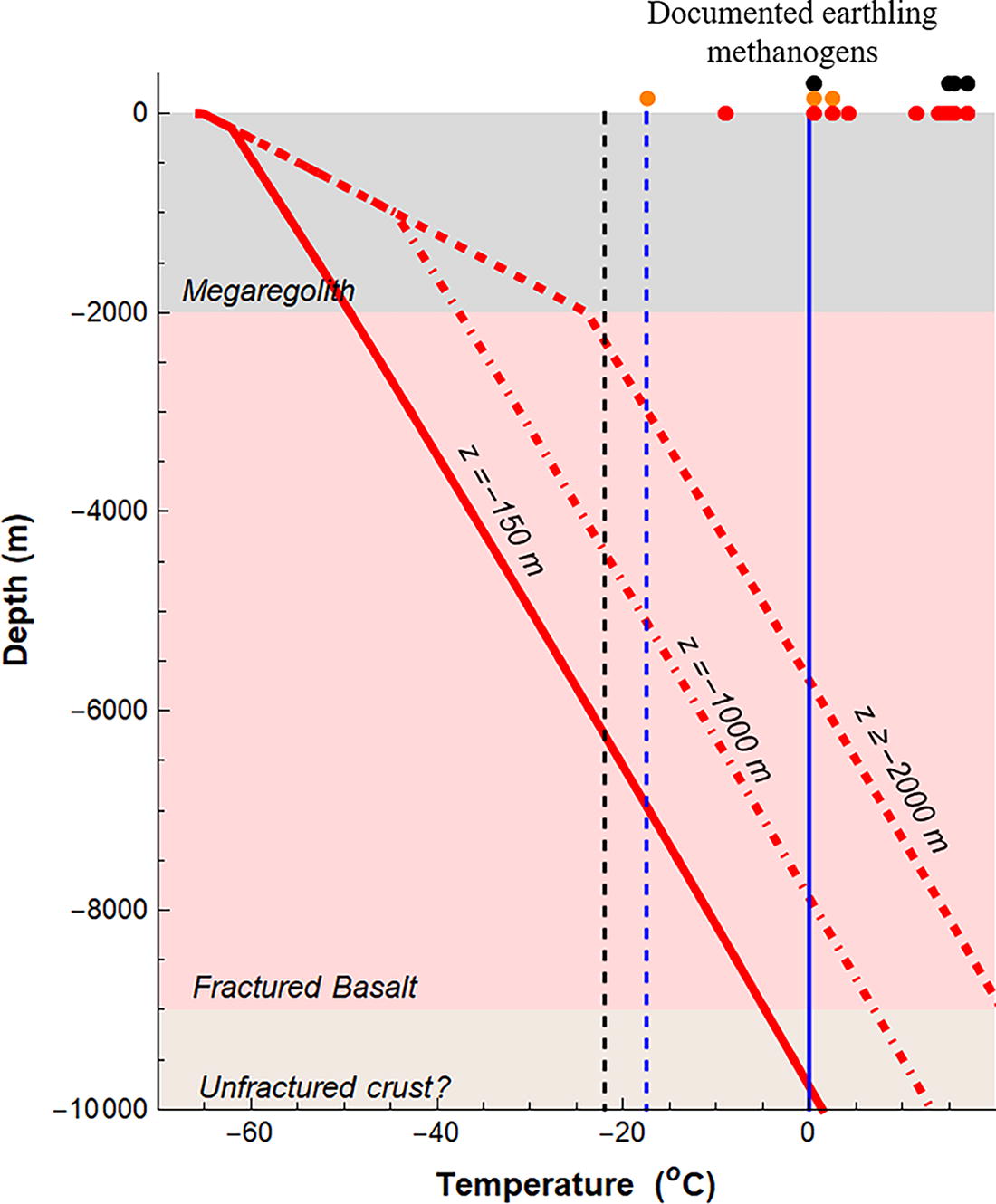
Subsurface temperature profiles (red lines) at 35N, 30W (south of Acidalia Planitia) modeled based on steady-state heat flow using a yearly-average surface temperature of –58°C. The three geotherms correspond to three depths z with no ice above them, reducing thermal conductivity in the upper-most kilometers. Vertical lines show the fusion temperature for freshwater (0°C; blue line), NaCl eutectic temperature (−21°C; dashed black line) and the lowest temperature reportedly tolerated by terrestrial underground methanogens (−17.5°C; Guglielmin et al., 2023; dashed blue line). The top x-axis shows the temperatures at which terrestrial subsurface methanogens are reported in the subsurface, as described in Section 3. The yellow, orange, and red dots are the Methanobacteriaceae, Methanomicrobiaceae, and Methanosarcinaceae families, respectively.
