
Abstract
Abstract
Background:
Human milk provides a multitude of glycoproteins, including highly glycosylated α-1-acid glycoprotein (AGP), which elicits anti-inflammatory and immunomodulatory properties. The milk AGP glycoforms may provide the breastfed infant with a wide range of biological benefits. Here, we analyzed the reactivity of O-linked sugar-specific lectins with human milk AGP over the process of lactation and compared the results with those of the lactating mother's plasma.
Materials and Methods:
Relative amounts of human skim milk AGP O-glycans were analyzed in early colostrum, colostrum, and transitional and mature milk samples of 127 healthy mothers by lectin–AGP enzyme-linked immunosorbent assay using sialyl T (sialyl-α2,3/α2,6 Galβ1,3GalNAc-), asialyl T (Galβ1,3GalNAc-), and Tn (GalNAc-) antigen-specific biotinylated Artocarpus integrifolia (Jacalin), Arachis hypogaea (PNA), and Vicia villosa (VVA) lectins, respectively.
Results:
Milk AGP elicited high expression of Jacalin- and PNA-reactive glycotopes and low expression of VVA-reactive glycotopes, which were absent on plasma AGP of lactating mothers and healthy individuals. The expression of sialyl, asialyl T, and Tn glycotopes of human milk AGP was lactation stage related. The relative amount of Jacalin-reactive AGP glycotope was highest in the colostrum samples and then decreased starting from Day 8 of lactation. In contrast, an increase of the relative amount of PNA-reactive glycotope with milk maturation was observed. The relative amount of VVA-reactive glycotope remained almost constant over the development of lactation.
Conclusions:
Milk AGP differs from mother's plasma AGP by the presence of O-linked sialylated and asialylated T as well as Tn antigens. The variation of the expression of sialylated and asialylated T and Tn antigens on AGP is associated with milk maturation.
Introduction
H
So far the free HMOs, 1 human milk N-glycome, 5 and glycan profiles of N- and/or O-glycans of some glycoproteins, 6 such as lactoferrin, 7 bile salt–stimulated lipase, 8 secretory immunoglobulin A,9,10 α-1-acid glycoprotein (AGP), 11 and fibronectin, 12 have been characterized in detail. Over the process of lactation, within milk maturation, the degree and type of sialylation and fucosylation of milk glycoproteins change and partly overlap with observed trends for HMOs abundantly present in milk.6,11,12 Gao et al. 13 have suggested that alteration in glycan structures during lactation goes hand in hand with a transformation in defense mechanisms that occur from newborns to young infants.
Human milk O-glycome has received less attention than N-glycome but appears to play an equally important role because O-glycans are also known to be involved in cell communication and adhesion, receptor–ligand interactions, and host–pathogen interactions, as well as protection of proteins from digestion.14–16 Moreover, O-glycosylation sites as well as O-glycan structures are tissue specific 17 and differ significantly during inflammation 18 and in cancer.19,20 Additionally, a wide range of O-glycans of colostrum secretory immunoglobulin A can interact with bacterial adhesins, resulting in blockage of their adhesion to host tissues, and they are considered to be a part of innate immunity. 9
AGP is an acute-phase plasma glycoprotein, produced mainly by the liver, but it can also be synthesized locally by human mammary epithelial cells, 21 alveolar macrophages, endothelial cells, monocytes, and leukocytes.22,23 In normal serum the concentration of AGP is 0.5–1.0 g/L, and during normal lactation, the AGP level in plasma of lactating mothers is at the same level as for healthy, nonlactating women, whereas its concentration in human skim milk is about 1/30th of that in plasma and ranges from 6 to 51 mg/L between Days 2 and 45. Additionally, its concentration decreased with milk maturation and correlates with the total protein concentration decline. 11
AGP glycosylation is tissue specific, is microheterogeneous in the degree of branching, fucosylation, and sialylation, and can change in relation to the physiopathological state of the human organism.11,23–25 Local-specific glycoforms of AGP have been reported for amniotic fluid 26 and human skim milk AGP, 11 and it was manifested by the increase of fucosylation as well as sialylation levels in comparison with those of normal as well as lactating mothers' plasma AGP. Moreover, the degree of sialylation and fucosylation of milk AGP was strongly correlated with milk maturation. 11
