
Abstract
Laminae are millimeter-scale features in rocks created by physiochemical processes that can be influenced by the presence and activities of communities of organisms that occur as biofilms and microbial mats. The structure and composition of laminae reflect the processes involved in their formation and can be preserved in the rock record over geologic time; however, diagenetic and metamorphic alteration can lead to the loss of primary information and confusion over the interpretation of their origins. As potential records of ancient life, laminae can preserve evidence of microbial activity over billions of years of Earth’s history. On planetary bodies such as Mars, laminae in sedimentary rocks are common and represent significant features of interest that can record habitable conditions (e.g., the presence of liquid water) at the time of their formation. Here we review the significance of laminae as targets for astrobiological exploration. We discuss common mechanisms by which laminae form in natural environments on Earth, present arguments and evidence for laminae as potential biosignatures, and describe how such information is presented in the NASA Life Detection Knowledge Base.
Introduction
Laminae produced from biofilm growth in and on sediments constitute a prime morphological feature of interest in the search for fossilized evidence of microorganisms in the earliest rock record on Earth and potentially other planetary bodies. Laminae can form through a variety of nonbiological depositional processes that involve detrital sediment accumulation and chemical precipitation; their formation, however, can be influenced by the presence, behavior, and metabolic activities of organisms, especially when those communities live at sediment surfaces in biofilms (even those only a few cell layers thick) and in thicker (millimeter to centimeter) trophically segregated microbial mats.
Laminae are important targets for astrobiology-oriented missions, particularly surface exploration activities on rocky bodies such as Mars, as they are readily identifiable using a wide variety of imaging and analytical instruments. Laminae are distinguishable by marked changes in color, granulometry, and chemical composition compared to their surrounding host deposits. In rocky outcrops, laminae appear in cross-section as thin dark or alternating light and dark layers and exhibit a variety of geometries (discussed in Section 2). Laminae range in size from micrometers (i.e., requiring microscopic observation) to millimeters (i.e., visible to the naked eye). They can occur as individual layers or be superimposed upon antecedent laminae to form thicker stacked units called stromatolites (discussed in Section 2.2.5). In this article, we discuss the common processes involved in the formation of abiotic and biotically influenced laminae, their significance as astrobiological targets, and how they are described as features of interest in NASA’s Life Detection Knowledge Base (LDKB), with emphasis on Precambrian examples documented in field and laboratory studies.
The LDKB, an online community tool, presents collated information relevant for understanding potential biosignatures that may be detected beyond Earth (Hoehler et al., 2025; Pohorille et al., 2025). A variety of potential biosignatures in the LDKB are organized into broad categories of “Chemistry,” “Structure,” and “Activity” (Davila et al., 2025; Shkolyar et al., 2025, Perl et al., 2025; Des Marais et al., 2025). Morphological features of interest, such as laminae, are grouped together in the Structure category of the LDKB. Arguments and evidence regarding the biogenicity of structural features are organized by observable characteristics, namely “size,” “spatial distribution,” “shape/form,” “orientation,” and “texture/fabric” (Shkolyar et al., 2025). We focus here on the description and interpretation of these characteristics with respect to laminae, which represent a key indicator of microbial life in the sedimentary rock record on Earth (e.g., Walter et al., 1980; Buick, 1992; Walsh, 1992; Riding, 2000; Noffke, 2000, 2010, 2021; Noffke et al., 2001; Cady et al., 2003; Westall et al., 2001, 2006, 2015a, 2015b; Awramik and Grey, 2005; Oehler and Cady, 2014; Homann et al., 2015, 2016, 2018; Trower and Lowe, 2016; Hickman-Lewis et al., 2016, 2018, 2023b; Homann, 2019; Bosak et al., 2021; Noffke et al., 2024). In addition to the structural characteristics discussed here, biogenic laminae may also retain other key signatures of life that include carbonaceous remnants of the original microbial community (e.g., biomolecules and their degraded residues), characteristic stable isotope fractionation patterns, and trace element concentrations consistent with biological activity, among others. As discussed in Section 2.2.5, multiple lines of converging evidence are necessary to discern biotic from abiotic laminated structures resembling microbial remains; examples of such evidence can be found in the “Chemistry” and “Activity” categories in the LDKB (e.g., Des Marais et al., 2025).
Laminae and Their Characteristics
In the field of sedimentology, the thickness of an individual lamina is regarded as less than 1 cm when observed in vertical cross-section, whereas thicker layers are considered “beds” (e.g., McKee and Weir, 1953; Tucker, 2009; Collinson and Mountney, 2019). Consistent with this, we define primary sedimentary laminae as micrometer-to-millimeter-scale layers in rock that consist of detrital and/or chemically precipitated material deposited commonly, though not exclusively, in an aquatic setting. In depositional environments, laminae can form as a result of changes in fluid energy and/or geochemistry (e.g., temperature, pH, Eh, ion saturation, and density), variations in sediment supply reflecting environmental shifts, or interruptions at the sediment–water interface during mineral accretion. These mechanisms can be driven by abiotic processes, biological activity, or a combination thereof. Evidence of such processes can be preserved in lithified laminated sedimentary rocks for geological timescales and thus serve as attractive targets for understanding the paleoenvironmental conditions and habitability of the original depositional environment. Although not intended to be an exhaustive list, some of the major environments and processes responsible for the formation of common lamination types, including those formed by the influence of biological activity, are described below.
Occurrences and composition
Laminae form in nearly every sedimentary environment and exhibit a wide range of structures and compositions from the macro- to microspatial scales. On Earth, laminae most commonly form in subaqueous environments at the sediment–water interface, such as shallow- and deep-water lacustrine, riverine, and marine settings. They can also form in environments where chemical precipitates accumulate around hot and cold springs, desert playas, and cave walls (e.g., Spear et al., 2007; Handley et al., 2008; Gonzalez-Pimentel et al., 2018; Lingappa et al., 2021). Laminae also form in the subsurface, for example, among caliche deposits in arid environments (e.g., Reeves, 1970 ), within the void space of vesicular igneous rocks, and among fracture openings where fluids can infiltrate all types of hard rock over geological time (e.g., Cavalazzi et al., 2007, 2021; Homann et al., 2016; Ivarsson et al., 2020; Neubeck et al., 2021; Lima-Zaloumis et al., 2022). Laminae can also form in subaerial environments (i.e., at the air–sediment interface) where wind dominates, such as aeolian ergs (e.g., Collinson and Mountney, 2019, p. 126).
Laminae formed in association with microbial activity (discussed in Section 2.2.4) are common features in Precambrian sedimentary rocks (e.g., Grotzinger and Knoll, 1999; Riding, 2000; Bosak et al., 2013; Grey and Awramik, 2020). Such structures can be found today forming in a variety of surface and subsurface environments (e.g., Hoffman, 1976; Reid et al., 2000; Heim et al., 2017; Vahrenkamp et al., 2024). Thus, laminae as structural features provide a long-term record of both direct and indirect aspects of biological processes that have been occurring in situ since the early appearance of life on Earth. We hypothesize that laminated structures preserved within sedimentary rocks on other planetary bodies where microbial life has emerged could contain similar paleobiological information.
The environments where laminae form are broadly categorized below into detrital and chemical sedimentary systems, each of which is dominated by specific depositional processes that can result in characteristic textures that manifest in lithified rocks. It is important to note that physical and chemical sedimentary processes are not mutually exclusive, as both detrital accumulation and chemical precipitation often occur simultaneously.
Detrital laminae
Detrital laminae commonly develop when clastic particles accumulate atop the bottom sediments of a water body (e.g., rivers, lakes, and marine environments, including deltas, lagoons, and estuaries). Detrital components can range from mud-sized (<4 µm) to silt-sized (<62.5 to >4 µm) to sand-sized (<2 mm to >62.5 µm) particles, or a combination thereof. These particles may be composed of individual minerals, rock fragments, biologically derived organic material, and, in Phanerozoic beds, shelly material. The composition and grain size of depositing clastic materials can change gradually over time due to shifts in sediment sources and supply (e.g., through weathering and erosion) or changes in water energy and depth, or they may oscillate regularly due to seasonal processes such as glacial meltwater influx that results in the deposition of laminated couplets known as varves (e.g., Anderson and Dean, 1988). Sediment characteristics may also change suddenly via bioturbation or during catastrophic events such as storms and earthquakes (e.g., Kreisa, 1981; Sabatier et al., 2022). In any given depositional environment, the final grain sizes and shapes of detrital sediments are influenced by the weathering of continental source rocks, transport of particles by flowing fluid (wind, water, or ice), and subsequent reworking and abrasion during bed load transport. In combination, these processes contribute to variations in detrital sediment that eventually manifest as distinctive laminated structures visible in lithified sedimentary rock.