The results provide insights into the following questions raised in Section 1:
(1) Are methanogens significant in DCBF, SGL, and DHAB habitats?
Methanogens are significant in terrestrial subsurface habitats at several study sites. However, they are not ubiquitous and should not be regarded as core members of the terrestrial subsurface ecosystems. Indeed, some DCBF sites that apparently fulfill the conditions for methanogenesis (anoxic fluids, at moderate-high temperature, abundant H2, formate, acetate) do not show unquestionable evidence that they host methanogens.
Thus, the proliferation of methanogens in terrestrial subsurface habitats is a response to intricate interactions with the physical environment and with organisms possessing different metabolisms. H2 is the crucial electron donor for many other anaerobic metabolisms reported in terrestrial subsurface ecosystems. Indeed, under standard conditions, the bioenergetics of hydrogenotrophic sulfate reducers is more advantageous than methanogenesis (Supplementary Appendix A1), and they might outcompete or almost eliminate methanogens (Lin et al., 2006). Mars is a sulfur-rich planet (King and McLennan, 2010), and sulfates have been reported at its surface (McLennan and Grotzinger, 2008; Murchie et al., 2009; Kounaves et al., 2010). Significantly, hydrogen and sulfates can be generated concurrently through the radiolysis of water within rocks containing pyrite minerals (Lefticariu et al., 2010), to turn autotrophic sulfate reduction into a plausible metabolic pathway in the martian subsurface.
(2) Do the environmental conditions of DCBF, SGL, and DHAB shape the significance of subsurface methanogens and their metabolic pathways?
Neither subzero temperatures nor high salinity necessarily poses an insurmountable barrier for terrestrial methanogens (or other microbes). Methylotrophic pathways often dominate in environments characterized by high ionic strength, high sulfate concentrations, low temperatures, and neutral to low pH waters. Conversely, hydrogenotrophs prefer moderate-high temperatures, low ionic strength, low sulfates, and high pH. However, that separation is not absolute. Indeed, strict hydrogentrophs have also been reported at low temperature and high salinity.
The comparison between environments indicates that DCBF sites are more challenging habitats for methanogens than DHAB sites. Two key observations support this conclusion: (1) methanogens are ubiquitous at DHAB but not at DCBF sites and (2) the ionic strength threshold limit for methanogens is around 2 M at DCBF sites, whereas at DHAB sites it reaches approximately 9 M.
(3) Is the almost single-species methanogenic ecosystem reported at Lidy Hot Springs representative for DCBF, SGL, and DHAB systems?
It is well established that the terrestrial methanogens often proliferate in complex syntrophic associations with bacteria and probably with viruses (Evans et al., 2019; Wang et al., 2022). This evidence has even led to the hypothesis that eukaryotes evolved precisely from a syntrophy based on methanogens (López-García and Moreira, 2020). Indeed, methanogens in subsurface environments belong to a taxonomically and metabolically complex community dominated in general by bacteria. Therefore, the monospecific methanogenic microbial community reported at Lidy Hot Springs does not represent a typical microbial community from subsurface terrestrial habitats. Consequently, if Earth-like methanogens were to exist in the martian subsurface, they would likely interact with a range of diverse organisms rather than form single-species ecosystems.
(4) How do the answers to the previous questions impact the viability of terrestrial methanogens in Mars’ subsurface?
