
Abstract
Abstract
The evolution of immunological agents in milk is intertwined with the general aspects of the evolution of the mammary gland. In that respect, mammalian precursors emerged from basal amniotes some 300 million years ago. In contrast to the predominant dinosaurs, proto-mammals possessed a glandular skin. A secondary palate in the roof of the mouth that directed airflow from the nostrils to the oropharynx and thus allowed mammals to ingest and breathe simultaneously first appeared in cynodonts 230 million years ago. This set the stage for mammalian newborns to nurse from the future mammary gland. Interplays between environmental and genetic changes shaped mammalian evolution including the mammary gland from dermal glands some 160 millions of years ago. It is likely that secretions from early mammary glands provided nutrients and immunological agents for the infant. Natural selection culminated in milks uniquely suited to nourish and protect infants of each species. In human milk, antimicrobial, anti-inflammatory, and immunoregulatory agents and living leukocytes are qualitatively or quantitatively different from those in other mammalian milks. Those in human milk compensate for developmental delays in the immunological system of the recipient infant. Consequently, the immune system in human milk provided by evolution is much of the basis for encouraging breastfeeding for human infants.
Emergence of Mammalian Precursors
And what was the Earth like in the Triassic? The principal landmass, the supercontinent Pangaea (from the Greek word for “entire earth”), was arid because it was concentrated along the Equator and except for its perimeter was far from oceans. Despite the harsh conditions, herbivorous dinosaurs survived because they adapted to a diet of the prevalent gymnosperms 4 and because of well-developed defenses against environmental microorganisms.5,6 Other clues to their survival are suggested from certain biological characteristics of their modern terrestrial relatives. Extant reptiles—lizards, snakes, and crocodilians—conserve water by excreting uric acid as a paste and by having thick, aglandular skins.
But before dinosaurs developed from basal dinosauromorphs7,8 300 to 310 mya, mammalian precursors—synapsids (from the Greek word for “fused arch”) (Fig. 1)—evolved from basal amniotes. 9 These terrestrial, four-footed vertebrates displayed amniontic, chorionic, and allantoic membranes during embryogenesis and laid eggs suited to a terrestrial existence. In addition, synapsids developed a glandular skin that in time led to the mammary gland.

Mammalian evolution. mya, millions of years ago.
In the next evolutionary step toward existing mammals, synapsids morphed into therapsids (from the Greek words for “wild animal” and “arch”) (Fig. 1) distinguished by powerful jaws, frontal incisors, large lateral canine teeth, and vertically oriented hind limbs. Then warm-blooded cynodonts (from the Greek word for “dog teeth”) 10 (Fig. 1) evolved from therapsid theriodonts (from the Greek word for “beast”) about 230 mya during the Late Permian–Early Triassic. One distinguishing feature of cynodonts was a secondary palate in the roof of the mouth 11 that directed airflow from the nostrils to the oropharynx. This allowed cynodonts to ingest and breathe at the same time. This set the stage for the time millions of years later when mammalian newborns nursed from the mammary gland.
Genetic Aspects of Mammalian Evolution
Interplays between the rigors of and opportunities in the environment and continual genetic changes (see sidebar) shaped mammalian evolution. Two examples of genetic changes may have led to different mammals and the mammary gland. (1) Genetic drift, the change in allele frequency in succeeding generations due to chance, is particularly important when selective environmental forces are weak. Moreover, genetic drift can cause populations with the same genetic structures to diverge into dissimilar populations. This was probably germane to the development of montherians, metatherians, and eutherians. (2) Genetic changes responsible for certain adaptations that originally served a particular function may coincidentally serve other purposes. As will be discussed subsequently, one exaptation (co-option) 12 probably played a major role in the dawn of the mammary gland. Some sequences of genetic changes and hence of mammalian evolution have been reconstructed from genomic studies of existing mammals. Examination of ancient fossils also helped to reconstruct mammalian evolution.
Single nucleotide polymorphisms
Addition of DNA sequences to ends of ancestral genes
Fusion of copies of preexisting DNA sequences rearranged by exon shuffling
Fusion of functional genes to form new multifunctional complexes
Gene duplications to generate families of proteins
Gene duplication and tandem fusion
Gene duplication followed by spontaneous, independent mutations
Utilization during translation of a hitherto unused or non-coding open reading frame
Environment of Early Mammals
To persist in a world dominated by dinosaurs, small, furry, quadruped nocturnal precursors of mammals found underground shelters to survive, reproduce, and raise their young. 13 In that respect, skeletons of the Triassic cynodont Thrinaxodon liorhinus 13 have been excavated from multichambered burrows. 14 Given their nocturnal activities and life in dark, earthen chambers during daylight, their hearing must have been good. However, the malleus and incus of the middle ear in modern mammals were not developed.
