
Abstract
Abstract
Breast cancer that develops during or shortly after pregnancy is frequently more aggressive than cancer diagnosed at other times in a woman's life. To better understand the patterns of cancer-related protein expression in the breasts of lactating women, we determined the differences in total and individual protein expression in milk based on (a) three time points during lactation (early, mid, and late), (b) length of lactation, and (c) parity. Breastmilk was collected from 72 healthy lactating women within 10 days of starting lactation (transitional [T]), 2 months after lactation started, and during breast weaning (W). Sixteen proteins whose expression is altered in breast cancer (11 kallikreins [KLKs], basic fibroblast growth factor [bFGF], YKL-40, neutrophil gelatinase-associated lipocalin, and transforming growth factor [TGF] β1 and β2) were evaluated. The concentration of total milk protein decreased over time (p<0.01 at 2 months and W compared with T). After we controlled for total protein, KLK6 and TGFβ2 significantly increased, and bFGF decreased from T to W. Neither length of nursing nor parity significantly influenced individual protein expression at the W time point. On the other hand, length of nursing did influence the difference in KLK6, -7, and -8 expression between the W and T time points. Total milk protein concentration is lower in the mid and late phases of nursing. Biomarker differences between T and W milk samples in KLK6, TGFβ2, and bFGF are consistent with a protective effect of nursing.
Introduction
The concentration of total milk protein influences the concentration of individual proteins in milk. The length of lactation and the number of children born, or parity, vary from woman to woman. Our hypothesis was that one or both of these variables influenced mature milk protein concentration. There are conflicting reports on the possible change in total protein concentration of mature milk related to length of lactation. Total protein concentration of mature milk was evaluated at 3, 6, 9, and 12 months after the start of lactation. 5 Protein concentration decreased at 6 and 9 months (compared with 3 months) but then increased at 12 months (Table 1). The authors did not find an association between protein intake by the mother and milk protein concentration. In a separate study (Table 1), milk protein concentration was slightly lower at 14 months compared with 3 months of nursing. 6
Concentration data are in g/dL.
At given months from birth or from the onset of lactation.
The concentration of total milk protein does not appear to be influenced by parity through 9 months of nursing5,7,8 but was influenced at 12 months in at least one report. 5 Less is known about the expression of individual cancer-related proteins in breastmilk based on length of lactation and parity.
We have identified cancer-associated proteins in breast nipple aspirate fluid, fluid from the milk ducts of nonlactating women, including kallikreins (KLKs), basic fibroblast growth factor (bFGF), 9 and YKL-40 (also known as chitinase 3-like 1). 10 The human KLK family consists of 15 highly conserved serine proteases. Several members of the family have been reported as potential cancer biomarkers, 11 and the genes for many KLK proteins are differentially expressed in hormone-related malignancies. 12 It has previously been demonstrated that multiple KLKs are detectable in breastmilk. 12 bFGF 13 has been detected in human milk. YKL-40 (and its bovine homolog, mammary gland protein-40) levels in breastmilk rise during the time of breast involution, as the breast returns to the prepregnancy state. 14
Neutrophil gelatinase-associated lipocalin (NGAL) (also known as lipocalin 2), a small, secreted glycoprotein that binds matrix metalloproteinase-9 to protect it from degradation, 15 is expressed in breast carcinomas. Inhibition of NGAL impairs breast tumorigenesis and metastasis, 16 and increased NGAL expression has been associated with decreased disease-specific survival. 17 The homolog of NGAL has been identified in cow's milk. 18
Transforming growth factor (TGF) β is linked to breast cancer in preclinical models 19 and to prognosis in human breast cancer. 20 In mouse models, TGFβ is induced during mammary gland lactation and involution, 19 and it mediates pro-apoptotic effects during involution. 21 Transgenic mice that overexpress TGFβ show increased apoptosis in the mammary epithelium throughout mammary development. 22
Three factors that have been reported to influence breast cancer risk are breast involution (initiated by breast weaning, in the late phase of lactation), parity, and length of nursing. Our strategy was to determine the differences in total and individual protein expression in milk based on parity, as well as phase and length of lactation.
Subjects and Methods
Recruitment
Healthy women were prospectively recruited prior to or soon after delivery. They were eligible if they delivered an infant ≥37 weeks gestational age and were planning on breastfeeding. After subjects agreed to enroll in an Institutional Review Board–approved project, three milk samples were requested from each participant: transitional (T), defined as within 10 days of the initiation of lactation; 2 months after lactation started (mature milk); and when the woman was weaning (W). We asked the participant to collect the W sample relatively soon after initiating the weaning process, in order to insure that a sample was easily collected. The participant was asked to provide the wean sample once they had decided to stop nursing and had started to decrease the number of daily feedings. At least two samples from a given woman were required in order for the samples to be included in our analyses. Each sample was collected from the same breast.