The impact of the N-glycan part of plasma AGP on its function has been well documented.23,24 The highly sialylated and fucosylated N-glycans of AGP are known to possess anti-inflammatory properties and to participate in the modulation of leukocyte–endothelial interactions by binding to E- or P-selectin. Additionally, AGP is known to take part, via competitive binding of its sialylated glycans, in inhibition of adhesion of some microorganisms causing disease, namely, Plasmodium falciparum and Mycoplasma pneumoniae, to the host's cells.23,24 In a bovine model, milk AGP, as an immunoregulatory molecule, was shown to induce the expression of some pro- and anti-inflammatory cytokines by monocytes during inflammation of the mammary gland. 27
The human plasma AGP is capped by the five N-linked glycans exclusively, but a fetal AGP, in addition to N-glycans, contained three O-linked glycans. 24 Here, we have documented that human milk AGP is also decorated by O-linked glycans whose expression changes over the course of lactation. Because the analysis of O-glycans is still challenging, especially due to a lack of a convenient method of O-glycan release from a core protein, 28 we have used an alternative, low-cost- and time-consuming, lectin-based enzyme-linked immunosorbent assay (ELISA). AGP extracted from biological samples directly on the ELISA plates coated with desialylated and defucosylated anti-AGP antibodies 11 was analyzed with Artocarpus integrifolia (Jacalin), Arachis hypogaea (PNA), and Vicia villosa (VVA) lectins specific to sialyl T (α2,3- and/or α2,6-sialyl-Galβ1,3GalNAc-), asialyl T (Galβ1,3GalNAc-), and Tn (GalNAc-) antigens O-linked to the protein part. 29 The degree of lectin reactivities with sialyl T, asialyl T, and Tn antigens was determined on milk and lactating mothers' plasma AGP over the course of normal lactation from Day 2 to Day 45.
Materials and Methods
Participants
Human milk (n=127) and plasma (n=25) samples were collected from healthy lactating mothers receiving regular perinatal care at the 1st and 2nd Departments of Gynecology and Obstetrics at Wrocław Medical University, Wrocław, Poland. Lactating mothers (from 19 to 36 years old) who were in a good state of health and had an uncomplicated and single normal pregnancy were recruited for the study, and informed consent was obtained from all the donor mothers. Mothers who used tobacco products, illicit alcohol, and drugs, as well as mothers with inflammatory diseases (e.g., mastitis or infection), were excluded from the study. The study protocol was approved by the Ethics Committee at Wrocław Medical University (protocol number KB-30/2013).
Sample collection and preparation
Human milk samples were collected at the end of nursing (hindmilk), once per day at the same time (8:00–10:00 a.m.) by a trained nurse by complete breast emptying. There was significant interindividual variation in volume of hindmilk, from 5 mL for colostrum to 100 mL for mature milk. To protect the integrity of samples, immediately after collection (up to 30 minutes), all milk samples were frozen in sterile plastic containers and stored at −20°C until analysis. Skim milk (aqueous phase) was prepared by centrifugation at 3,500 g at +4°C for 30 minutes, after which cells and the fat layer were removed. All samples were assayed by the same person.
Milk samples were divided into the following groups: (1) early colostrum from Day 2 to Day 3 of lactation, n=19 (AGP concentration, 23±11 mg/L; Maackia amurensis lectin (MAA) reactivity, 0.4±0.2 absorbance units [AU]; Sambucus nigra lectin [SNA] reactivity, 1.2±0.2 AU 11 ); (2) colostrum from Day 4 to Day 7 of lactation, n=53 (AGP concentration, 18±5 mg/L; MAA reactivity, 0.5±0.2 AU; SNA reactivity, 1.2±0.3 AU 11 ); (3) transitional milk from Day 8 to Day 14 of lactation, n=37 (AGP concentration, 18±7 mg/L; MAA reactivity, 0.5±0.2 AU; SNA reactivity, 1±0.3 AU 11 ); and (4) mature milk from Day 15 to Day 45 of lactation, n=18 (AGP concentration, 14±5 mg/L; MAA reactivity, 0.4±0.2 AU; SNA reactivity, 0.6±0.3 AU 11 ).