Laminated chemical sediments
Laminated chemical sediments develop within ion-supersaturated aqueous environments such as evaporative subaerial settings and, in combination with cooling, in hot spring settings via the precipitation of amorphous mineraloids and microcrystalline minerals from aqueous solutions. Environments where chemical sediments commonly form include shallow marine environments, hot and cold springs, lakes and sabkhas, and subsurface environments that include fractures and vesicles. Chemical precipitates commonly found in such settings include opaline silica, carbonates (e.g., calcite, dolomite, aragonite, and magnesite), anhydrous and hydrous sulfates (e.g., gypsum, barite, and mirabilite), chlorides (e.g., halite and sylvite), phosphates (e.g., apatite), and iron oxides (e.g., ferrihydrite and hematite). In chemical sediment precipitating environments, the size of crystals (and hence the resulting thickness and texture of laminae composed of such crystals) is influenced by several factors that include the rate of nucleation and the duration of time that fluid supersaturation is maintained within the local environment; crystalline precipitates can continue to increase in size, depending on the duration of undisturbed growth during relatively quiescent supersaturated conditions (García-Ruiz and Otálora, 2015). In aqueous environments, chemical precipitates may form at or beneath the sediment–water interface, or they may precipitate within the water column or at the air–water interface and settle onto the benthic floor. Such processes lead to distinctive textures that can be recognized as primary forms (e.g., Warren, 2006, p. 22). Laminae may also form by secondary diagenetic processes, as is the case with concretions, geodes, and botryoids (see Section 2.2.3), where chemically oscillating reactions result in the precipitation of fine-scale laminar patterns that consist of quartz, apatite, and/or carbonate (Papineau et al., 2021). Due to their relatively higher solubilities as compared to resistant siliciclastic material, chemical sediments can be susceptible to diagenetic effects, including mineral dissolution, replacement by more stable mineral forms, and crystal coarsening that results in the destruction/homogenization of primary laminae (e.g., Warren, 2006, p. 38; Flügel and Munnecke, 2010, p. 237; Lima-Zaloumis et al., 2024).
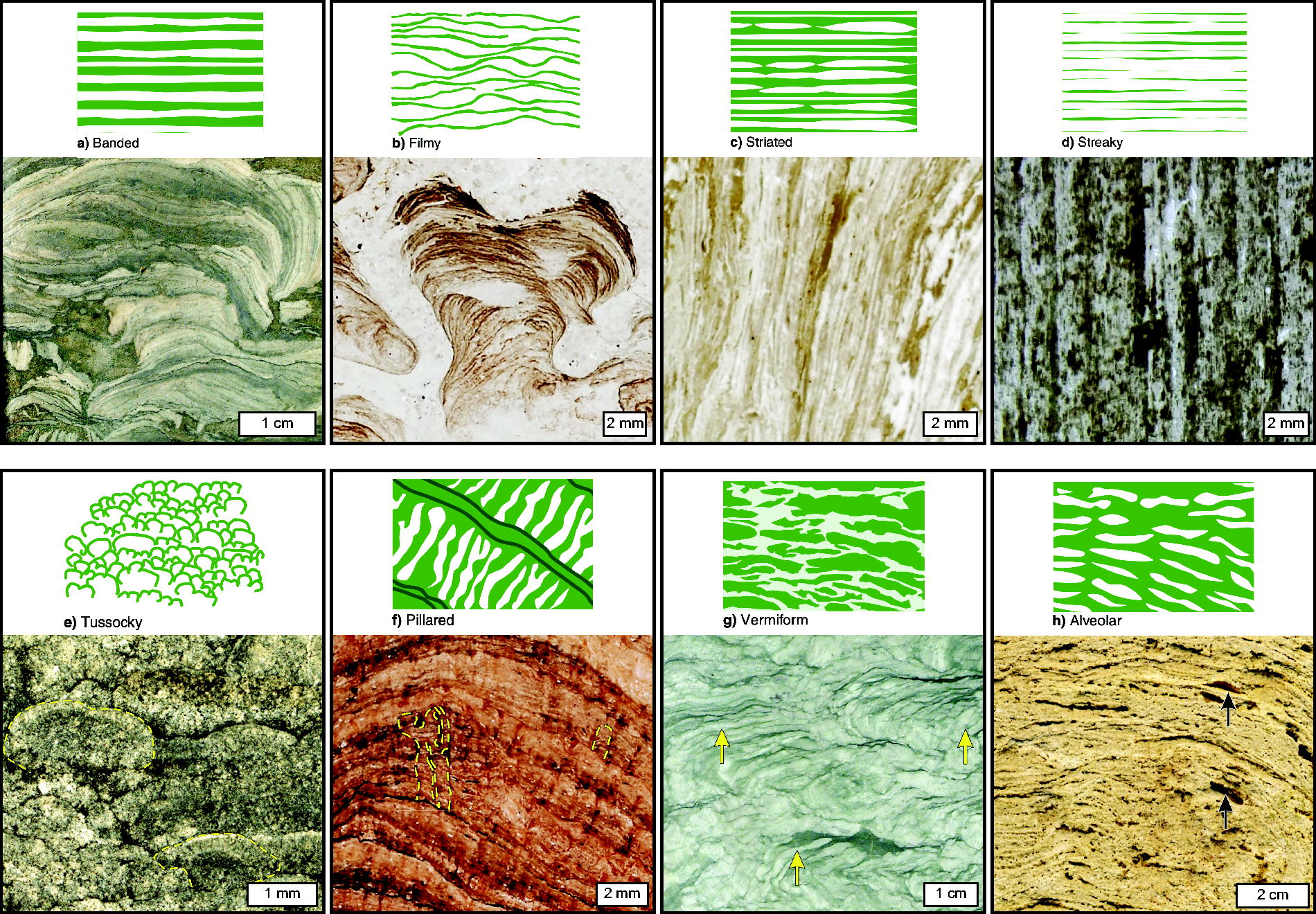
Common lamination geometries
In the rock record, laminae exhibit a variety of shapes and orientations that reflect their formation origins and postdepositional history. Below, we categorize common lamination geometries observed in sedimentary rocks (summarized in Table 1), each of which can reflect abiotic and/or potentially biological growth mechanisms. Key structural characteristics, such as orientation, thickness, and lateral extent, can serve as first-order indicators of potential biosignatures and are useful for evaluating environments during field investigations, whether on Earth or in the context of human or robotic planetary exploration. While any lamination geometry may incidentally preserve biosignatures (e.g., organics deposited in place from the surrounding environment), irregular wavy or crinkly geometries are more commonly associated with the potential presence of fossilized microbial remains (discussed in Section 2.2.4). However, several abiotic processes are also capable of producing similar irregular geometries (discussed in Section 2.2.5).
Summary of Lamination Types Reviewed in This Manuscript
Summary of Lamination Types Reviewed in This Manuscript
Parallel horizontal laminae that display a flat, smooth morphology are commonly formed in aqueous environments by the settling of fine-grained sand, silt, and mud through the water column, often from slow-moving sediment clouds above the depositional surface, or by low-density turbidity currents (Tucker, 2009, p. 25). They can also form under upper flow regime conditions, where high-velocity currents generate sufficient energy to plane off or erase preexisting bedforms such as ripples and dunes (Collinson and Mountney, 2019, p. 118). In chemically precipitating environments, flat laminae can form by evaporation and ion supersaturation of fluids; seasonal precipitation and settling of particles through the water column, as well as precipitation on the benthic floor, lead to the development of parallel, laterally continuous laminae that can be traceable for tens to hundreds of kilometers (Fig. 1A; Anderson et al., 1972; Warren, 2006, p. 23).

Examples of lamination morphologies formed principally by abiotic processes.
Although flat laminae can contain biological constituents, such as organics transported from a surrounding basin or formed in place (e.g., by nonlithifying biofilm growth or settling of phytoplankton through the overlying water column), their flat, parallel geometry is not strongly indicative of past biological activity. It should be noted, however, that some flat laminated structures have been interpreted to form in the presence of or directly influenced by microorganisms, for example, in the case of “stratiform” stromatolites (Walter, 1976), “biolaminites” (Gerdes and Krumbein, 1987), and “cryptalgal laminites” (Monty, 1976); in such cases, lamina alternate between light-colored mineral precipitates and dark carbonaceous layers. Additionally, some postdepositional modifications of flat laminae may more clearly constitute potential structural biosignatures, such as disruptions caused by grazing or burrowing Metazoa (e.g., Taylor et al., 2003). Similar activities by prokaryotes have been inferred at the microscale among glass and carbonate phases (e.g., Garcia-Pichel, 2006; Ivarsson and Holm, 2008; Fisk et al., 2019).
These structures commonly form in aqueous environments characterized by unidirectional or bidirectional fluid flow, such as wave- or current-dominated settings, and can develop in subaerial environments where wind is the primary transport mechanism. As observed in cross-section, these lamination geometries typically occur as layers that dip at one or more angles, often forming distinctive crest and trough patterns that may be sinuous, linguoid, or asymmetrical, among other forms (e.g., Fig. 1B; McKee and Weir, 1953; Rubin and Carter, 1987; Tucker, 2009, p. 27). The migration of ripple marks over time results in the deposition of cross-laminated sequences, which in vertical cross-section reflect two dimensions of the original three-dimensional structure of ripples or larger-scale dunes. Paleoenvironmental information such as paleocurrent direction and strength may be inferred by analyzing cross-bedded laminae and their orientations in space. The presence of ripples and cross laminae may indicate the past presence of water based on the ripple index (a calculation of wavelength to wave height; Tanner, 1967); therefore, such structures serve as important astrobiological targets to understand an environment’s past habitability potential.