After Institutional Review Board approval, subjects were recruited in Columbia, MO, and Grand Forks and Fargo, ND. Mothers were asked not to breastfeed their infants for at least 2 hours before milk collection, which involves draining the breast using manual extraction or a breast pump. Samples were immediately frozen after collection. Collected milk was thawed and centrifuged (1,500 g, 20 minutes, 4°C), and the fat and cellular layers were separated. The aqueous phase was then centrifuged at 12,000 g for 15 minutes at 4°C, and the second lipid layer was removed and stored at −80°C prior to analysis.
Assessment of biomarkers
Total protein
Twenty-five microliters of standard and milk samples were added in duplicate to each microplate well (Pierce, Rockford, IL), followed by 200 μL of working reagent with mixing. The microplate was then incubated at 37°C for 30 minutes, and absorbance was measured at 562 nm.
KLKs
The concentration of KLKs was measured with a highly sensitive and specific noncompetitive immunoassay. Each assay incorporated two KLK-specific monoclonal and/or polyclonal antibodies, raised in mice and rabbits, respectively. One antibody was used for coating and the other for detection in a sequential two-site immunometric format with time-resolved fluorescence detection. Many publications have used these assays.12,23–27 KLK2 and KLK3 antibodies were obtained from MedixBiochemica (Kauniainen, Finland). The remaining KLK antibodies were developed in the laboratory of Dr. Eleftherios Diamandis, Director of Clinical Biochemistry, University of Toronto, Toronto, ON, Canada. The KLK assays have a detection limit ranging from 0.005 μg/L for KLK2 to 2 μg/L for KLK12 and a dynamic range of 10–20 μg/L, depending on the KLK. Standards (recombinant KLK) and samples were analyzed in duplicate with inclusion of three quality control samples in every run.
bFGF
bFGF was analyzed using an enzyme-linked immunosorbent assay kit from R&D Systems (Minneapolis, MN), following the manufacturer's instructions. The kit uses a quantitative sandwich enzyme immunoassay technique. The detection limit of the kit is 10 ng/L.
YKL-40
Samples were analyzed by immunoassay as per the manufacturer's instructions (Quidel Corp., San Diego, CA). The kit uses a monoclonal anti-YKL-40 antibody conjugated to biotin that binds to streptavidin and captures YKL-40 in the standard or sample. The detection limit is 10 ng/mL.
NGAL
NGAL content in milk was measured by immunoassay (BiPorto Diagnostics, Gentofte, Denmark) according to the manufacturer's instructions. In brief, 100 μL of standard and diluted milk samples was added to wells coated with a monoclonal antibody against human NGAL. Bound NGAL was detected with a second antibody labeled with biotin, the signal was developed with horseradish peroxidase–conjugated streptavidin and substrates, the reaction was ended with stop solution, and absorbance was measured at 450 nm. The detection limit of the NGAL kit is 4.0 pg/mL.
TGFβ isoforms
The protein expression levels of TGFβ1 and β2 in milk samples were determined by immunoassay (R&D Systems) following the manufacturer's instructions. In brief, 50 μL of standard and activated milk samples were added to wells in duplicate, incubated for 2 hours at room temperature, and washed, and 100 μL of TGFβ1 or β2 conjugate was added. Following another wash, 100 μL of substrate solution was added for 30 minutes at room temperature, the reaction was ended with 100 μL of stop solution, and absorbance was measured at 450 nm. The detection limits of the TGFβ1 and β2 kits are 4.61 and 7.0 pg/mL, respectively.
Statistical analysis
The expression levels of proteins were first logarithm-transformed using natural base prior to analysis. The differences in protein expression at three time points (T, 2 months, and W) were tested with longitudinal analysis using a linear mixed model. The p values were adjusted for multiple tests with Holm's step-down procedure. For proteins that showed a significant change over the three time points, a paired t test was performed to detect the change between each pair of time points.
Results
Breastmilk samples were collected (Table 2) from 72 women: 35 parimiparas and 37 multiparas (second child for 25 women, third child for five, and fourth or more child for seven). Mean age was 28.3 years, and mean time from T to W milk collection was 278 days. One woman enrolled in the study after already breastfeeding for 2 months, and no T milk was available. A 2-month milk sample was not collected from two women because they weaned prior to the 2-month time point. Milk was collected at all three time points from 47 women. W milk samples were not collected from 25 women (14 had not yet weaned, nine had weaned but did not provide a W sample, and two were lost to follow-up). There was no association of individual protein expression with parity.