These adaptations protected adults and sheltered the offspring until they could fend for themselves. The underground environments were, however, problematic because there was little nutrition for the young except for roots, crawling insects, and annelids.
During the early Jurassic, rodent-like mammals 15 appeared in response to changing environments. Their “cheek-teeth” were separated from chisel-like front teeth by a wide gap. Each cheek-tooth displayed several rows of tubercles (hence the name, Multituberculate) (Fig. 1) that meshed with similar mandibular teeth. This apparatus efficiently masticated seeds and nuts. Their ecological niches ranged from burrows to trees. They persisted until the Oligocene, some 25 mya.
Multitubulates retained the temporal fenestra found in synapsids and formed the malleus and incus from bones in the jaw and quadrate, respectively. Those middle ear bones transmit sound-generated vibrations to the more ancient stapes that in turn transmits vibrations to the oval window of the cochlea of the inner ear. Because hearing was improved, those early mammals were better able to detect, avoid, and escape from predators and care for their young in dark environments.
Immunological Determinants of Survival of Early Mammals
Long before vertebrates appeared, the Earth was home to microorganisms, many of which were pathogens. For multicellular animals to survive, a wide spectrum of defense mechanisms evolved. The first evidence was found in 1883 by Ilya Ilich Metchnikoff (1845–1915), who observed that mobile cells engulfed splinters of rose thorns introduced into transparent, ciliated, free-living, bipinnaria starfish larvae.16,17 He called these mobile cells phagocytes (from the Greek, phagein and kytos, meaning devouring cells) and the process, phagocytosis. Metchnikoff further examined the defense by phagocytes by feeding Daphnia (small aquatic crustaceans) needle-like spores of the parasitic fungus Monospora bicuspidate. 18 Once the spores penetrated the digestive tract, mobile phagocytes surrounded them. The Daphnia died if the spores germinated, but lived if the phagocytes eliminated the ungerminated spores. Later, Metchnikoff demonstrated that similar mobile cells from mammalian blood (microphages, later called neutrophils) were induced during infections to pass through the walls of small blood vessels (diapidesis), migrate to the infecting agents (chemotaxis), engulf them (phagocytosis), and destroy them by intracellular digestion. 19
It was later found that many other specialized leukocytes and a host of soluble immunological agents, often structurally related to each other, evolved over many millions of years. 20 Some agents remained remarkably unchanged, whereas others further evolved to better resist pathogens. One prime example, the immunoglobulin superfamily, consists of over 700 different but genetically related proteins.21,22 They include cell membrane-bound or secreted antigen receptors (immunoglobulins and T-cell antigen receptors), antigen-presenting molecules (major histocompatibility molecules, Classes I and II), receptors for immunoglobulins on leukocytes and epithelial cells, receptors for cytokines, and immunomodulating molecules. Some members, such as antigen receptors, first evolved for systemic or mucosal immunity but were later adapted to operate in mammalian milks and thus protect the recipient infants.
Emergence of the Mammary Gland
The earliest therians (mammals capable of lactation
It is unclear when the mammary gland appeared, but the prototype may have surfaced during the early Cretaceous. It is suggested that secretions from dermal apocrine-like glands associated with hair follicles on the ventral thoraco-abdominal surface of mothers of some mammalian precursors provided nutrients and defense agents for the young to survive in dark, nutrition-poor, pathogen-laden environments. Nutrients from these secretions were central to the successful outcome of the offspring, but immunological agents may also have been important. In that respect, defense agents produced by human dermal glands are found in many mammalian milks. These include fatty acids in human sebum, 27 phylogenetically ancient lysozyme 28 produced by mammalian apocrine glands, 29 and melanotransferrin, an iron-binding protein found in human sweat glands, which has a 40% homology with lactoferrin, which is prominent in milk from many eutherian mammals. 30
Prototherians
Since the evolution of immune agents in mammalian milks was discussed in 1981, 31 much more has been learned about this subject from existing prototherians, the Monotremata (from the Greek words for “single” and “hole” which refers to the cloaca). However, that has to be placed in context with the general evolution of those creatures.