In environments dominated by ripple and dune bedforms, biofilm-forming microorganisms can template onto existing sediment relief (a process termed “imprinting”; Noffke et al., 2001). Continued biofilm growth may act to stabilize ripples and enhance their preservation during later reworking (Noffke et al., 1996; Noffke, 1998). However, while cross-laminated sediments may incidentally be associated with biosignatures, such as burrows and other ichnofossils, body fossils, and extracellular polysaccharides (EPS) secreted by biofilm communities (e.g., Noffke et al., 2006, 2008, 2013), their geometry alone does not indicate biological activity.
Concentric and botryoidal lamination
Concentric laminae can form as cements around moving grains in shallow wave-dominated environments. Concentrically laminated spherical to subspherical particles smaller than 2 mm are termed ooids, while larger grains termed pisoids can take on a variety of shapes (Peryt, 1983; Tucker and Wright, 2009, p. 2). Biofilms can develop on the surfaces of moving grains and influence the precipitation of irregular laminae (see further discussion in Section 2.2.4), which manifest as couplets of dark organic-rich laminae alternating with lighter-colored organic-poor laminae (Flügel and Munnecke, 2010, p. 124). Such structures are termed oncoids (originally described by Heim, 1916), where lamination thickness is dependent upon the activity of cyanobacteria and changes in fluid geochemistry over time (e.g., seasonal conditions affecting carbonate saturation).
Concentric laminae may also form within sedimentary concretions, botryoids, geodes, granules, and rosettes (Fig. 1C, D). Generally, spheroidal concretions precipitate via diffusive transport of reactants, while elongate concretionary forms precipitate via advective flow of reactants. Concretions precipitate via two endmember growth mechanisms: radial and pervasive. Radial growth occurs when cements preferentially precipitate in the subsurface around a nucleus and the concretion grows radially outward to form concentric laminae (e.g., Mozley, 1996; Raiswell and Fisher, 2000; Potter et al., 2011); pervasive growth does not produce laminae. Concretionary and botryoidal laminated structures can be associated with organics, which may be derived from biological or abiotic sources, and they can be preserved over billion-year timescales (Sugitani et al., 2007, 2013; Brasier et al., 2011; Wacey et al., 2012; Papineau et al., 2021; Papineau, 2024), although some ancient examples may have been subject to younger postdepositional oil migration (Coutant et al., 2022).
Diagenesis can also produce Liesegang bands where reactants diffuse toward each other in sediments, which creates laminae with spacing controlled by the rate of diffusion (Liesegang, 1896; Ostwald, 1897). These forms can be identified by their crosscutting stratification and often concentric geometries. They can occur as poorly cemented laminae, created by a layered coloration pattern that typically involves iron or manganese oxides, or they can be so well cemented that they weather differentially and produce positive topographic features in weathered rocks (e.g., Chan et al., 2000).
Irregular laminae and the role of biology
Unlike abiotic processes that produce relatively uniform deposits of flat-lying, isopachous (even-thickness), cross-layered, or regularly oscillating layers of sediment, micrometer-to-centimeter-scale benthic biofilm and mat communities can directly produce or influence the formation of irregularly shaped laminae at comparable scales (Figs. 2 and 3). It is this irregularity that suggests biogenicity. Microbially associated laminae are commonly described as wavy, which Walter (1972) defined as having flexures of wavelength >2 mm, and wrinkled, defined as having flexures of wavelength ≤2 mm (Grey and Awramik, 2020, p. 129). Other descriptions of microbially associated laminae in the literature include terms such as undulating, nonisopachous (i.e., uneven thickness), crenulated, wispy, and crinkly, among others. Irregular biogenic laminae in sediments may exhibit discontinuities in lateral extent, bifurcation, anastomoses, and tapering along ends. Additionally, they can be associated with structural elements such as tufts (Fig. 3), horizontally elongated pores (i.e., “fenestrae”), trapped siliciclastic grains, and encrusted/permineralized microfossil structures (e.g., Flügel and Munnecke, 2010; Noffke et al., 2013; Homann et al., 2015; Hickman-Lewis et al., 2018; Hickman-Lewis and Westall, 2021).

Laminations interpreted as microbial mat remains.

Laminations exhibiting tufted morphology consistent with microbial mat remains.
The irregular distribution and morphology of microbially associated laminae reflect nonuniform colonization and growth characteristics of biofilms, as well as their responses to a dynamic environment with changing fluid energy regimes, sediment burial rates, and nutrient availability, among other factors. In natural environments inhabited by microorganisms, chemical and radiative energy availability can vary over micrometer spatial scales and time owing to dynamic environmental processes (e.g., Jørgensen and Des Marais, 1990; Visscher and Stolz, 2005; Lichtenberg et al., 2020). For example, zones inhabited by photosynthetic communities at the sediment–water interface may become subject to sediment accumulation and burial, as is the case in most active shallow marine, lacustrine, and hot spring environments on Earth; as zones of energy availability migrate in space (e.g., upward as deposition proceeds), biofilm communities can react via active motility or passive dispersion, preferentially colonizing zones with greater access to nutrients and energy supplies necessary for cellular upkeep and reproduction (e.g., Sauer et al., 2022). Biofilm colonization also depends upon surface adhesion and roughness characteristics, favorable fluid flow velocities, and temperature (Donlan, 2002). Biofilm thicknesses can vary depending on several factors that include turnover rates (i.e., the balance between autotrophic growth and heterotrophic decomposition; Reid et al., 2024), community composition (e.g., Murga et al., 1995), and the hydrodynamic environment (Vieira et al., 1993; Liu and Tay, 2002). Biofilms can mature into centimeter-thick layered microbial mats with complex, stratified community structure (e.g., Krumbein et al., 1977; Des Marais, 1995; Visscher and Stolz, 2005), although thicker mats with high biomass production may hinder their lithification (and thus preservation in the rock record) by limiting ion diffusion (discussed further in Section 2.2.6 and Reid et al., 2024). Living microbial mats can also exhibit structural complexity, for example, via the formation of trapped gas bubbles, tufts, pinnacles, and reticulates (Walter et al., 1976; Gerdes, 2007; Bosak et al., 2009; Shepard and Sumner, 2010). When observed in a vertical cross-section, such structural complexity lends to the appearance of irregular lateral mat geometries. In addition to variations in biofilm distribution and structure, microbial interactions with (and influence on) chemical and clastic sedimentation also affect the texture of lithified, microbially influenced laminae, as discussed below.
Biofilm communities are commonly composed of a consortium of species that utilize a variety of autotrophic and heterotrophic metabolisms; the net effect of metabolic activity can result in shifting the pH of the surrounding microenvironment such that mineral supersaturation and precipitation is favored (referred to as the “alkalinity engine” in carbonate precipitating systems; Dupraz et al., 2009; Gallagher et al., 2012). Biofilms also contribute to sedimentation by reducing energy barriers to nucleation; thus, they serve as favorable sites for mineral precipitation (Braissant et al., 2007; Dupraz et al., 2009; Gallagher et al., 2010) even in cases when the ambient aqueous medium is undersaturated with respect to the precipitating phase (Moore et al., 2020). Cell lysis and heterotrophic EPS degradation can also release cations that contribute to mineral saturation and precipitation (Dupraz et al., 2009). In addition to extracellular mineral precipitation, some organisms precipitate minerals intracellularly for a variety of purposes that include use as navigational aids within Earth’s magnetic field (e.g., Bazylinski et al., 2007) and as possible inorganic carbon stores (e.g., Benzerara et al., 2014; Cam et al., 2018). Sulfur-metabolizing microbial consortia can form pyrite deposits with specific morphologies (e.g., amorphous nanopyrites and framboids) that can serve as evidence for biogenicity (Marin‐Carbonne et al., 2018, 2020; Perl et al., 2025). These and other biominerals can become preserved and contribute to the overall lithic texture and/or elemental component of biologically associated laminae (e.g., Chang et al., 1989; Perl et al., 2025).
Biofilm influence on clastic sedimentation and the formation of MISS
Microbial activities can influence clastic sedimentation and the formation of characteristic structural features recognizable in the rock record (Noffke et al., 2001). Through the production of sticky EPS, biofilms can induce sediment baffling and trapping (enhancing the accumulation of sediment), biostabilization (protecting sediments from physical erosion), and sediment binding via cellular and/or extracellular incorporation of sediment into biofilm fabrics (e.g., Reid et al., 2000; Gerdes et al., 2000; Noffke et al., 2001, 2024; Frantz et al., 2015; Noffke, 2021). These interactions can be recorded in rocks and serve as indicators of biogenicity even when direct evidence of microorganisms (e.g., cellular microfossils and/or organic carbonaceous remains) is absent. For example, detrital grains adhered to laminae at steep interfaces beyond the angle of repose (i.e., the maximum angle at which particles will remain in place without sliding) can indicate evidence of trapping and binding onto “sticky” organic mat surfaces that once covered high-angle surfaces (e.g., Mitchell, 1985; Schieber, 1998; Hofmann et al., 1999; Riding, 2000; Kenter et al., 2005; Allwood et al., 2006, 2009; Westall et al., 2006; Murphy and Sumner, 2008; Le Heron et al., 2013; Homann et al., 2015; Tice et al., 2017; Wilmeth et al., 2019). Such interpretations are supported by laboratory experiments that have investigated steep-angle grain trapping among living mat communities (Frantz et al., 2015).