If the optimal habitable martian anoxic subsurface is expected to be a cold (subzero) hypersaline water body, attention should preferentially be focused on more versatile methanogens such as the Methanosarcinaceae-like and the Methanomicrobiaceae-like families rather than the Methanobacteriaceae reported in hot waters such as Lidy Hot Springs. The Methanosarcinaceae-like organisms are candidates because they are able to feed on methyl-compounds, H2/CO2, and acetate and tolerate high sulfate concentrations, which might represent an advantage on a sulfur-rich planet such as Mars (King and McLennan, 2010). Nonetheless, both Methanosarcinaceae and Methanomicrobiaceae are likely able to proliferate under these extreme conditions by establishing syntropic associations with fermenters or acetate oxidizers (Pan et al., 2021); thus, they should not establish “one-species ecosystems.” Our literature survey revealed that the availability of H2 and small organics (such as formate and acetate) does not guarantee the proliferation of methanogens at DCBF sites. This situation might be extrapolated to Mars’ subsurface if sites of active production of H2, formate, and acetate exist. Moreover, the apparent reduction of the salinity maxima threshold reported at DCBF sites when compared with DHAB sites might have severe implications for the habitability of putative deep martian rock-fractured fluids. At DCBF sites, other constraints besides salinity might limit methanogens, for example, competition with other metabolisms for limiting resources (i.e., H2), an excess of strong oxidants produced by water radiolysis, or physical factors such as porosity (Tanikawa et al., 2018). Importantly, the in situ salinity tolerance threshold is based on information from NaCl- and Ca/MgCl2-rich brines. More “hostile” (in a biological context) and oxidative salts, such as perchlorates, have been identified on Mars (Clark and Kounaves, 2016). On Earth, natural perchlorate brines are limited to a few hyperarid habitats such as the Atacama Desert (Ericksen, 1981), and knowledge regarding the tolerance of terrestrial biota (including methanogens) to these conditions is largely derived from experimental bioassays. These experiments indicate that it is unlikely that terrestrial methanogens proliferate in perchlorate solutions with an ionic strength greater than 0.03 M (Shcherbakova et al., 2015; Serrano et al., 2019). Therefore, the habitability of a martian subsurface brine in fractured rocks would be drastically reduced if perchlorates accumulated in the martian subsurface.
(5) What environments in Mars’ subsurface could support Earth-like methanogens? What methanogenic pathways might prevail?
By combining the available criteria and data on water availability and the abundance and spatial heterogeneity of HPE, we were able to identify martian regions that might host deep water and a relatively high concentration of radiogenic elements. Our most robust target area is the southern Acidalia Planitia at midlatitude (as low as 35°N).
The target site overlaps with a large area with a high density of cones and domes (Mills et al., 2024) that have been interpreted as mud volcano-like structures (Farrand et al., 2005; Brož et al., 2023). If this interpretation is correct, it supports the presence of fluid (water-saturated mud and gases) upwelling in a not-yet identified period during the Amazonian (Brož et al., 2023) and provides clues about the martian subsurface hydrosphere (Allen et al., 2009) (Fig. 4b). In addition, the target region almost overlaps with an area with highest density of recurring slope lineae (RSL) formations in the entire northern lowlands (Stillman et al., 2016) (Fig. 4b). Although a consensus about mechanisms giving rise to RSL is lacking (McEwen et al., 2021), one of the hypotheses supports the presence of water/brine seepage (Rummel et al., 2014). Finally, relatively close to the target region is the McLaughlin Crater, which has exposed clay and carbonate deposits together with signs of groundwater activity (Michalski et al., 2013) (Fig. 4b). Ultimately, combining all these observations with our analysis led to the conclusion that the subsurface of the southern Acidalia Planitia could hold significant astrobiological potential, thus reinforcing Brož et al.’s (2023) recommendation that this region should be prioritized for future missions.
In the target area, the subsurface temperature profiles have been estimated up to a depth of 9 km. Crucially, according to the InSight mission, salts are apparently scarce in the subsurface at low latitudes. Extrapolating this finding to the Acidalia Planitia implies that the most probable temperature at which porous ice melts would be around 0°C. The depth at which ice melts and methanogens can proliferate depends on the depth of the ice/water datum. A hypothetical subsurface with an ice datum near the surface might be inhabitable because the zero-degree geotherm might be located at approximately 9 km depth. At this depth, the pressure-induced collapse of porosity is a significant possibility. Although this scenario needs further validation, this possibility allows speculation that the lower depth limit of a putative habitable subsurface on Mars might be determined by a physical factor rather than thermal or geochemical factors.