Fossils of the existing duck-billed platypus (Ornithorhynchus anaticus) date to the Paleocene, about 65.5–58 mya, when terrestrial dinosaurs disappeared. The other existing monotreme, the echidna (Tachyglossus aculealus), diverged from the platypus lineage 19–48 mya when they transited from a semi-aquatic to a terrestrial life. 23
Some features of monotremes are unlike those in metatherians and eutherians and harken back to mammalian precursors. They include (1) microchromosomes, (2) filiform sperm, (3) epipubic bones found otherwise only in fossils of therapsids, (4) egg yolk genes 32 that result in the formation of leathery, uncalcified eggs, and (5) venom production. 33 Monotremes are also unlike other therians in that they have 10 rather than two X chromosomes in females and five X- and five Y-chromosomes in males. 34 Some genetic markers of the platypus are found in other mammals, and some are not. 35 Perhaps by convergent evolution, the gene for the venom produced by the crural gland located near the male's ankle spur is shared with reptiles. In contrast, genes for many specific milk proteins found in monotremes are shared with other mammals. 35
An echidna female lays a single egg and deposits it into her ventral pouch. After hatching, the young echidna remains in the pouch for 45–55 days. The mother then digs a nursery burrow and deposits the young, returning every 5 days to suckle it until it is weaned at 7 months. The need of a burrow may recapitulate the behavior of ancient mammalian precursors.
In keeping with the previously stated lactation hypothesis, female monotremes do not have nipples. Richard Owen illustrated in 1832 36 (Fig. 2) that milk produced by the platypus (O. anatinus) flowed from a cluster of openings of mammary gland ducts located in a patch of the ventral abdominal surface. Because birthing precipitates the flow of those secretions, hormones may be involved in the process.
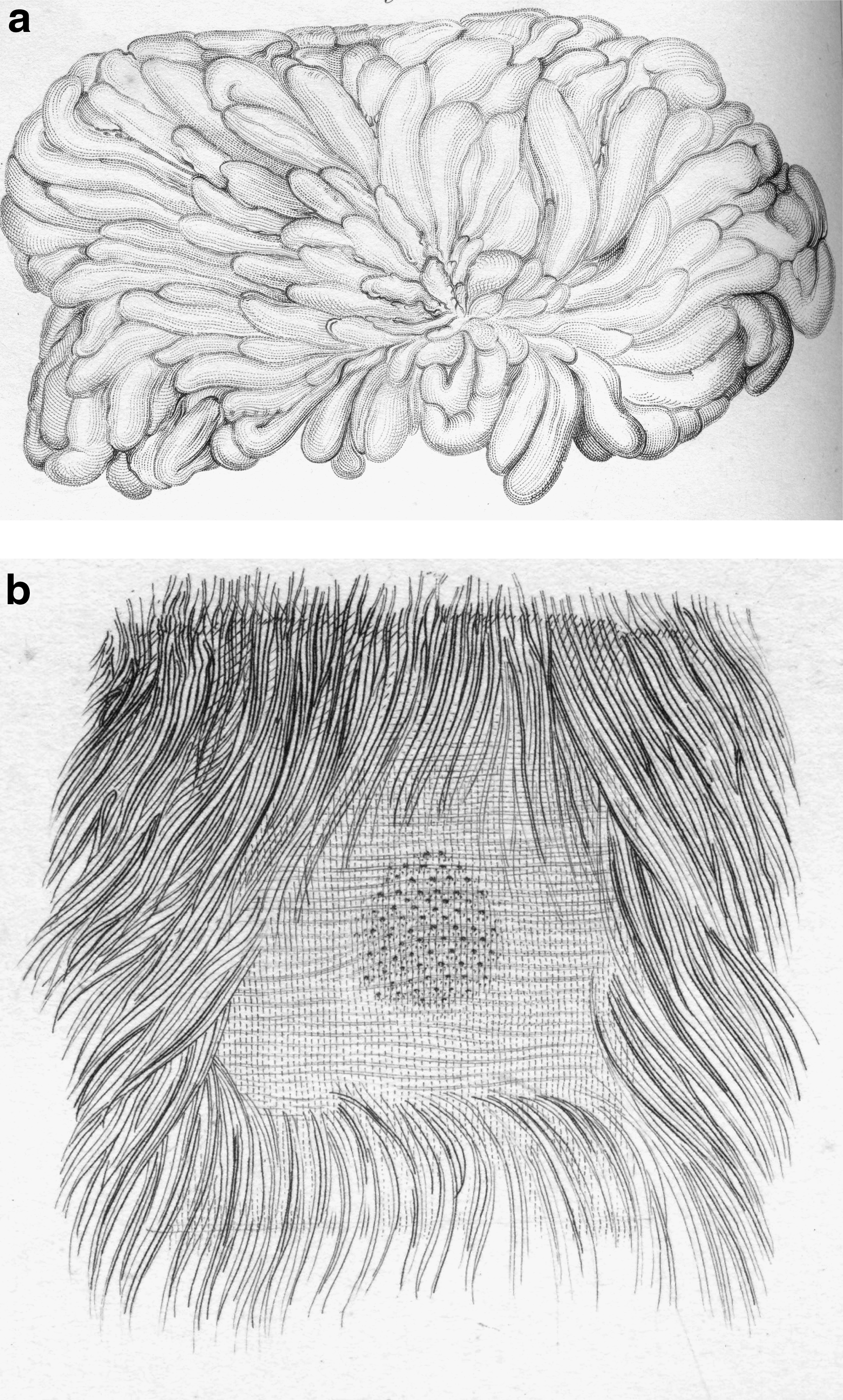
The figure published by Richard Owen
36
in 1832 showed that milk produced by a platypus (O. anatinus) mammary gland
Metatherians
Metatherians probably appeared in the Cretaceous.37,38 They developed a yolk sac type of placenta to support the establishment of a fertilized ovum and its development into a fetus. In contrast to eutherians, the gestational period in marsupials is much shorter than the duration of lactation. 39 In that respect, the newborn metatherian is atricial and survives by crawling to the mother's ventral pouch and attaching to a teat to feed from mammary gland secretions. Lactation in these animals is complex. The composition of kangaroo milk changes as lactation proceeds. 40 For instance, the tammar wallaby (Macropodidae eugenii) has an asynchronous, concurrent lactation. Adjacent glands feed two offspring of different ages with milk of different nutrient compositions. 40
Milk from modern metatherians has many immunological agents in common with eutherian milks. These include polymeric immunoglobulins, transferrin, lysozyme, α-lactalbumin, complement factors, CCL28, cathelicidin, and leukocytes.40–46 Furthermore, concentrations of many of those agents change as lactation proceeds.40–46 In addition, the immunological functions of the metatherian mammary gland are crucial to neonatal survival because marsupial young do not develop adult immune responses until just before detaching from a teat and leaving the pouch.