Evidence of sediment biostabilization can also be recorded in sedimentary rocks (Noffke et al., 1997, 2024); eroded fragments of biofilms in the form of mat chips and roll-up structures indicate initial plasticity and cohesiveness of an organic-rich layer and a strong enough water current able to transport bioclasts (Fig. 4; Simonson and Carney, 1999; Noffke et al., 2001; Tice and Lowe, 2004; Tice and Lowe, 2006; McLoughlin et al., 2023; Howard et al., 2024). In the rock record, these features are recognized as microbially induced sedimentary structures (MISS; Noffke et al., 1996). In addition to mat chips and roll-ups, MISS can exhibit a variety of morphologies and sizes that include multidirectional ripple marks, polygonal oscillation cracks, erosional remnants and pockets, and wrinkle structures (Noffke, 2021; Noffke et al., 2024). Such structures are recognized in sediments from the earliest Archean to modern settings, with specific examples from the ancient rock record described further in Section 3.1.
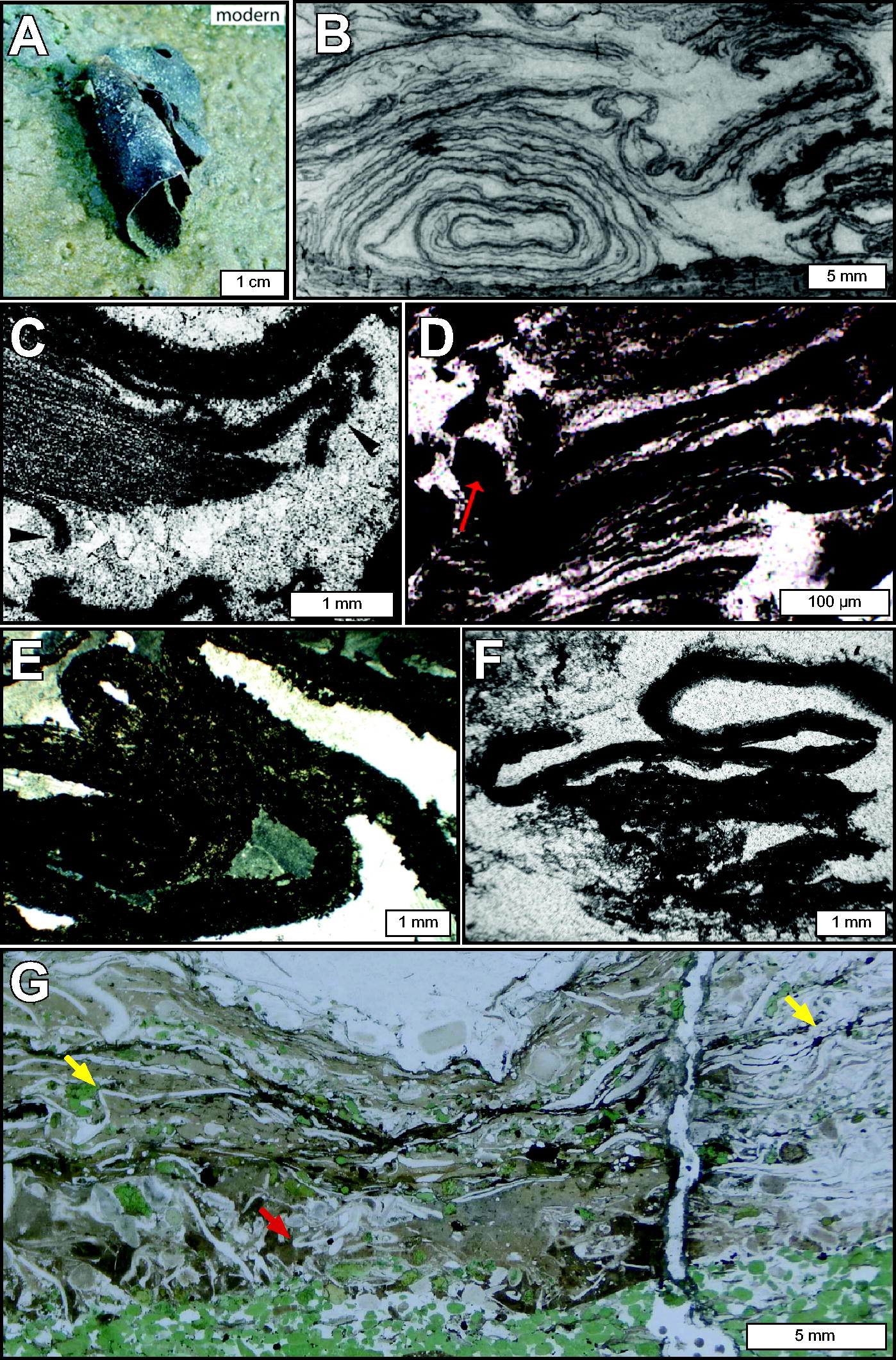
Lamination morphologies indicative of microbial mat cohesion.
2.2.4.3. Laminae as defining features of stromatolites
Biologically influenced laminae can accumulate into organized stacks that form distinctive emergent structures known as stromatolites. Grey and Awramik (2020) define stromatolites as “a laminated organosedimentary structure produced by precipitation or by sediment trapping and binding as a result of the growth, behaviour, and metabolic activity of microorganisms, principally cyanobacteria.” Internal laminae are the defining feature of stromatolites, where the word “stromatolite” [Stromatolith] was derived originally from the laminated appearance of microbial bioherms (Kalkowski, 1908). Stromatolites and MISS constitute laminated categories of “microbialites,” defined by Burne and Moore (1987) as “organosedimentary deposits formed from interaction between benthic microbial communities (BMCs) and detrital or chemical sediments.” In the rock record, stromatolites exhibit a wide range of macroscale morphologies (Walter, 1976). In planar (top-down) view, individual stromatolite bioherms (or “heads”) may appear circular, ovate, lobate (e.g., Fig. 5A), or linear, among other forms (Grey and Awramik, 2020, p. 55). In the cross-sectional view, stromatolite profiles can take a variety of shapes that include domes that develop over time to form columns and conical and branching structures (Fig. 5; Grey and Awramik, 2020, p. 67). Internally, stromatolites exhibit a variety of lamination architectures (Fig. 6) that Grey and Awramik (2020) describe as “products of specific combinations of microorganisms, sediment, cement and any other components, formed within a limited timeframe” and “includes the manner in which successive combinations are related to each other.”

Morphological varieties of stromatolites in plan view and vertical cross-section.
Stromatolite lamination architectural types and associated examples, reproduced from Grey, K. and Awramik, S.M. 2020 under Creative Commons Attribution 4.0 License.
As with the development of individual lithified lamina by microbial mats and biofilms, the same microbial interactions influence the formation of stromatolites, which include trapping and binding of detrital sediment by sticky EPS, metabolic processes that contribute to mineral saturation and precipitation, and organic-mineral templating. Informed by decades of study in active microbialite-forming systems, Reid et al. (2024) described stromatolite formation as a “microbial balancing act” that involves the production of biomass, mineral grain agglutination, and the precipitation/dissolution of chemical sediments. The key to the formation of layering in biosedimentary structures is the ability of a biofilm or microbial mat to eventually become lithified, generating a stable substratum upon which subsequent biofilm or mat communities colonize and grow. Mat lithification depends upon the ratio of mineral precipitation and accumulation to biomass production, processes that are influenced by extrinsic (e.g., environmental pH, temperature, hydrodynamics, and cation concentrations) and intrinsic factors (e.g., metabolic activity, abundances of microbial cells, and extracellular organic matter).
Changes in both extrinsic and intrinsic processes that affect the balance of biomass growth and mineral precipitation/accumulation influence the lithification, upward accretion, and ultimate preservation as microbialite structures in the rock record (Reid et al., 2003, 2024; Dupraz et al., 2006). Periodic or episodic changes in this balance, such as those driven by environmental changes or interruptions in sediment supply, can produce lamination in microbialites. In other words, macro- and microscale microbialite morphology reflects not only the microbial responses and influences on chemical and clastic sedimentation, but it is also strongly influenced by environmental factors such as aqueous fluid energy and shoreline physiography (Suosaari et al., 2019), ion availability and diffusion (Petroff et al., 2013), and even large-scale climate patterns (Petryshyn et al., 2012).
As structures preserved in the rock record, microbialites represent the cumulative sedimentary record of biofilm and microbial mat activities over time (Hofmann, 1969; Grey and Awramik, 2020). In stromatolites, each lamina represents a snapshot of growth and development, marking past positions of the microbe–sediment–water interface, that is, the “synoptic profile,” while the height of a stromatolite documents the progress of the stromatolite-building processes (Hofmann, 1969). As described by Hofmann (1969), “The fundamental feature of a stromatolite is its mineralogically or texturally differentiated lamination. The synoptic morphology of the laminae in an assemblage represents the dynamic equilibrium conditions as well as the micro-bathymetries of the biologically, chemically, and mechanically active interfaces in the environment.” This is also true for MISS, though the combination of chemical and mineral precipitation that lead to upward sediment accretion does not play a role in MISS formation (Noffke et al., 2001).