Additionally, blood samples from lactating mothers on postpartum Day 2 (n=25) and healthy volunteers (n=25) were collected and centrifuged at 2,000 g for 15 minutes, and the plasma obtained was divided into aliquots and stored at −76°C until use.
Lectin-based ELISA of AGP glycotopes
Expression of T and Tn antigens on a constant amount of 100 ng of AGP, estimated previously, 11 was determined by lectin–AGP ELISA according to a slightly modified procedure described previously11,25 using specific biotinylated lectins (Vector Laboratories, Inc., Burlingame, CA) with well-described binding preferences described by Wu et al. 29 PNA, Jacalin, and VVA are known to have binding preferences to asialylated T antigen (asialyl T) (Galβ1,3GalNAc), sialylated T antigen (sialyl T) (α2,3/α2,6 sialyl-Galβ1,3GalNAc), and Tn antigen (GalNAc), respectively.
The experimental details of lectin–AGP ELISA were as follows: goat anti-human AGP antibodies, oxidized with sodium periodate and desialylated with Vibrio cholerae neuraminidase,11,25 diluted (2,000 times) in 10 mM Tris-buffered saline (pH 8.5), were coupled to a polystyrene microtiter ELISA plate and used to extract AGP from the milk and plasma samples. For the test, 100-μL samples of milk or plasma, which were prediluted in 10 mM Tris-buffered saline and 0.05% Tween 20 (pH 7.4) to a final AGP concentration of 1 mg/L, were taken.
The presence of sialyl T, asialyl T, and Tn glycotopes on AGP was detected by a reaction with specific biotinylated lectins: Jacalin, PNA, and VVA. The AGP–lectin complex was quantified with phosphatase-labeled ExtrAvidin® (Sigma, St. Louis, MO) and then detected by the reaction with disodium 4-nitrophenyl phosphate (Merck, Darmstadt, Germany). The absorbance was measured in a Stat Fax® 2100 microplate reader (Awareness Technology Inc., Palm City, FL) at 405 nm with the reference filter at 630 nm. All samples were analyzed in duplicate. Controls were carried out to show the specificity of the lectins as well as the absence of detectable endogenous reactive materials. The positive control for Jacalin, PNA, and VVA was glycophorin, asialo-glycophorin, and asialo-submaxillary mucin, respectively. 12 The negative control was a human albumin preparation (Sigma) included in the test instead of milk and plasma samples. The background absorbance was low (range from 0.03 to 0.06 AU) when Tris-buffered saline was included in the tests instead of (1) lectin, (2) ExtrAvidin–alkaline phosphatase, and (3) milk or plasma samples.
Statistical analysis
The statistical analysis was performed with the STATISTICA version 10.0 software package (StatSoft, Inc., Tulsa, OK). Comparisons between groups were performed by means of the Mann–Whitney U test and the Kruskal–Wallis test. Data are presented as mean±SD values and the median with 25th–75th percentiles. The correlations were estimated according to the Spearman method. A p value of<0.05 was regarded as significant.
Results
Expression of Jacalin-, PNA-, and VVA-reactive glycotopes
AGP from all milk samples reacted with Jacalin, PNA, and VVA (Table 1).
The reactivity of 100 ng of α-1-acid glycoprotein (AGP) with A. integrifolia lectin (Jacalin) (specific to sialyl-T antigen), A. hypogaea lectin (PNA) (specific to T antigen), and V. villosa lectin (VVA) (specific to Tn antigen) is expressed as the absolute value of absorbance (absorbance units [AU]) at 405 nm based on the lectin–AGP enzyme-linked immunosorbent assay as described in Materials and Methods. Data are mean±SD values and the median (25th–75th percentiles) as indicated.
The Mann-Whitney U and the Kruskal–Wallis tests were used for statistical calculations. A p value of<0.05 was considered significant. Values that are significantly different from the groups of aearly colostrum (2–3 days), bcolostrum (4–7 days), ctransitional milk (8–14 days), and dall groups of human milk are indicated.