Eutherians
In contrast to other mammals, eutherians have a complete placenta characterized by a definitive interface or barrier, an orderly fetomaternal interdigitation, a distinct shape, and an organized fetomaternal blood flow.47,48
Comparative genomic analyses and evolutionary simulations reveal that the three ancestral vitellogenin-encoding genes in reptiles were progressively lost during mammalian evolution about 30–70 mya in all but egg-laying monotremes. 49 In contrast, caseins, which appeared in a common mammalian ancestor approximately 200–310 mya, have functions similar to vitellogenins and have persisted in all mammalian milks.
Molecular dating suggests that the earliest eutherians appeared some 145 mya,37,50 but eutherians did not diversify and radiate until the great Cretaceous extinction 65 mya50–52 (Fig. 3). Bayesian analysis of molecular genetic evidence from existing species suggests that the primary branching that gave rise to superordinal clades of eutherians Afrotheria (aardvarks, elephants, elephant shrews, hyraxes, manatees, tenrecs), Xenartha (anteaters, armadillos, sloths), Euarchontoglires (rabbits, rodents, primates), and Laurasiatheria (carnivores, cows, horses, whales) coincided with the breakup of the giant southern continent Gondwana into Africa and South America. 52 Most eutherians became extinct.

Evolution of eutherian species including primates and H. sapiens.
Our lineage began when primates appeared 65 mya (Fig. 3). The earliest hominids (the genus Australopithecus) diverged from other primates 5–6 mya. 53 Later hominids arose 2.5 mya.54,55 Homo sapiens emerged 100,000–250,000 years ago56–59 (Fig. 3).
The evolutionary proximity of existing mammals has been reconstructed from phenotypic, genotypic, and cladistic analyses of current mammals. Genomes of humans and chimpanzees (Pan troglodytes and Pan paniscus) are remarkably similar,59,60 whereas far greater differences in genomic and phenotypic features are found between primates and other mammalian orders. 60
Evolutionary relationships between the immunological activities of the mammary gland have been investigated. Some defense agents in mammalian milks are remarkably conserved. Two examples are the antigenic similarities between the carboxy-terminal, intracytoplasmic regions of the MUC 1 mucins found in milks from many mammalian species 61 and structural similarities in secretory antibodies, 62 lysozymes, 28 and α-lactalbumin 28 in mammalian milks. In contrast, some immunological functions of the mammary gland were not as highly conserved. Because of exposures to different microorganisms, it seems that certain immune responses diverged for the young of various orders of mammals to survive. Furthermore, because of major distinctions in the rate of appearance and degree of motor development, the type and extent of exposure to microorganisms by infants of those species vary considerably. It is plausible that immunological adaptations to these different environments are also reflected in the different immunological composition of milk in diverse species.