As products of microbial activity, stromatolites and MISS represent some of the oldest evidence of life on Earth; notable examples are found in 3.4–3.5 billion-year-old (Ga) rocks from the Dresser and Strelley Pool formations in the Pilbara, Western Australia (e.g., Buick et al., 1981; Hofmann et al., 1999; Allwood et al., 2006; Noffke et al., 2013, 2024), and among 3.4–3.3 Ga Kromberg and Mendon formations in the Barberton greenstone belt in southern Africa (e.g., de Wit et al., 1982; Byerly et al., 1986). Additional examples of ancient, laminated structures are included in the LDKB and are further discussed in Section 3.
Several abiotic processes can result in the formation of laminae with irregular geometries (e.g., nonisopachous, discontinuous, wavy) that resemble those formed in association with microbial biofilms or mats. Stylolites, for example, are irregular abiotic structures (Fig. 1E) characterized by zig-zagged laminae formed by pressure solution. This process results in the removal of mineral material and the accumulation of an insoluble residue, resulting in morphologies that can be confused with microbial textures. However, such structures may be distinguished by their chemical composition and microscale texture (Noffke et al., 2008), particularly by their highly irregular undulation, which is inconsistent with microbialite morphogenesis (Hickman-Lewis et al., 2023a).
Soft-sediment deformation caused by slumping, slope failure, shear stress, and liquefaction can also form irregular “convolute” laminae and wrinkle structures, which are particularly common among turbidite sequences (Fig. 1F; Mills, 1983; Tinterri et al., 2016; Davies et al., 2016). Additionally, “hummocky” and “swaley” cross laminae form irregular curved geometries that occur in fine-to-medium-grained sandstones in shelf environments. These structures exhibit nonuniform thicknesses with thinning toward convex-up hummocks (topographically high areas) and thickening around swales (low areas of sedimentation) atop complex wave-scoured surfaces (Dott and Bourgeois, 1982; Collinson and Mountney, 2019). Notably, this relationship is inverse to stromatolite lamination architectures, where layer thickness is commonly greatest toward the apex of lamination curvature in the direction of upward growth, representing topographic highs during accretion. Irregular rippled laminae may also be formed by the adhesion of moving sand grains onto wet surfaces in windy environments (forming “adhesion ripples”; Davies et al., 2016 and references therein). Wrinkly laminae have also been posited to form by biotite exfoliation during weathering and interfoliar growth of pyrites in biotite, forming structures that resemble microbial mat remnants (Lepot, 2020, p. 19, and references therein).
Metamorphism can also produce irregular fabrics (e.g., Fig. 1G) that may complicate or obscure the interpretation of laminated structures. In the absence of direct evidence of microorganisms (e.g., carbonaceous microfossils), it can be particularly difficult to resolve the origin of stromatolite-like structures (i.e., “stromatoloids,” Buick et al., 1981; or “dubiostromatolites,” Awramik and Grey, 2005). For example, Nutman et al. (2016) interpreted structures occurring in 3.7 Ga dolomitic rocks in the Isua supracrustal belt (SW Greenland) as biogenic conical and domical stromatolites grown in a shallow marine environment. These interpretations were challenged by Allwood et al. (2018) and Zawaski et al. (2020), who reinterpreted the structures as likely products of compressional deformation induced by tectonic activity. In the field, Allwood et al. (2018) observed convex-up and convex-down stromatoloid growth orientations, which, they argued, precluded an interpretation of growth in a shallow marine environment. Additionally, they described “extreme elongation” of the three-dimensional structure, where the stromatoloids, “…are not cones or elongate cones, but ridges extending at least 10 cm (our sampling depth) into the rock … that is consistent with a tectonic origin.” They also described the texture-specific distribution of major and minor elements and the REE + Y composition as “consistent with original deposition in a marine environment, followed by secondary carbonate alteration.” Nutman et al. (2019) argued that despite intense metamorphism, the stromatolites were preserved in a rare, low-strain fold hinge, as indicated by the presence of both dolomite and quartz that had not reacted to form tremolite. They also indicated that the stromatoloids were unlikely to be “flame structures” produced by soft-sediment deformation due to their height relative to the thickness of the source beds, the presence of internal laminations, their relationship to onlapping sediment, and their broad, domed tops. Additionally, the authors presented geochemical data that indicate the dolomite was not a product of secondary alteration and instead retained a seawater-like REE+Y signature. Through geologic mapping and sample microanalysis, Zawaski et al. (2020) challenged the occurrence of internal layering, as would be expected in true stromatolites. They demonstrated via electron backscatter diffraction that the structures experienced similar levels of deformation to the surrounding area and were instead consistent with “boudinage” formed by tectonic processes. As illustrated in this example, proving a biogenic origin for sedimentary structures can be exceptionally challenging, particularly among sediments that have experienced significant alteration. We emphasize that if similar structures were observed on Mars, morphological observations alone would likely be insufficient to conclusively determine biogenicity, and that multiple lines of evidence and knowledge of the geological context of the deposit would be required to better decipher possible origins. Additional discussion with regard to the interpretation of potentially biogenic laminated structures is given in Section 4.1.
On Mars, the lack of plate tectonic development may have limited widespread recycling and destruction of crustal rocks and thus may have enhanced the potential for long-term preservation of primary sedimentary structures (Westall et al., 2015a). However, this also means there were fewer mechanisms to uplift and expose buried sediments for investigation. Other processes such as high cosmic ray and UV fluxes at Mars’ surface have likely contributed to the degradation of organic carbon in the upper surface over geologic timescales (Pavlov et al., 2012). Future exploration activities that integrate subsurface investigation, for example, via subsurface drilling by the planned European Space Agency Rosalind Franklin Rover (Vago et al., 2017), may provide the most promising opportunities yet to search for laminated structures that retain carbonaceous remnants potentially representing evidence of life. Ultimately, however, a Mars Sample Return (MSR) mission will be necessary to distinguish possible signs of life using sophisticated laboratory instrumentation with sufficiently high spatial resolution and analytical precision (Meyer et al., 2022; Kminek et al., 2022; c.f., Mars Sample Return Special Issue, Astrobiology, Volume 22, Issue S1).
Laminae as Features of the LDKB
LDKB structure and language
The LDKB is a tool designed to help collate and present information on a wide range of possible biosignatures that may be detected beyond Earth. Members of the scientific community contribute to the database by providing arguments and evidence from the peer-reviewed literature that describe potential biosignatures as well as features that may confound biogenic interpretations. To coherently present information about disparate biosignature categories in a standardized format (e.g., isotopes, lipids, laminae, technosignatures, and more; Hoehler et al., 2025; Pohorille et al., 2025; Davila et al., 2025; Shkolyar et al., 2025; Des Marais et al., 2025; Perl et al., 2025), the LDKB employs a formal organizational structure and language (Fig. 7). Each piece of evidence presented in the LDKB and its supporting arguments are organized into two main categories: prevalence and signal strength, described below.
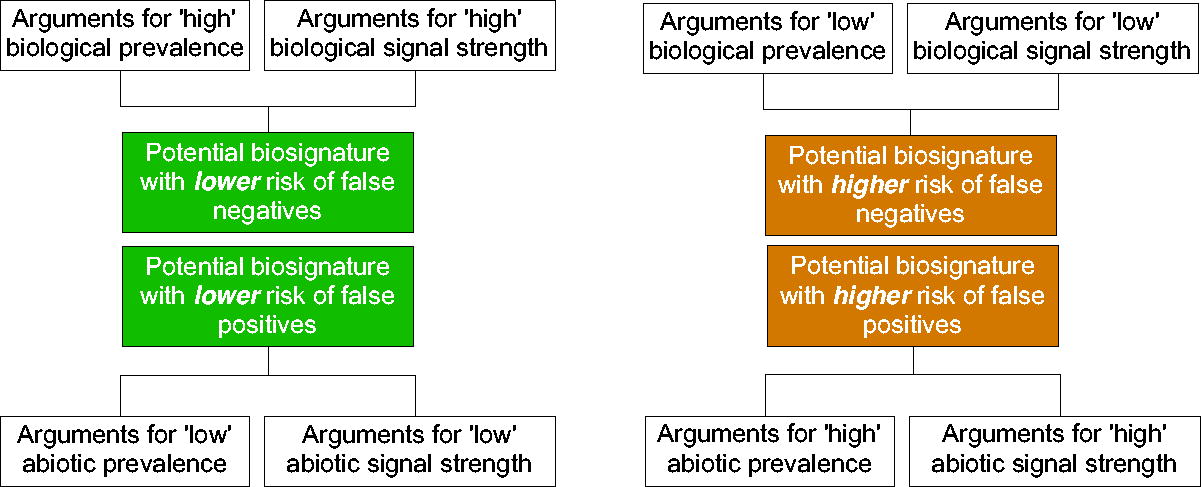
Schematic representation of the relationship between criteria, “arguments,” and false positive/negative potential within the Life Detection Knowledge Base (LDKB). Knowledge sourced from peer-reviewed publications is represented in the LDKB in the form of user-created “arguments” that are organized according to whether they (i) relate to biological or abiotic sources of a given potential biosignature, (ii) concern prevalence or signal strength, and (iii) argue for high versus low prevalence or signal strength. Arguments can be formulated in environment- or context-dependent terms.