Total milk protein decreases during the course of lactation
W milk total protein concentration did not significantly vary based on time from T to W milk sample collection. The concentration of total protein was significantly lower (p<0.01) in the 2-month and W compared with the T milk samples (Fig. 1). Moreover, there was considerable variability in total milk protein concentration between samples. For these reasons, all individual protein results are listed, and all statistical analyses were conducted controlling for total protein concentration in the milk sample. Although mean protein concentration of W milk trended higher (1.34 vs. 1.25 ng/dL) than 2-month milk (Table 1), the difference was not significant. The mean total protein concentration of W milk did not significantly vary among women who nursed for <12 months versus those who nursed for 12 months or longer (1.34 vs. 1.35 g/dL, respectively).
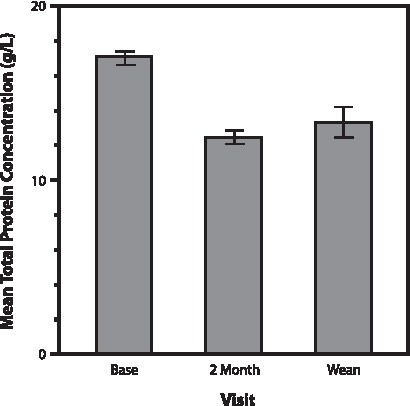
Total milk protein content (in mg/mL) at the beginning of lactation (transitional [baseline]), 2 months after starting, and at the end (wean). Bars represent SEM values.
Difference in protein expression based on timing of milk collection during the lactation period
We compared the protein expression of T and 2-month, T and W, and 2-month and W samples for each protein (Table 3). The expression of KLK5, -7, -11, and -14, YKL-40, TGFβ1, NGAL, and bFGF was significantly lower and that of TGFβ2 was marginally lower comparing T with 2 months. For 2 months versus W, expression was significantly higher in W samples for most of the same proteins (KLK5, -6, and -14, YKL-40, TGFβ1, TGFβ2, NGAL, and bFGF). Comparing T and W, KLK6 and TGFβ2 were significantly higher, whereas bFGF was significantly lower in W samples.
Data are in ng/g. Breastmilk collections were transitional (T) (within 10 days of the onset of lactation), 2 months after lactation started, and during weaning (W).
P value adjusted for multiple tests.
bFGF, basic fibroblast growth factor; KLK, kallikrein; NGAL, neutrophil gelatinase-associated lipocalin; N, sample size; NS, not significant; TGF, transforming growth factor.
The expression of some kallikreins is influenced by the length of time that a woman nurses
We evaluated the association of length of time nursing with individual protein expression in two ways: (1) expression at the W time point and (2) difference in expression in W and T milk for each woman for whom samples were collected and evaluated at both time points. Although length of nursing did not significantly influence W expression for any of the 16 proteins, it did influence the difference in KLK6, -7, and -8 expression between W and T time points. Specifically, the difference in expression of KLK6 (p=0.0012), KLK7 (p=0.020), and KLK8 (p=0.011) (Fig. 2) was related to length of time that a woman nursed.
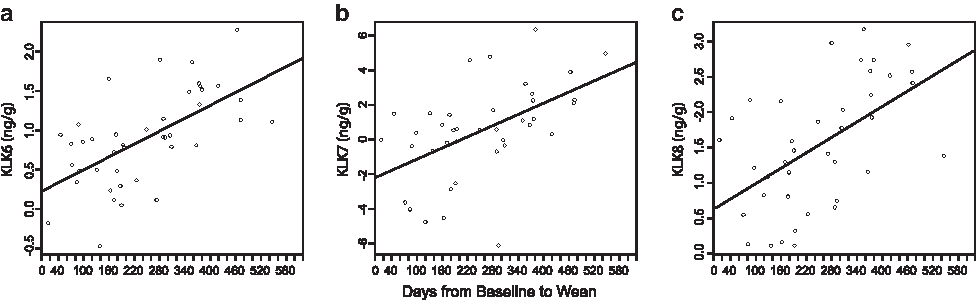
Change (wean – transitional [baseline]) in natural log of the kallikrein (KLK) proteins
Discussion
It is important to understand the pattern of total protein expression in breastmilk if one wants to assess the expression of individual proteins. Prior reports compared milk protein concentration between 3 and 12 months 5 and 3 and 14 months after the start of nursing, 6 with differing results. In our cohort, we did not observe a significant influence of length of nursing on total protein concentration. We therefore compared protein concentrations at three time points during lactation, regardless of the length of time that a woman nursed. We observed a significant decrease in total milk protein expression at 2 months and W compared with T. Although the reason(s) for this are uncertain, a possible explanation is the greater milk production during the 2-month and wean periods compared with T. Indeed, total milk protein concentration has been shown to negatively correlate with 24-hour milk volume. 28 The nonsignificant increase in milk protein concentration in our W compared with our 2-month milk samples is consistent with the samples being collected in the early-mid weaning period (Table 1).