The reactivity of Jacalin (specific to α2,3/α2,6 sialyl-Galβ1,3GalNAc) with milk AGP remained nearly at the same level during Days 2–3 and 4–7 of lactation (0.9±0.4 AU and 1.1±0.4 AU, respectively) and then decreased within 8–14 (0.8±0.3 AU; p<0.003) and 15–45 (0.6±0.2 AU; p<0.009) days of lactation. The reactivity of PNA (specific to Galβ1,3GalNAc) with milk AGP was the lowest during Days 2–3 of lactation (0.3±0.2 AU) and increased within 4–7 (0.5±0.3 AU; p<0.043), 8–14 (0.7±0.3 AU; p<0.0002), and 15–45 (0.9±0.2 AU; p<0.03) days of lactation. In contrast, the reactivity of VVA (specific to GalNAc) with milk AGP was very low and remained almost at the same level in the early colostrum, colostrum, transitional milk, and mature milk groups (0.2±0.1 AU, 0.2±0.1 AU, 0.1±0.1 AU, and 0.1±0.1 AU, respectively) (Table 1).
In contrast to milk AGP, the plasma AGP of lactating mothers as well as the normal plasma group lacked reactivity with sialyl and asialyl T antigen- as well as Tn antigen-specific lectins (Jacalin, PNA, and VVA, respectively).
Correlations between expressed glycotopes