In general, the greater the evolutionary distance, the more dissimilar is the immunological composition of milks from those species. For example, differences between human and cow milk, including the types and quantities of antimicrobial oligosaccharides63–65 and the concentrations of immunoglobulin isotypes,66–70 lactoferrin,66–71 lysozyme,66–70,72 lactoperoxidase,73,74 and milk mucins, 75 are striking.
For example, certain immunoglobulin isotypes that are rare or absent in human milk are prominent in rat and cow milk. The concentration of immunoglobulin D (IgD) is approximately 100-fold greater in rat milk than serum. 76 IgD in rodent milk is not absorbed into the systemic circulation of the recipient infant. IgD is a primordial immunoglobulin expressed in most jawed vertebrates, but other than its role as a transmembrane antigen receptor on B cells 77 and some role in inflammation by binding to myeloid cells, 78 its functions are unknown. The biological effect of ingested IgD in the developing rat pup is unknown.
The second example is immunoglobulin E (IgE), which is prominent in bovine colostrum and which is transferred to the calf's systemic circulation. 79 In most mammals IgE is antiparasitic. In mice, IgE induces the production of mast cells from murine bone marrow precursors and increases their ability to produce certain cytokines. 80 The systemic effect of IgE in bovine milk in bovine calves is unknown.
Requirements for the Evolution of the Mammary Gland
As previously hypothesized, the mammary gland most likely evolved from dermal apocrine glands (Fig. 4). The ventral part of the thoraco-abdominal area was the logical place for the mammary gland to develop because that was where eggs would have been incubated and where newborns would have hatched. The molecular genetic events that led to this development are not entirely established, although co-option seems plausible.

Approximate steps in evolution of the mammary gland and its immune system.
A discussion of this possibility began when the controversial British biologist St. George Jackson Mivart 81 commented 82 upon Charles Darwin's first book on the origin of species. 83 Mivart stated “that natural selection is incompetent to account for the incipient stages of useful structures.” With respect to the mammary gland, he observed, “Is it conceivable that the young of any animal was ever saved from destruction by accidentally sucking a drop of scarcely nutritious fluid from an accidentally hypertrophied cutaneous gland of its mother?”
Darwin (Fig. 5) replied in the sixth edition of his famous book 84 :

Charles Darwin, the originator of the concept of evolution due to natural selection, was the first to hypothesize that the mammary gland originated from dermal glands. The photograph was taken in 1854 when Darwin was age 45. At that time, he was preparing his famous book The Origin of Species by Means of Natural Selection or the Preservation of Favoured Races in the Struggle for Life, which was published in 1859. 83 Reproduced with permission from the Truman G. Blocker History of Medicine Collections and the Moody Medical Library at the University of Texas Medical Branch.
… is it not at least possible that the young might have been similarly nourished? And in this case, the individuals which secreted a fluid, in some degree or manner the most nutritious, so as to partake of the nature of milk, would in the long run have reared a larger number of well-nourished offspring, than would the individuals which secreted a poorer fluid; and thus the cutaneous glands, which are the homologues of the mammary glands, would have been improved or rendered more effective. It accords with the widely extended principle of specialization, that the glands over a certain space of the sack should have become more highly developed than the remainder; and they would then have formed a breast, but at first without a nipple. …
Darwin further hypothesized that mammary glands evolved from cutaneous glands in brood pouches of seahorses (Hippocampus). The general idea was correct, but the origin of the mammary gland was not in fish but in terrestrial vertebrates. Nearly a century after Darwin's hypothesis, the population geneticist John Haldane suggested that lactation arose from frequent sucking of water droplets from hypertrophied cutaneous glands and commented upon the potential genetic aspects of the development of lactation. 85
The most compelling reconstruction of morphological aspects of mammary gland evolution and its relationship to the newborn infant was by Olaf Oftedal in 2002.86,87 He hypothesized that secretions from ventral dermal glands associated with hair follicles of early mammals could have moisturized the mother's eggs. 87 After hatching, the young would have therefore quickly encountered the mother for affection, warmth, and secretions from those ventral dermal glands. Oftedal further commented that advanced Triassic therapsids, such as cynodonts, almost certainly secreted complex, nutrient-rich milk, allowing a progressive decline in egg size and an increasingly altricial state of the young at hatching. Synapsid eggs did not have calcified eggshells. 87 But, it is unclear whether eggshells from all other early mammals were parchment-like and therefore required additional water after being laid or were more impermeable such as found in avians. 88 It is also possible that such eggs were buried to prevent a loss of water. Further studies may clarify those possibilities.