Prevalence expresses the likelihood that a feature is produced by a particular source, such as by life or a nonbiological process. “Prevalence” in epidemiology indicates the proportion of a population that has a specific characteristic (e.g., a disease; Krug and McNutt, 2008). Nominally, “prevalence” in astrobiology applies across all biological systems and all environments; therefore, it includes all occurrences of a feature on Earth and beyond. The implication is that an argument about general prevalence is essentially independent of environmental context and rather about the intrinsic aspects of a feature. However, some features, such as biogenic laminae that are found among different types of sedimentary rocks and/or precipitates, are expected to be prevalent in particular environments and unlikely in others. Therefore, it should be specified whether the prevalence of a feature is considered in the context of all possible environments or only specific environmental settings. When a feature is specific to its environmental context, the evaluation of its prevalence in that particular setting becomes an evaluation of its “congruence.”
Signal strength
Signal strength is the prominence of a feature (e.g., its abundance, rate, structure, patterns, and intensity) that provides evidence for the past or extant presence of life or a nonbiological process. The prominence of a feature is determined by the balance between its rates of production and its rates of modification and/or destruction. For a biosignature, this balance is determined by its biological production, its survivability, and the patchiness of its occurrence in a target environment. The “target environment” includes the environment that prevails during either feature production or its modification, degradation, and/or destruction. These environments may be different (separated by time and/or space).
Arguments and evidence in the LDKB
In the following sections, we discuss example LDKB entries of laminae with emphasis on examples found in Precambrian rocks from the Barberton greenstone belt, South Africa, and from the Pilbara, Western Australia. The entries discussed relate to three types of structural observations that might be made by a current or future planetary exploration mission, namely (1) “size distribution,” (2) “spatial distribution,” and (3) “texture and fabric.” These measurable parameters are applicable for all potential “structural” biosignatures in the LDKB (see Shkolyar et al., 2025). These broad categories were determined with reference to well-established geological terminology and are intended to be sufficiently “agnostic” to describe a given visual feature of interest.
Biotic prevalence arguments for laminae
The following examples describe LDKB entries that address the prevalence of laminae that are generally considered attributable to the presence or influence of microbial cells, populations, or communities, as supported by evidence presented in the scientific literature. The greater or higher prevalence that favors true positive biosignature detections is presented below as “HIGH” arguments. In general, arguments are made in reference to the potential prevalence of such features as they may exist on Mars.
Evidence—Rip-up and roll-up structures co-occurring with carbonaceous material serve as evidence for biogenicity. In the 3.42 Ga Buck Reef Chert (Barberton greenstone belt), fine laminae deposited in open-marine volcanic platform facies contain ripped-up, plastically deformed, rolled-up fragments and occur within black bands in banded chert. These fabrics, consistent with cohesion displayed by modern microbial mats, are interpreted as remnants of microbial mats (Simonson and Carney, 1999; Tice and Lowe, 2004). The fine laminae in these rip-up structures incorporate and loosely drape detrital grains and occur as silica-filled lenses. They also form fine meshworks of filament-like strands <5 µm in diameter that drape over detrital grains. “Gamma-type” laminae display even-spaced, flat laminae that tightly drape underlying sediments and exhibit lightly fractionated δ13Corg values consistent with a biological origin (see also the LDKB entry under “Chemistry”—isotope ratio patterns: carbon redox states; Des Marais et al., 2025).
Evidence—Rip-up and roll-up structures interpreted as microbial mat remains are widespread in sedimentary environments on early Earth. They have been identified in the 2.5 Ga Hamersley Group (Simonson and Carney, 1999); ∼2.5 Ga Campbellrand Subgroup (Gandin et al., 2005); 3.22 Ga Moodies Group (Noffke et al., 2006; Homann et al., 2015); 3.48 Ga Dresser Formation, Pilbara, Western Australia (Noffke et al., 2013); 3.33 Ga Josefsdal Chert (Westall et al., 2015b); 3.46 Ga stratiform “Apex chert” (Hickman-Lewis et al., 2016); and 3.47 Ga Middle Marker horizon (Hickman-Lewis et al., 2018), where geometries and geochemical signals coincide and support biogenicity.
Evidence—Precambrian laminae from the Barberton greenstone belt occur as horizontally continuous packets and display macro- and microtufted geometries. Homann et al. (2015), Hickman-Lewis et al. (2018), and Hickman-Lewis and Westall (2021) argue that the microtufted spatial organization observed in the 3.22 Ga Moodies Group and 3.47 Ga Middle Marker horizon is consistent with growth phenomena associated with phototrophic microbial mats, namely the accumulation of gases produced as a result of microbial metabolic activity as shown in modern microbial mats (e.g., Bosak et al., 2010).
Evidence—Microtufted laminae occur in the 3.48 Ga Dresser Formation, Pilbara, Western Australia. The tufts have a similar height/base ratio of 10/50 to 25/75 µm, arranged at regular distances of 100–125 µm from each other. Raman analyses showed a composition of pyrite and relic carbonaceous matter via peaks at 1350 cm−1 (D1 disordered peak) and 1600 cm−1 (the G graphite peak). The positions and widths of these peaks and the D1/G peak heights are consistent with maturation related to the metamorphic grade of the dresser formation. Based on their morphological similarity to modern tufted microbial mats and carbonaceous composition, these features are interpreted as biogenic microbial mat remains (Noffke et al., 2013).
Evidence—In the Moodies Group, Barberton greenstone belt, chert-filled cavities in the interior of tufts were interpreted to represent silicified gas bubbles that were trapped within a microbial mat fabric (Homann et al., 2015; Homann et al., 2016), as they are common features in cyanobacterial mats that produce oxygen-rich bubbles (Bosak et al., 2010).
Evidence—Tufts with gas bubbles occur in the 3.48 Ga Dresser Formation, Pilbara, Western Australia (Noffke et al., 2013), and within stromatolitic laminae in the 2.7 Ga Ventersdorp Formation, South Africa (Wilmeth et al., 2019, 2022). Gas bubbles concentrated along laminae in tufts are interpreted to form due to the upward migration of biogenic gases such as O2 and H2S (Noffke et al., 2003).
Evidence—Laminae occurring within the Moodies Group, Barberton greenstone belt, drape and stabilize underlying sandstones and are enriched in fine-grained quartz and feldspar, as well as heavy mineral grains (zircon and rutile or anatase), interpreted to be caused by microbial baffling and trapping (Homann et al., 2015, 2018); such microbe–sediment interactions are commonly observed in epibenthic microbial mats (Gerdes et al., 2000). Laminae here are also enriched in organic material and include “oriented grains” interpreted to be derived from the substrate beneath a microbial mat that was dragged upward during mat growth (Noffke et al., 2006, 2024). In addition, the carbonaceous material exhibits carbon and nitrogen isotope distributions that are consistent with a biological origin (Noffke et al., 2006; Homann et al., 2018).
Evidence—Laminae are composed of carbonaceous materials in the Middle Marker horizon, Barberton greenstone belt, drape underlying sedimentary layers, and exhibit petrographic evidence for grain trapping (Hickman-Lewis et al., 2018). This grain-trapping behavior has also been detected in three-dimensional renderings of microbial mat horizons, where organic laminae enwrap denser mineral particles (Hickman-Lewis et al., 2019).
Signal strength arguments for laminae
The following examples describe LDKB entries that address the signal strength of laminae, or the prominence of a feature as determined by the balance between its rates of production and its rates of modification and/or destruction. The arguments below present a useful framework to recognize that laminae as structural features may persist on planetary surfaces for billion-year timescales and may be laterally extensive at kilometer scales. Here, greater signal strength that favors true positive biosignature detections is presented as “HIGH” argument. “LOW” arguments indicate weaker signal strength, which increases the probability of false negative detections.
Evidence—Laminae within the Middle Marker horizon (3.472 Ga) of the Barberton greenstone belt that have been interpreted as fossil microbial mats occur as packets with a thickness of 200 µm to 2.5 mm (Homann et al., 2015, 2018; Hickman-Lewis et al., 2018, 2019).
Evidence—Within the Buck Reef Chert (3.416 Ga) of the Barberton greenstone belt, laminae interpreted as microbial mat remains occur as fine, anastomosing (i.e., branching and merging) carbonaceous laminae (cf., Tice and Lowe, 2004; Tice, 2009; Greco et al., 2018; Hickman-Lewis et al., 2022; Pellerin et al., 2023; Reinhardt et al., 2024).
Evidence—Within the Josefsdal Chert (3.33 Ga) of the Barberton greenstone belt, wavy carbonaceous laminae that entrain fine-grained detrital volcanic clasts and quartz grains occur within banded black-and-white cherts as layered packets. These features are interpreted as microbial mat remains (Westall et al., 2006, 2011, 2015b; Homann, 2019).
Evidence—Black-and-white cherts of the 3.26 Ga Mendon Formation (Barberton greenstone belt) preserve layered packets of laminae formed of thin, wavy carbonaceous laminae up to several tens of micrometers in thickness (Trower and Lowe, 2016).