Milk AGP reactivity showed a weak negative correlation with milk maturation with respect to its interaction with Jacalin (r=−0.29) and VVA (r=−0.21) and a positive correlation for PNA (r=0.58) (Table 2 and Fig. 1). Additionally, the PNA reactivity showed a weak negative correlation (r=−0.24) with the reactivity of VVA (Table 2). The obtained results of AGP–lectin reactivities were statistically analyzed with already published data for human milk AGP sialylation. 11 In particular, the values obtained for human skim milk AGP reactivity with sialic acid–specific lectins were used for statistical correlation with the results presented here. The reactivity of Jacalin with AGP showed a positive correlation with the reactivity of MAA (specific to α2,3-linked sialic acid) with AGP (r=0.58; Table 2 and Fig. 2A). In contrast, the reactivity of PNA with AGP showed a negative correlation with the reactivity of SNA (specific to α2,6-linked sialic acid) with AGP (r=−0.5) (Table 2 and Fig. 2B).
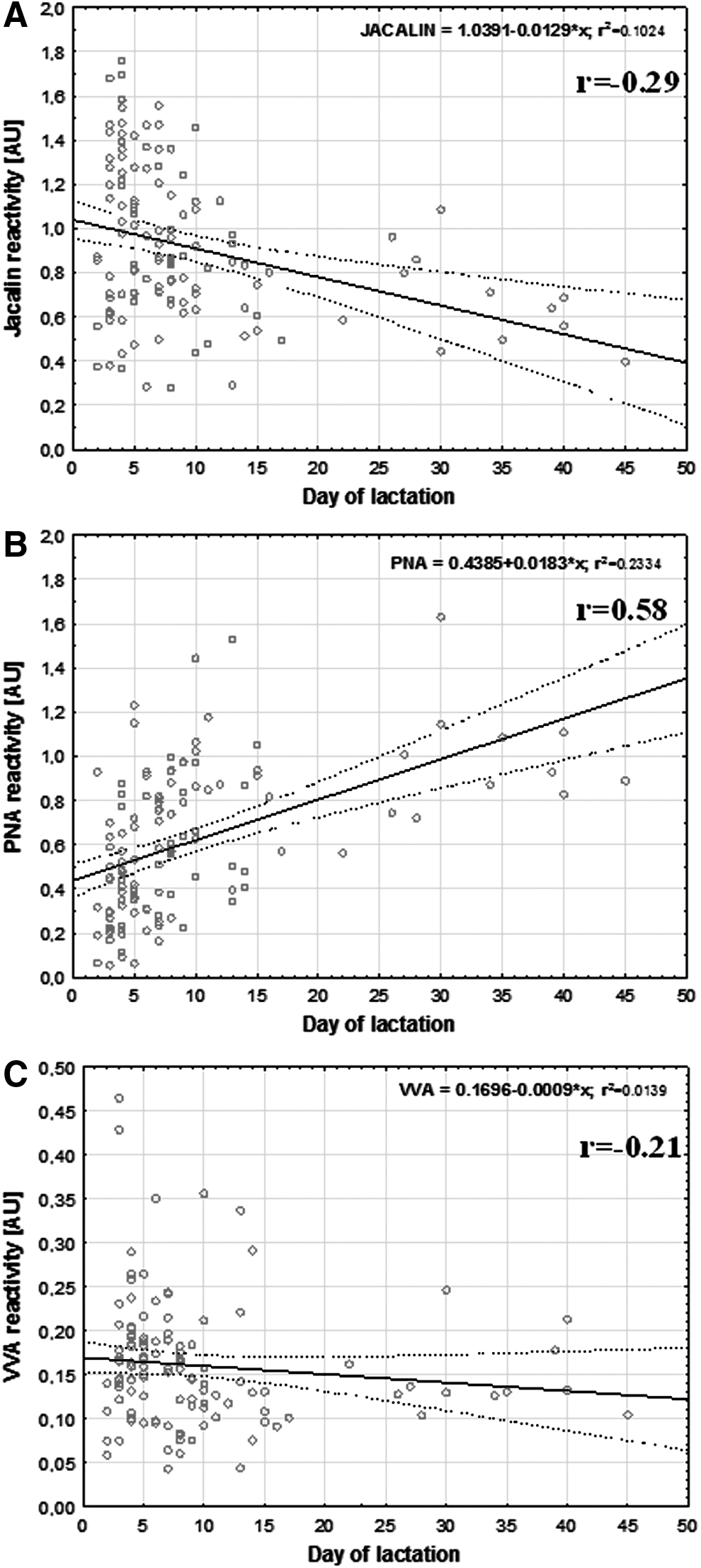
Reactivity of skim milk α-1-acid glycoprotein with
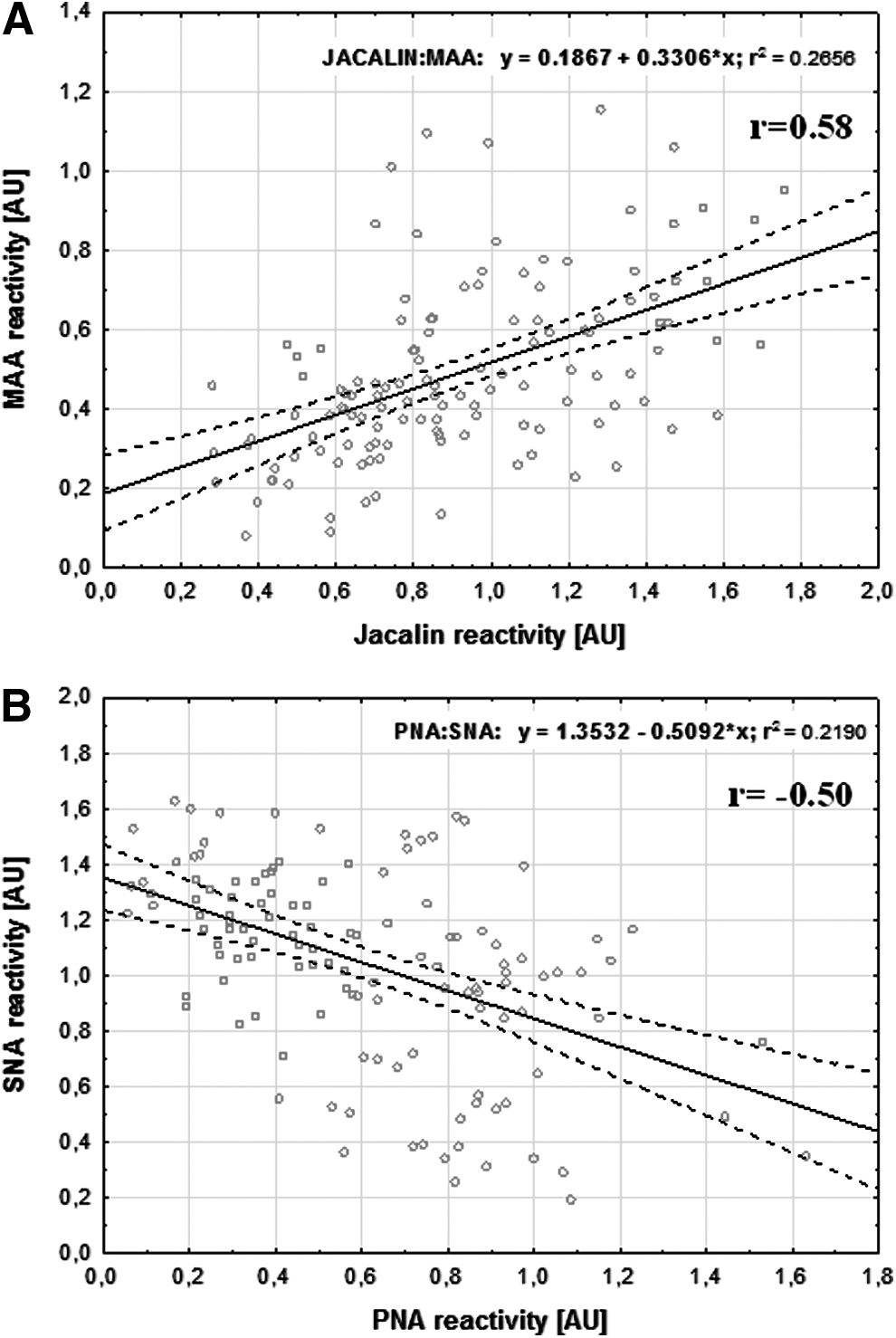
Correlation of skim milk α-1-acid glycoprotein reactivity between
The values of r calculated according to the Spearman method correspond to correlation between the relative amounts of glycotopes expressed on human milk α-1-acid glycoprotein (AGP) over milk maturation from Day 2 to Day 45 of lactation. Data of milk AGP reactivity with sialic acid–specific lectins (M. amurensis lectin [MAA] and S. nigra lectin [SNA]) were previously reported. 11 The analysis was performed for the same set of human milk samples. Jacalin-, PNA-, VVA-, MAA-, and SNA-reactive AGP are AGP glycovariants recognized by the lectins Jacalin (specific to sialyl antigen T), PNA (specific to asialyl antigen T), VVA (specific to antigen Tn), MAA (specific to α2,3-linked sialic acid), and SNA (specific to α2,6-linked sialic acid), respectively.
NS, nonsignificant correlation (p≥0.05).
Discussion
Our present analyses show that milk AGP differs from mothers' plasma AGP by the presence of O-linked oligosaccharides: sialylated and asialylated T as well as Tn antigens. Additionally, milk maturation from early colostrum to mature milk was associated with a significant gradual decrease of the relative amounts of sialylated glycoforms of T antigen, almost unchanged Tn antigen, and the occurrence of an increased amount of asialylated antigen T.