Mammary gland development very likely also depended upon the acquisition of oxytocin receptors on dermal secretory glands. The precursor to that event was the production of oxytocin (Fig. 4). In that respect, all vertebrates produce an oxytocin-like nonapeptide. 89 The synthesis of oxytocin is marked in the magnocellular neurosecretory cells of the supraoptic and paraventricular nuclei of the hypothalamus. Oxytocin is then transported to the axon terminals of the posterior pituitary. There, oxytocin is bound to neurophysin. 90 Secretion of oxytocin from neurosecretory nerve endings is regulated by electrical activity of hypothalamic oxytocin cells that generate action potentials that propagate down axons to nerve endings in the pituitary. 91 Oxytocin-containing vesicles in those nerve endings are released when nerve terminals are depolarized. If oxytocin receptors were on certain contractile cells in dermal secretory glands of early mammals, then stimulation of afferent nerves in those areas by suckling would have led to an electrical stimulation of spinal nerves that connect with the hypothalamus and hence cause the release of oxytocin from the pituitary. The consequent binding of oxytocin to receptors on those dermal glands would have caused them to secrete more vigorously and thus augmented the amount of secretions to the suckling newborn.
Because prolactin plays an important role in lactation, it is of interest that prolactin receptors have been demonstrated on human hair follicles. 92 This suggests that prolactin receptors were present in the primordial mammary gland that evolved from similar epidermal structures (Fig. 4).
Releaser-type pheromones may also helped to establish lactation and nursing. In that respect, pheromones released in rabbit milk trigger immediate nursing behavior in their kits. 93 The potential role of releaser-type pheromones in early lactating mammals is also in keeping with the notion that olfaction was well developed in these furtive creatures.
Development of Eutherian Mammary Gland
Certain evolutionary events were essential for the development of lactation in eutherian (placental) mammals. Placental mammals appeared in the Ecocene (approximately 55 mya), and their complexity increased in response to less competition from other species and more available environmental niches. At the same time the eutherian mammary gland assumed its present form and function by developing (1) branching arbor-like structures94,95 (Fig. 4), (2) hormones such as progesterone, 96 estrogen, 96 prolactin, 97 and growth hormone 97 that stimulate the development, maturation, and function of the mammary gland during the transition from childhood, adulthood, pregnancy and lactation, and (3) secretory mechanisms that transfer soluble or compartmentalized bioactive agents and whole cells into milk.
In parallel was the development of the true placenta, which produces placental lactogen and estrogen that promotes the growth of the mammary gland, supplies oxygen, nutrients, and immunoglobulin G (IgG) to the fetus of certain eutherian species and eliminates waste products from the fetus.
As the complexity of the mammary gland evolved, mammalian mothers required more energy, nutrients, and metabolic processes. Indeed, the initiation and continuation of lactation are expensive biological processes.98,99 This suggests for the mammary gland to evolve into its present form, adult mammals would have required increased sources of calories, proteins, and other essential nutrients.
Indeed, the complexity and uniqueness of milk from each mammalian species attest to its importance. In that respect, milk is the most complicated external secretion in every eutherian species. It is therefore no surprise that formidable amounts of energy, nutrients, enzymes, and other cellular machinery are needed to initiate and maintain lactation.
The formation of secretory immunoglobulin A (IgA) antibodies in human milk is a prime example. The process begins when immunoglobulin M (IgM)-positive B cells in Peyer's patches in the ileum are stimulated by ingested foreign antigens with the help of certain cytokines produced by local helper T cells to switch isotypes from IgM to IgA. Then these IgA-positive B cells sequentially migrate to regional lymphatics, the cisterni chyli, the thoracic duct, the peripheral blood, and the lamina propria of the mammary gland. The homing to the mammary gland only occurs during lactation. In the mammary gland, IgA-positive B cells transform into plasma cells that produce and secrete dimeric IgA that binds to polymeric immunoglobulin receptors on basolateral membranes of mammary gland epithelium. The ligand–receptor complexes are interiorized, the intracytoplasmic part of the receptor is cleaved off, and the remaining assembled molecule, secretory IgA, is secreted into milk. The antigen-binding specificity of those antibodies matches that of the enteric antigens that set the process in motion in Peyer's patches. Each step requires the activation of metabolic processes and the expenditure of considerable energy.