Evidence—The presence of 0.5–1 mm thick crinkly laminae within the Saddleback Syncline in the Moodies Group (∼3.22 Ga) of the Barberton Greenstone Belt is interpreted as an example of siliciclastic tidal mats (Noffke et al., 2006; Heubeck, 2009; Gamper et al., 2012; Homann et al., 2015). Terrestrial (fluvial) environments within the Moodies Group also preserve laminae interpreted as the fossilized remains of microbial mats with thicknesses of up to 4 mm (Homann, 2019). Similar laminae occur in sandy tidal flat horizons of the 3.48 Ga Dresser Formation, Pilbara, Western Australia (Noffke et al., 2013, 2024).
Evidence—Numerous horizons of the Campbellrand–Malmani carbonate platform sequence are associated with layered carbonaceous material and form bifurcating anastomosing laminae that locally form more complex fenestral (cavity-forming) stromatolitic mesostructures (Sumner, 1997; Gandin et al., 2005).
Evidence—Within the Precambrian Moodies Group on the Barberton greenstone belt, fossil mats are laterally traceable for ∼15 km in a ∼1000 m thick succession in the lower part of the Saddleback Syncline and show distinct morphological adaptations to different hydrodynamic settings: (1) planar-type in coastal floodplain, (2) wavy-type in intertidal, and (3) tufted-type in upper inter- to supratidal facies (Homann et al., 2015).
Evidence—Archean MISS that include laminae consistent with microbial origin are described from numerous sites that include the 3.48 Ga Dresser Formation, Pilbara, Western Australia (Noffke et al., 2013, 2024); the 3.2 Ga Moodies Group, Barberton Greenstone Belt, South Africa (Noffke et al., 2006); and the 2.8 Ga Pongola Supergroup, South Africa (Noffke et al., 2003, 2008).
Evidence—The fossilized remains of 1–4 µm thick biofilms are exposed on fresh fractured bedding surfaces of the Josefsdal Chert in the Barberton greenstone belt (Westall et al., 2001, 2006, 2011). Scanning electron microscope observations reveal the presence of multiple layers of parallel filament-like structures that exhibit consistent diameters of 0.25 µm embedded in a granular-to-smooth film interpreted as EPS (Westall et al., 2006, 2011).
Evidence—Filamentous microstructures of carbonaceous composition were identified in a single 2 cm thick black-and-white banded chert layer in the upper Hooggenoeg Formation of the Barberton greenstone belt (Walsh and Lowe, 1985; Walsh, 1992). These thread-like or cylindrical, unbranched filaments are 0.2–2.5 µm in diameter, up to 200 µm long, occur in association with carbonaceous laminae, and are interpreted as microbial mats (Homann, 2019).
Evidence: Filaments associated with fossil mats occur in several localities of the Buck Reef Chert of the Barberton greenstone belt and consist of hollow cylindrical filaments (1.2–1.4 µm in diameter and 10–150 µm in length) that are composed of carbonaceous matter and fine-grained pyrite, with most oriented subparallel to bedding or sometimes as interwoven, tangled clumps (Walsh and Lowe, 1985; Walsh, 1992; Homann, 2019).
Evidence—Ancient (∼3.42 Ga) filamentous microfossils discovered in the Barberton greenstone belt occur in specific associations (single or in clusters) within different parts of vein microhabitats and in association with biofilms (Cavalazzi et al., 2021).
Evidence—Putative microfossils in the carbonaceous cherts of the Onverwacht Group are extremely rare. The systematic analysis of such materials by Walsh (1992) and Walsh and Lowe (1999) revealed that only 9 of >400 analyzed samples contained possible microfossils of filamentous, lenticular, and spheroidal shapes (Homann et al., 2016).
Abiotic prevalence and signal strength arguments
In addition to presenting evidence and arguments for a variety of biogenic features, the LDKB contains information about abiotic features that mimic those produced by biology. Below are some examples of abiotic prevalence and signal strength arguments that may contribute toward higher false positive identifications of signs of life. Note: Arguments below are classified as “HIGH” or “LOW” with respect to the potential to identify such features on Mars.
Evidence—Laminated stromatolite-like or stromatoloid structures within the Isua supracrustal belt from southwest Greenland have been interpreted as being of biological, stromatolitic origin (Nutman et al., 2016, 2019; Van Kranendonk et al., 2025), but they occur in metamorphosed and deformed rocks and are interpreted by others as deformational features (Allwood et al., 2018; Zawaski et al., 2020).
Evidence—Pseudocolumnar and cylindroidal deformation structures that contain laminae that strongly resemble stromatolitic laminae occur in the Upper Onverwacht and Fig Tree groups (de Wit et al., 1982); however, detailed observation shows that these structures are medusoid-like surface impressions that are more consistent with abiotic formation in mudpools associated with hydrothermal activity.
Evidence—Stromatoloid structures produced during laboratory spray deposition experiments suggest that similar structures in environments characterized by spray deposition, for example, splash pools, silica sinter-forming settings, and desert varnish crusts, may also be formed by abiotic processes (McLoughlin et al., 2008), though it is worth noting that these experiments produced similar 2D rather than 3D structures and that modern silica sinters with such shapes (in 2D and 3D) contain remnants of fossilized microbial biofilms along lamination boundaries and within sinter laminae (Cady and Farmer, 1996; Handley et al., 2008).
Evidence—Cavities within tufts may form by tidally driven hydraulic pumping of the ambient air trapped in pore space (Homann et al., 2016). Such structures may be misconstrued as biological in origin in the absence of further information, for example, in situ geochemistry within the tufts.
Evidence—Pseudo-tufted structures that occur up to 0.8 mm in height within the Middle Marker horizon, Barberton greenstone belt, may form abiotically as a result of plastic deformation caused by the settling of dense particles (Hickman-Lewis et al., 2018). Such structures may be incorrectly interpreted as fossilized (micro-)tufted microbial mats.
Evidence—McMahon (2019) demonstrated the growth of abiotic filamentous structures in laboratory experiments that mimicked plausible natural geochemical environments.
Evidence—Experiments by García-Ruiz (2000) using alkaline fluids produced “biomimetic” microstructures of abiotic origin.
Evidence—McMahon et al. (2021) described filaments associated with laminae in Jurassic mineralized veins formed in association with serpentinization. Here, filamentous structures are recognized as “dubiofossils” of uncertain origin, which may have formed by abiotic processes such as those described in laboratory experiments.
Evidence—In the Kromberg Formation of the Barberton greenstone belt, solid carbonaceous spheroids and ellipsoid structures were interpreted to potentially represent abiotic structures resembling microbial reproductive structures (Walsh, 1992), and that “some of the small spheroids have cross-sections that are slightly rhombic, suggesting that they may have formed in association with original carbonate minerals.”
We acknowledge that the examples provided above are not exhaustive and may omit important cases from the literature. We encourage members of the scientific community to contribute to the LDKB and help expand and refine the range of arguments and evidence related to potential biogenic and abiogenic features relevant to astrobiological exploration.
Discussion
Considerations for interpreting laminae as potential biosignatures in the context of planetary exploration
Given our Earth-centric understanding of life, it is important to recognize that any discovery of potential extinct or extant evidence for life beyond Earth must be supported by the occurrence of multiple potential biosignatures in a plausibly habitable setting. It is crucial that future astrobiology exploration missions include multiple instruments capable of characterizing, at a minimum, the morphological attributes and chemical composition of potential biosignatures at relevant spatial scales (e.g., from the centimeter to submicrometer scales for individual laminae) and result in sample return efforts where materials can be analyzed with state-of-the-art instruments in well-equipped laboratories on Earth. The search for life requires stringency and consistency in its methodologies and an understanding of the variety and types of evidence that will be useful to determine the origins of potentially biogenic features. In light of these requirements, we outline below key considerations for interpreting laminae as potential biosignatures in the context of planetary exploration:
Microbially influenced laminae retain important information about detrital and/or chemically precipitated components at the microscale. As discussed in Reid et al. (2024), biogenic sedimentary laminae form as a result of a balance between extrinsic and intrinsic forces, which result in morphological characteristics that can be preserved over geologic timescales. To have the best chance of resolving the origins of laminated structures suspected to have a biogenic origin, it is crucial that they be examined in detail at the microscale, where observations of grain size, mineralogy, orientation, and composition can be characterized to better understand the potential roles of extrinsic and intrinsic processes. Spaceflight missions focused on characterizing evidence of past life should be capable of resolving these features using imaging and chemical mapping techniques. The combination of the SHERLOC (including the WATSON camera) and PIXL instruments on the Mars 2020 Perseverance rover is an example of such an instrument combination capable of resolving sub-millimeter rock textures and their compositional variation (Allwood et al., 2020; Bhartia et al., 2021; Hickman-Lewis et al., 2022).