The lectin-reactive ELISA we used, which is based on similar interactions occurring in vivo between tissue lectins (selectins, galectins) and their natural ligands (sialylated and/or fucosylated epitopes on glycoconjugates), serves as a good analytical tool for deciphering a wide range of glycotopes. The advantage of the present strategy over the mass spectrometry and high-performance liquid chromatography methods preceded by enzymatic release of glycans is that they are a more convenient approach that allows for simultaneous analysis of a large number of samples at different time points bound with variable physiopathological conditions without the need for glycan release prior to analysis.11,16,30 In particular, binding of lectins to ELISA plate-isolated glycoproteins and other lectin-based methods allow rapid and exhaustive analysis and are useful for detecting even weak interactions due to multivalent glycotope–lectin interactions.11,12,29,31 Additionally, the results obtained using lectin-based methods are reported to be well correlated with mass spectrometry analysis of glycan structures. 16
The observed differences in the reactivities of human milk and plasma AGP with the panel of O-glycan specific lectins (Table 1 and Fig. 1) are closely connected with AGP origin. The human plasma AGP that originated from hepatic synthesis did not contain O-glycans. 23 In contrast, the milk AGP that originated mainly from local synthesis in the mammary gland elicited the presence of asialyl and sialyl T and Tn glycotopes. Apart from Shiyan and Bovin, 24 who reported the presence of O-glycans on AGP isolated from fetal serum, there is no report confirming the existence of O-glycans attached to this protein. Differences in O-glycosylation between serum/plasma and milk were reported for secretory immunoglobulin A 9 and fibronectin. 12 The differences are likely to be due to the different glycosylation enzymes in the plasma cells located in the bone marrow/spleen/liver from those at different mucosal surfaces. 9 The O-glycan structures and sites of their attachment differ significantly among tissues.17,20 The O-glycans are characteristic for secreted glycoproteins and mucins19,32,33 that line the upper respiratory tract epithelium and are involved in innate defense mechanisms that are based on glycan–lectin receptor interaction. The appearance of the O-linked T and Tn glycotopes on human milk AGP, which are not found on the free milk oligosaccharides,15,34 can be considered in the context of evolutionary adaptation of bioactive components of human milk to act as a part of innate immunity passed down through the mother to the breastfed newborn because they can as soluble decoys participate in preventing the adhesion of viruses and bacteria to their carbohydrate mucosal receptors or in protection against bacterial and viral diseases.3,4,9,15,35
The relative amounts of sialyl, asialyl T, and Tn glycotopes of human milk AGP were lactation stage related (Table 1 and Fig. 1). The expression of the sialylated form of T (Jacalin-reactive) and Tn (VVA-reactive) glycotopes decreased with milk maturation, in contrast to the increase of expression of the asialylated form of T glycotope (PNA-reactive). Taking into account the results of lectin–AGP reactivities and the correlation calculations (Table 2), we suggest that the decrease of the expression of T glycotope per se with milk maturation and the observed increased reactivity with PNA are closely connected with the decrease of T glycotope sialylation.
The observed lactation stage-related alterations in the expression of T and Tn antigens were probably caused by hormonal regulation of N- and O-glycan synthesis over lactation, namely, the decreasing level of estrogen after delivery. 36 The observed trends for the sialylated and asialylated glycoforms of T glycotope are consistent with the changes reported recently for sialylated O-glycans of human milk fibronectin 12 and bile salt–stimulated lipase 8 with progression of lactation. The observed trends correspond exactly to the gradually decreasing sialylation over lactation, reported for HMOs1,37 and for free as well as glycoconjugate-bound sialic acids, 38 and seem to be similar to the well-known alterations in the expression of stage-specific embryonic antigens because in human milk glycans that are related to embryonic stem cell-specific epitopes were identified. 39
Throughout lactation the secretions of mammary glands undergo a “metamorphosis” from colostrum to mature milk. Gao et al. 13 have suggested that the differentiation and function of these secretions overlap with the changes in the needs and requirements of the intensively growing and developing newborn.
The biological function of human milk AGP glycosylation is still uncertain. However, apart from the presence of O-linked oligosaccharides affecting the function and stability of AGP, the attached glycotopes may themselves have a potential biologically important role because they can constitute an additional carrier of sialic acid, which is extremely important for blocking adhesion of sialic acid–dependent pathogens. 40 Their occurrence increases the diversity of glycotopes capable of binding pathogens, adhesins, and rotaviruses, particularly those that are the leading cause of severe gastroenteritis in neonates. 40 In that way the milk sialylated and asialylated glycotopes acting as inhibitors of the above-mentioned interactions are able to influence the effectiveness of defense mechanisms of newborn innate immunity.
Conclusions
The appearance of O-linked oligosaccharides—namely, sialylated and asialylated T and Tn antigens—on human milk AGP is tissue specific and lactation stage related. However, detailed studies with pure glycoprotein are needed to provide information on how the glycoforms of human milk AGP translate into bioactive functions and potential health benefits.
Footnotes
Acknowledgments
This work was supported by grant ST-561 of the Faculty of Medicine, Wrocław Medical University, Wrocław, Poland.
Disclosure Statement
No competing financial interest exist.