The advent of and subsequent changes in the mammary gland were surely driven by natural selection to provide survival advantages to the newborn of the species. If the young were better nourished, more quickly developed to contend with extrauterine life, and better protected from microbial pathogens that pervaded the maternal environment, then those infants not only survived, but also developed into more able reproductive adults.
The evolutionary adaptations of the composition of mammalian milks are remarkable. For instance, seals, which leave their young for considerable periods to search for food, 100 produce milk that has a much higher fat content that provides more energy than carbohydrates or proteins and much less lactose than mammals who feed their young more frequently. 101
Evolutionary Advantages of Lactation
Recently it has become more apparent that lactation provides numerous evolutionary advantages for the suckling young. In that respect, a host of growth and maturation factors and an exceptionally complex immunological system are provided for the recipient to compensate for developmental delays in the recipient's immunological system. The latter permits the infant to direct more energy and nutrients to the growth and development of other vital body parts such as the brain, lungs, endocrine organs, muscles, and skeleton. In that respect, the immune system is metabolically expensive to develop, maintain, and replenish. Because of the antimicrobial, anti-inflammatory, and immunoregulatory properties of human milk, during infections and other inflammatory processes, the integrity of the gastrointestinal tract and the respiratory system is preserved to insure normal nutrition and normal gas exchange, respectively.
Furthermore, the immune system in human milk evolved to aid the switch from marked intestinal permeability, a physiological state suited for fetal life, to a much less permeable state that protects against enteric pathogens. That system also permits infants to be actively immunized to environmental pathogens without developing signs of infection or inflammation. Such a sheltered immunization was the natural precursor to live attenuated microbial immunizations.
An insight into the tremendous evolutionary advantages of lactation is revealed by the interplay between human milk and the developing infant. First, human milk supplies the optimal requirements for total energy, amino acids, proteins, carbohydrates, lipids, and micronutrients for the infant. The infants digest and absorb these nutrients because of their innate abilities and the aid of bioactive agents in the milk. In that respect, the growth of parts of the absorptive and digestive system of the infants is also often assisted by differentiation and growth factors in the milk.
The many immunological advantages accrued from the evolution of the mammary gland are very important.102–104 One example recognized soon after the discoveries of immunoglobulins in milk was that ruminants such as bovine and ovine species, whose newborn offspring are pan-hypogammaglobulinemic, produce colostrum that has a high concentration of IgG. 105 Furthermore, that maternal IgG is transported from the newborn's intestines into the systemic circulation where it protects from many bacterial infections. 105 This is in sharp contrast to humans, in whom IgG is actively transported via the placenta to the fetus 106 but is in short supply in human milk. 107 In that respect, the dominant immunoglobulin in milk is the non-inflammatory, digestive-resistant secretory IgA that is adapted to protect mucosal surfaces from infections.
The positive immunological effects of human milk upon the recipient infant102–104,107,108 that have evolved can be summarized as follows:
1. Certain postnatal developmental delays in the infant's immune system are compensated for by the transmission of the same agents in human milk. 2. Agents in human milk other than those that are developmentally delayed offset certain postnatal delays in the immune system in the infant. 3. Some agents in human milk initiate or augment functions poorly expressed in the infant. 4. Many of the antimicrobial agents in human milk act synergistically with each other. 5. Some agents in human milk alter the physiological state of the alimentary tract from one suited for fetal life to one that is appropriate for extrauterine life. 6. Cells that produce antibodies in human milk originate in the maternal intestines and bronchi. 7. Defense agents in human milk protect against microbial pathogens without provoking inflammation in the infant. 8. Some agents in human milk inhibit inflammation. 9. Some agents in human milk display antineoplastic properties. 10. Defense agents in human milk resist enzymatic digestion and thus function in the recipient's gastrointestinal tract. 11. Certain defense agents are created by partial digestion of substrates in milk in the infant's gastrointestinal tract. 12. When defense agents in human milk interact with some pathogens, the infant develops specific adaptive immune responses but no symptomatic infections. Such a sheltered immunization is similar to immunizing with an attenuated microbial pathogen. 13. A host of immunomodulating agents in human milk appear to lessen the risk to certain immune-induced diseases long after weaning. 14. Agents in human milk augment the growth of commensal enteric bacteria adapted to infants that produce compounds that protect against bacterial pathogens and convey other immunological benefits. 15. There is often a reciprocal relationship between the defense agents that are transmitted in milk and those transmitted during fetal life via the placenta.