Physiochemically formed laminae may preserve biotic or abiotic organics. Uniform laminated sediments formed principally by abiotic processes (as described in Section 2.2) can still retain biosignature material, especially those that contain significant clay fractions that readily adsorb organics. Though the structural character of laminae in these cases may not be initially indicative of possible microbial activity, they may still serve as repositories of biosignatures concentrated from the wider sedimentary basin. However, given the continual flux of meteorite impacts on planetary surfaces throughout their geologic history (e.g., Lagain et al., 2022), it is likely that carbonaceous chondrites delivered significant amounts of abiotic organic materials that, through reworking and transport, were eventually deposited in sedimentary basins. On Earth, compelling evidence for impact events is recorded in the strata of the Barberton greenstone belt and Pilbara craton (Lowe et al., 1989; Glikson et al., 2004, 2016; Koeberl et al., 2015; Lowe and Byerly, 2024); one such horizon preserves geochemical evidence for extraterrestrial carbonaceous materials (Gourier et al., 2019). Thus, an exogenous abiotic origin for the presence of carbonaceous material in sediments on Mars must be considered.
Metamorphic and diagenetic processes create irregular lamination geometries. Several syn- and postdepositional processes can form irregular laminae that resemble those produced by biofilm communities (discussed in Section 2.2.5). To assess their potential biogenicity, it is first crucial to characterize the wider geological context of the studied area. The depositional paleoenvironment should be understood through a facies-based framework; this can be useful not only to estimate the potential habitability of environments but can also help identify paleoenvironments susceptible to alteration, for example, by soft-sediment deformation common near the steep slopes of subaqueous platforms and shelves (Tinterri et al., 2016). Such environments may be especially common in chaos terrains on Mars, where subaqueous landslides and mass flows are posited to have occurred (e.g., Nizam et al., 2022). At smaller scales, proximate features may indicate loading stresses, such as disruptions to laminae by sand lenses, which signify pressurized injection into surrounding strata (e.g., Brasier et al., 2019). Where possible, potentially biogenic laminated structures should also be investigated with regard to their three-dimensional orientations in space to better characterize their geometry in relation to known biogenic structures.
Structural characteristics of laminae alone will likely not be sufficient to determine biogenicity on Mars. Measurements of lamination thickness, orientation, and lateral extent alone are unlikely to provide definitive evidence for the biological origin of laminated structures. In studies of preserved microbial biosignatures on Earth, claims of biogenicity are bolstered by multiple lines of evidence, for example, (1) their occurrence in a habitable environment, as interpreted from local geologic context; (2) the presence of microfossils and structural organization; (3) organic carbon in the form of insoluble kerogen and/or soluble biomarkers (e.g., lipids and their diagenetic products); and (4) isotope fractionation patterns consistent with metabolic activity (e.g., Buick et al., 1981; Noffke, 2000; Awramik and Grey, 2005; Schopf et al., 2007; Wacey, 2009; Wagstaff and Corsetti, 2010; Homann et al., 2018; Homann, 2019; Lepot, 2020).
It is important to note that on Earth, however, microfossils are rarely preserved among laminated stromatolitic structures (Schopf, 2012). This may be due to alteration caused by regional tectonic activity or, on shorter timescales, caused by destructive diagenetic effects that operate syn-depositionally or immediately after stromatolite formation (e.g., Lima-Zaloumis et al., 2024). However, structural elements may still be useful to help interpret the origins of suspected biogenic structures. For example, Hickman-Lewis et al. (2022) demonstrated an approach that considered two- and three-dimensional structural observations at the microscale (via synchrotron computed tomography) to interpret the origins of ca. 3.5 Ga stromatolites from the Dresser Formation, Pilbara, Western Australia. In examined samples, they observed onlapping relationships consistent with stromatolite growth, thickening of laminations toward crests, nonisopachous laminations that lack morphological inheritance from underlying layers, the presence of structural elements resembling microbial palisade structure, and the presence of void spaces consistent with fenestrae. The occurrence of these elements together serves as mutually reinforcing evidence to support the hypothesis that such structures were formed by biological processes.
Existing knowledge gaps
By viewing and contributing content to the LDKB, knowledge gaps in biosignature research can be readily identified. Through the development of the LDKB’s initial content, we have recognized several directions for future research on laminated structures as potential biosignatures, described below.
Details of how extant microbial mats create successive mineral layers are not well explored. As indicated by Reid et al. (2024), “detailed study of environmental characteristics on multiple scales (diurnal to seasonal) must be integrated with investigation of accretion rate and initial architecture, achieved through petrographic, biogeochemical, and metabolomic approaches.” Additionally, although many important insights are gained from the study of natural microbialite growth, further laboratory experimentation and modeling studies (e.g., Dupraz et al., 2006) are warranted to explore the various parameters that influence microbial mat lithification and upward growth, such as community composition, mineral saturation states, and the role of physiochemical environmental processes, such as evaporation and degassing, and the resultant changes in fluid composition.
Biologically influenced lamination formation is poorly studied in noncarbonate systems. To date, the dynamics between microbial mat growth and lithification have been best explored in carbonate-precipitating systems (for good reason, as carbonate beds are by far the most common host of stromatolites and other microbialites). However, these dynamics should be further explored in systems that exhibit dominant precipitation of minerals such as silica, sulfate, and iron and in a range of environments, such as epicontinental basins, lakes, hot springs, evaporative systems, and subaerial settings.
Diagenetic processes that influence the long-term preservation of biogenic laminae are not well understood. The detection of biogenic sedimentary laminae beyond Earth depends principally on (A) whether microbial organisms existed that could form lithified laminated structures (as they are known to on Earth) and (B) whether those structures were preserved in the rock record by favorable taphonomic processes (Farmer and Des Marais, 1999). It remains unclear, however, how biologically induced laminae, along with their associated biogeochemical signs of life, may become altered or lost throughout early-to-late-stage diagenesis. For example, the relative importance of early postmortem mineralization, short- and long-term diagenesis, and metamorphism should be evaluated experimentally, by modeling, and through relevant field studies.
Abiotic mechanisms that form irregular laminated structures resembling microbial mat remains are not sufficiently understood. Abiotic mimics of biological structures have been demonstrated to form under laboratory growth conditions and have been reported in natural environments (e.g., Davies et al., 2016; McMahon and Cosmidis, 2022). To date, few studies have investigated the occurrence of abiotic mimics that resemble microbialites (e.g., McLoughlin et al., 2008). Experimental studies are warranted to explore how such structures form through abiotic processes and what parameters (e.g., fluid energies, grain composition and size, and geochemical conditions) result in the formation of cohesive mat-like structures. Such investigations with an eye toward sedimentary environments on Mars are likely to be highly informative for understanding the potential for false positive detections.
Additional strategies should be developed to discriminate between biotic and abiotic formation origins of various lamination geometries. As discussed in Section 2.2.5, many lamination geometries have a high degree of overlap between biogenic and abiogenic forms. Thus, additional methods should be developed to improve the discernment of true from false positives. For example, quantitative multiscale observations (e.g., regional to micron) can be integrated to refine models of stromatolite morphogenesis to better discern their origins from abiotic counterparts (e.g., Ibarra and Corsetti, 2016). Additionally, detailed morphometry analyses (e.g., Suarez-Gonzalez et al., 2014) and assessments of complexity between biogenic and abiogenic forms (e.g., Corsetti and Storrie-Lombardi, 2003; Wagstaff and Corsetti, 2010) may also contribute to useful strategies for source discrimination. Additionally, new methods of discrimination driven by machine learning tools may also prove useful to identify enigmatic patterns among high-dimensionality datasets, which could reflect biogenic and/or abiogenic source processes.
Conclusions
Throughout the astrobiological and paleontological literature, researchers have utilized a wide variety of methods to characterize morphological signs of life in terrestrial samples. The development and application of ever more advanced laboratory instrumentation, miniaturized flight instruments, and analytical methods and protocols (including data treatment) are likely to be driven by the search for life at the surface and near-surface of Mars and other bodies of astrobiological significance, as well as among samples returned to Earth. The LDKB represents a growing repository of key observations required to interpret possible signs of life in planetary analog materials through the use of existing instruments. By viewing and contributing to the LDKB, researchers may identify gaps in life detection capabilities and strategies and may thereby guide the development of new tools suited to diverse geological settings and materials.
Here we have detailed how laminae can serve as key features of interest in astrobiological exploration activities beyond Earth, and how they are featured in the LDKB. Though laminae can form by a wide range of processes, many of them are influenced by the activity of microorganisms. On Earth, evidence of microbial activity among laminated sediments has been preserved over geological time. Laminated structures therefore represent potential targets of interest for current and future space missions focused on life detection on rocky planets that host aqueous environments. Though we have described only a few of the structural attributes of laminae that may be consistent with a biological origin, such observations must be supported by additional data, particularly by their composition and environmental setting, to better enable robust interpretations about their biogenic origin.
Footnotes
Author Disclosure Statement
The authors declare they do not have conflicts of interest.
Funding Information
Support for this work was provided by NASA’s Planetary Science Division Research Program, through the Center for Life Detection NASA Internal Scientists Funding Model (ISFM) package at Ames Research Center. S.S. acknowledges support from NASA under award number 80GSFC24M0006. B.C. acknowledges support from Agenzia Spaziale Italiana and ministry of university and research (grant I53D24000060005). K.H.L. acknowledges funding from the UK Space Agency (grant nos. ST/V00560X/1 and ST/Z000491/1).
Authors’ Contributions
All authors contributed to writing the manuscript and provided LDKB argument examples with corresponding evidence.
Abbreviations Used
Associate Editor: Christopher P. McKay