The depth of the zero-degree geotherm can decrease to 4.3 km if the upper 2 km of insulating regolithic layers are dry. If this condition is met, then Earth-like bacteria—Methanosarcinaceae and Methanomicrobiaceae, syntrophic clusters can proliferate at those depths with temperatures in the range 0–10°C. In contrast, more thermophilic methanogenic genera, such as the hydrogenotrophic Methanobacteriaceae, might be constrained preferentially at the highest depths provided that the putative collapse of porosity is not reached.
Conclusions and Perspectives
Two main conclusions can be drawn from the present study as follows:
First, methanogens are significant, but not ubiquitous, in the microbial ecosystems that thrive in the subsurface ecosystems believed to be terrestrial analogs of the martian subsurface. Thus, although methanogens feed on simple molecules and their metabolism can be described by straightforward redox equations—which makes them ideal models for astrobiologists—studies of terrestrial deep temperate–hot crystalline fractures are revealing that these habitats are challenging for methanogens. Consequently, the cold martian subsurface is likely an even harsher habitat for putative Earth-like methanogens.
Beyond the need for water, appropriate environmental conditions, and adequate energetic and carbon resources, the proliferation of methanogens also depends on complex ecological constraints. Indeed, methanogens establish ecological and energetic interactions with organisms with diverse metabolisms; this suggests that the conjecture that they might form mono-specific communities does not fit with what is regularly observed in terrestrial subsurface ecosystems. So, if Earth-like methanogens were to thrive in Mars’ subsurface, they would most likely be members of a complex and diverse ecosystem.
Second, the subsurface of the southern Acidalia Planitia is a putative target region for hosting cold-adapted Methanosarcinaceae-like and Methanomicrobiaceae-like methanogens (if they can associate with bacteria-like organisms). In this region, the radiogenic HPEs are at the highest abundance and subsurface water is likely. This not only brings the 0°C temperature as shallow as 4.3–8.8 km below the surface but also favors water radiolysis, which potentially supplies the energetic resources required for these hypothetical methanogens.
Our knowledge of Mars’ subsurface is advancing thanks to orbiters, landers, and rovers, but critical gaps exist. To make substantial progress in identifying habitable niches in the subsurface of Mars, it will be essential to elucidate the availability of inorganic carbon in the subsurface and to determine more accurately the depth at which water is located and the porosity/fracturing of the regolith, as these factors directly affect the thermal gradients and the efficiency of water radiolysis.
As a result, both our analysis (which builds upon recent advances in understanding Mars’ subsurface) and previous research (which has been more focused on its surface converge) have led us to propose the southern Acidalia Planitia as a promising target area for future missions in the search for extant life in Mars’ subsurface.
Footnotes
Acknowledgments
The authors thank Gareth Morgan and the SWIM team for providing ancillary data for the ice contingency maps, Laura Parro for providing data on Mars-modeled surface heat flows, Mackenzie Mill for providing location of pitted cones, Martin Schimmel for the enlightening conversations about the InSight mission, and Alfonso Mota for comments on a previous version of this article. The authors thank S.L. Cady and the anonymous referee for comments and suggestions that strongly helped to improve their initial submission.
Authors’ Contributions
A.B. conceived the study, collected the information from the literature, and wrote the original draft followed by review and editing. O.F., R.B.-F., and D.G.-C. collected the information from the literature, wrote and supervised the article, made figures, and reviewed and edited the final version.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by MCIN/AEI/10.13039/501100011033 (PID2021-123735OB-C22) and by “ERFD A way of making Europe.” A.B. is a member of the SGR976. O.F. acknowledges financial support from the Departament de Recerca i Universitats of Generalitat de Catalunya through grant 2021SGR00679. O.F. was (partially) supported by the Spanish MICIN/AEI/10.13039/501100011033 and by “ERDF A way of making Europe” by the “European Union” through grant PID2021-125627OB-C31, and through the “Center of Excellence María de Maeztu 2020–2023” award to the ICCUB (CEX2019-000918-M). D.G.-C. was funded via the projects AGAUR 2021 SGR00410 and PID2022-139943NB-I00.
Supplementary Material
Supplementary Appendix A1
Supplementary Appendix A2
Supplementary Appendix A3
Supplementary Appendix A4
Supplementary Appendix A5
Supplementary Appendix A6
Supplementary Appendix A7
Supplementary Data S1
Supplementary Data S2
Associate Editor: Jack Mustard
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.