We first attempted proteomic analysis of milk samples to obtain a global, unbiased assessment of differences in individual milk protein expression at three time points in lactation (T, mature, and W). Our results were disappointing, as only abundant proteins, which did not have a clear association with breast cancer or breast involution, were identified. We therefore performed a more focused approach using the observations in the involuting or involuted mammary glands of humans and animals.29,30 There is cross-reactivity using immunoassay between proteins in human and bovine milk, most notably bovine β-lactoglobulin. 31 On the other hand, many milk proteins have been successfully measured by immunoassay, including bFGF, 32 multiple KLKs, 12 and TGFβ. 33 All analyses involved immunoassays.
The worse outcome of pregnancy-associated breast cancer has been attributed to the pro-inflammatory, angiogenic environment of the breast present during involution, which is tumor promotional. 3 We investigated proteins previously associated with breast involution and/or breast cancer: inflammatory (YKL-40, NGAL16,30), angiogenesis (bFGF), 9 and TGFβ1 and -β2.29,30 In addition to these five proteins, we analyzed a panel of KLKs, many of which, as mentioned, are differentially expressed in hormone-related malignancies and are detectable in breastmilk. 12
The most common trend observed in the expression of individual milk proteins (after controlling for total protein concentration) was a lower concentration in the 2-month compared with the T samples and a higher concentration comparing 2-month and W samples. Because of this trend, as well as the importance of the time of weaning and involution on future cancer risk, our greatest attention was on the differences between T and W milk samples, in which KLK6 and TGFβ2 significantly increased, whereas bFGF significantly decreased.
The increase in KLK8 expression from T to W samples was greatest in women whose first full-term pregnancy occurred at a young age, which is consistent with the known protective effect of early age at first full-term pregnancy 34 and a tumor-suppressive effect of KLK. 8
KLK6 and -8 expressions are down-regulated in breast cancer. 35 KLK6 gene silencing has been reported in tumors. 36 As such, it has been speculated that it may function to suppress tumors, perhaps by inhibiting the epithelial-to-mesenchymal transition. 36 KLK8 has been shown to function as a serine protease. 37 The mechanism of KLK8 gene silencing in breast cancer has not been reported, although alternative splice variants of the gene that influence prognosis have been reported in lung cancer. 38 TGFβ proteins regulate apoptosis and matrix remodeling during breast involution induced by breast weaning, acting as tumor suppressors in the early stages of cancer. 39 TGFβ2 and its receptor are epigenetically silenced in early breast cancer by histone modifications. 40 On the other hand, we have found that bFGF expression is significantly increased in the breast nipple aspirate fluid of women with breast cancer.9,41 The expression of KLK7 and its association with breast cancer is more complicated. 42 The differences in breastmilk protein expression between T and W samples (increase in KLK6 and TGFβ2, decrease in bFGF) support a protective effect of breastfeeding, consistent with epidemiologic evidence. The greater increase in KLK6 and -8 expression (comparing W and T samples) with length of nursing is also consistent with a protective effect of a longer period of nursing. Whereas expression of KLK6 and -8 was consistently higher at the W than the T time point (with the exception of two women for KLK6), KLK7 W versus T differences were mixed for lactation periods of less than a year, but consistently higher for women who nursed for more than a year. Although this proposal is speculative, these proteins may act to counter the overall microenvironment of the involuting mammary gland, which has been proposed to promote mammary tumor progression. 4
This report has limitations. Most notable is the modest sample size. This may have prevented us from detecting more subtle differences in expression based on time of milk collection or parity. A second limitation is that there was some variability between when the W milk sample was collected and when the woman stopped nursing entirely. A third limitation is that we cannot say with certainty that the protein differences noted at the time of weaning would put these women at lower risk of future breast cancer because we do not have sufficient follow-up to address this.
Conclusions
We observed that total milk protein concentration was lower in 2-month and W milk compared with T milk samples. We document the up-regulation of KLK6 and TGFβ2 and down-regulation of bFGF at the time of breast weaning and involution. Finally, we observed that the length of time that a woman nursed influenced the degree of increased KLK6, -7, and -8 protein expression at the W versus T time points. These trends are consistent with a tumor-suppressive effect of breastfeeding. Further investigations of these proteins to validate our observations and to determine how the proteins work to prevent tumor formation in the lactating breast should be pursued.
Footnotes
Acknowledgments
This study was funded in part by the Avon Foundation for Women.
Disclosure Statement
No competing financial interests exist.