It is also remarkable that some agents in human milk are in macromolecular compartments such as milk fat globules or casein micelles or are bound to other proteins such as specific soluble receptors.107,108 These compartments and receptors may enhance their survival or direct the site or type of their actions. Also, human milk contains significant numbers of neutrophils, macrophages, and T cells, all of which are activated.107,108 Furthermore, the quantities of many of these agents and leukocytes in milk change as lactation progresses and the infant's immune system matures.102–104,107,108
All of these processes resulted from millions of years of experiments in nature that are difficult to reconstruct. However, biochemical analyses of milk from many mammalian species reveal that evolutionary adaptations depended upon the environment of the species in question. As in other aspects of biological evolution, interactions between random genetic mutations and environmental circumstances were decisive. Although many basic features of mammalian milks, as shown by biochemical and genomic analyses, 109 were retained during evolution, 109 the compositions of mammalian milks were found to be most similar in closely related species.
The study of the evolution of the mammary gland reveals molecular adaptations that are peculiar to the mammary gland such as the expression of α-lactalbumin, 110 whereas others are shared in other sites. Two examples of the latter are lysozyme 111 and lactoferrin,112–114 which are found in many tissues, cells, and secretions. Furthermore, many bioactive agents in milk including almost all of its immunological agents are found in systemic or mucosal parts of the immune system.
The evolution occurred in a very parsimonious manner from proto-mammals to extant mammals. In that respect, monotreme milk contains several components that are similar to those found in milk from certain eutherian species.115–117 They include difucosyl-lactose 115 (which interferes with the attachment of Campylobacter jejuni or Escherichia coli stable toxin to epithelial cells), lysozyme, 116 and transferrin, 117 which is structurally and functionally similar to lactoferrin in eutherian milks.
Medical Significance of Mammary Gland Evolution
Many healthcare professionals have not considered evolution as a determinant of infant nutrition. Thus, many may equate human milk with milks from many other mammalian species. Although milks produced by all mammalian species are somewhat similar, milk from each mammalian species is unique. Moreover, milk from each species changes quantitatively and sometimes qualitatively according to the duration of lactation. Furthermore, immunological functions of the mammary gland in each mammalian species evolved to compensate for particular developmental delays in the immune system of the recipient infants of each species.
The robustness of lactation and of the immune system in milk of each mammalian species is also attested by the necessity for many species to adapt to changing environmental circumstances and to the stream of new pathogens that develop because of spontaneous mutations. Mammalian milks have thus greatly helped newborns and young infants of each species to survive long enough to gain sufficient independence and a sufficient immune system for defense against the pathogens that pervade their habitat.
Our species was not exempt from these evolutionary dictates. As humans migrated out of African many tens of thousands of years ago,118,119 very different and at times challenging climes were encountered that were marked by different microbial flora, vegetation, and animal species. 120 But, lactation was preserved even when nutrients were limited. Moreover, humans modified their environments by creating agricultural communities, industrial societies, and better sanitation. However, those measures did not replace the unique advantages of breastfeeding. Indeed, the advent of agricultural and then industrial communities posed new problems such as the increased density of microbial pathogens within closed communities. Thus, the immune system in human milk was needed for those new challenges.
The dynamic features of lactation and the specificity of the composition of milk produced by each species indicate that it will be exceeding difficult to artificially replicate the milk of any species at any given point of lactation. Although progress has been made to “humanize” certain milk formulas for human infants, it is unlikely that such formulas will have the health impact of human milk. Furthermore, it is unclear whether desirable effects occur when an isolated defense agent from human milk is added to an artificial milk formula because of the interplay between many agents in human milk. In addition, donor milk fed to an unrelated infant may not have the full biologic advantages as milk from one's own mother because of the synchronizations between the production of immunological agents in human milk and the timing of the development of the infant's immune system. Finally, banked human milk, although superior to artificial feedings, is dissimilar to freshly secreted milk because of changes that occur during collection, processing, and storage.67,121,122
These issues should be viewed through the lens of biological evolution that began eons ago and culminated in the complex structure and many beneficial functions of the mammary gland. Indeed, the specific advantages of milk of each species, including macronutrients, micronutrients, growth factors, hormones, and immunological agents, for its particular progeny are an outcome of myriads of experiments in nature. Unless there are health preclusions to the mother or child, 123 breastfeeding remains by far the best choice for feeding human infants.
Footnotes
Acknowledgments
The late Dr. Otakar Koldovský from the University of Arizona Heath Science Center encouraged this work. The illustrations by Richard Owen were provided by The Truman G. Blocker History of Medicine Collections at the University of Texas Medical Branch in Galveston. Finally, Pfizer Nutrition supported this work.
Disclosure Statement
No competing financial interests exist.