
Abstract
Abstract
Background:
Women diagnosed with pregnancy-associated breast cancer postpartum have a worse prognosis, stage for stage, than other women with breast cancer. The time of breast involution is tumor promotional. The extracellular matrix protein tenascin-C is upregulated during involution in animal models and promotes breast cancer progression. It interacts with transforming growth factor (TGF)β, which also is involved in breast involution and breast cancer progression. Little is known about the expression of tenascin-C during human breast involution, nor its relationship to TGFβ. The purpose of this study was to investigate the expression of tenascin-C throughout lactation, as well as its relationship to TGFβ1 and TGFβ2.
Material and Methodology:
Three milk samples from 25 lactating women (transitional, whole, and wean) were collected, separated into components (cells, fat, and skim), and the skim fraction analyzed for total protein, tenascin-C, TGFβ1, and TGFβ2. Tenascin-C, TGFβ1, and TGFβ2 were detectable in all milk samples.
Results:
Highest tenascin-C levels on average were found in whole milk, whereas highest mean TGFβ1 and TGFβ2 levels were in wean milk. Wean samples on average had higher levels of both TGFβ1 (26%) and TGFβ2 (>500%) than matched transitional milk samples. Tenascin-C levels in wean milk were associated with nursing length (p = 0.048). Combining all three milk collection time points, tenascin-C exhibited a weak inverse correlation with TGFβ1 and TGFβ2 (p < 0.1). The inverse correlation at the wean time point was stronger for TGFβ2 than −1 (−0.37 versus −0.25). Tenascin-C, a protein known to promote breast cancer progression, is expressed throughout lactation.
Conclusion:
The inverse correlation with TGFβ2 in wean milk suggests a possible interaction during breast involution, which should be further investigated.
Introduction
P
Weaning initiates breast involution, a process in which the breast must carry out all of the steps to revert to its prepregnant state. In doing so, it recruits enzymes to break down the glandular architecture that was created during pregnancy and lactation. This microenvironment increases the concentration of proteins, which stimulate apoptosis, such as transforming growth factor (TGF)β, 4 and metalloproteinases, which break down the extracellular matrix (ECM). 5 A preclinical study identified the ECM protein tenascin-C as a potential mediator of tumor progression during involution. 5 This microenvironment, enriched in growth factors, is hypothesized to drive the conversion of precancerous lesions to cancer and the progression of preexisting cancerous lesions in the postpartum gland. 5 The tumor promotional environment at the time of breast weaning and involution is thought to contribute to this worse prognosis. 5
Analyzing ECM proteins from the involuting rodent mammary gland, Schedin and colleagues found that tenascin-C was upregulated (compared to the nulliparous mammary gland) and decreased by nonsteroidal anti-inflammatory treatment. 5 Tenascin-C is overexpressed in breast stromal tissues and in the serum of breast cancer patients 6 ; it promotes breast cancer metastases. 7 Tenascin-C influences the cell signaling pathway to promote epithelial mesenchymal transition, with the downregulation of E-cadherin and simultaneous upregulation of vimentin and several matrix metalloproteinases. This leads to increased cell motility and metastases. 8 Work by Massague et al. 9 suggests that tenascin-C promotes breast cancer metastatic initiation and progression by providing a supportive scaffold for cell survival and spread by concentrating growth factors and/or interacting with ECM components in the breast involution microenvironment. 7
TGFβ is known to regulate the transcription of at least some tenascin isoforms in breast cancer metastases; 10 however, less is known about the association between TGFβ and tenascin isoforms in the development of breast cancer, particularly during breast involution. Two publications report the analysis of tenascin-C expression in breast milk during the first 1 month 11 to 2 months 12 postpartum. We are unaware of publications that report tenascin-C expression in the breast milk of lactating women at the time of weaning.
Materials and Methods
Preparation of milk and serum samples
Three milk samples (transitional, within the first 10 days of nursing; whole, 2 months postpartum; and wean, when the mother was weaning the baby from nursing, a time of early breast involution) were collected from 25 lactating women after informed consent was obtained from the Institutional Review Boards of the University of Missouri and the University of North Dakota. The women ranged in age from 24 to 39 (median 30) years. Age, body mass index (BMI), and length of the nursing period for these women are summarized in Table 1. The samples were separated by centrifugation into the following three components: fat, supernatant (skim milk), and cell pellet, then snap frozen, labeled, and placed in a −80°C freezer until analysis.
BMI, body mass index; SD, standard deviation.
Tenascin-C, TGFβ1, and TGFβ2 analysis in milk
The skim milk fraction was analyzed for total protein, tenascin-C, TGFβ1, and TGFβ2. All analyses were performed in duplicate. To measure total milk protein, 25 μL of standard and milk samples was added in duplicate to each microplate well (Pierce), followed by 200 μL of working reagent with mixing. The microplate was then incubated at 37°C for 30 minutes and absorbance measured at 562 nm.
Tenascin-C was measured using an immunoassay kit from MyBioSource following the manufacturer's instructions. Briefly, 100 μL of standard and milk samples was added to wells in duplicate, followed by 50 μL of conjugate. The plates were incubated for 1 hour at 37°C, the plate washed, 50 μL substrate A and 50 μL substrate B added to each well, and the plate incubated for 15 minutes at 37°C. Fifty microliters of stop solution was then added and the signal read with a plate reader at 450 nm. The sensitivity of the Tenascin-C Kit is 1.0 ng/mL. The interassay coefficient of variation was <15%. R2 was 0.9996.
TGFβ isoforms
TGFβ1 and TGFβ2 protein levels in milk samples were determined by immunoassay (R&D Systems) following the manufacturer's instructions. Briefly, 100 μL of standard and milk samples was added to wells in duplicate, incubated for 2 hours at room temperature (RT), washed, and 200 μL of TGFβ1 or TGFβ2 conjugate added. Following another wash, 200 μL of substrate solution was added for 20 minutes at RT, the reaction ended with 100 μL of stop solution, and absorbance measured at 450 nm. The detection limits of the TGFβ1 and TGFβ2 kits are 4.61 and 7.0 pg/mL, respectively. The interassay coefficient of variation was <15%. R2 for TGFβ1 and TGFβ2 was 0.9990 and 0.9997, respectively.
Statistical analysis
The correlation between each pair of proteins was tested by determining the Pearson product-moment correlation coefficient. All correlations were computed from at least 20 pairs of expression values, and therefore, asymptotic distribution of the standardized Pearson correlation was assumed to be valid. A test of significance was performed using t-distributed standardized Pearson product-moment correlation coefficient. However, multiple samples from the same subject are expected to be dependent, and therefore, a statistical model accommodating for the longitudinal dependence pattern was invoked as a supplemental analysis to explain the association between the proteins. We assumed tenascin-C level to be the dependent variable and TGFβ1 or TGFβ2 to be the independent measurement in model 1 and model 2. Model 1 addresses the relationship between tenascin-C and TGFβ1, and model 2 addresses the relationship between tenascin-C and TGFβ2. Correlation within an individual was modeled through a compound symmetric covariance pattern. Reversing the role of tenascin-C and TGFβ did not affect the pattern of association. We performed multivariate analysis using a linear mixed-effects model with the subject as the random effect and phase (Transitional, Mature, or Wean) as the fixed effect, with a compound symmetric covariance pattern fit to the data. The coefficients of TGFβ1 and TGFβ2 were taken as measures of association between tenascin-C and TGFβ1 and TGFβ2. All computations were performed using software R v3.2.2.
Results
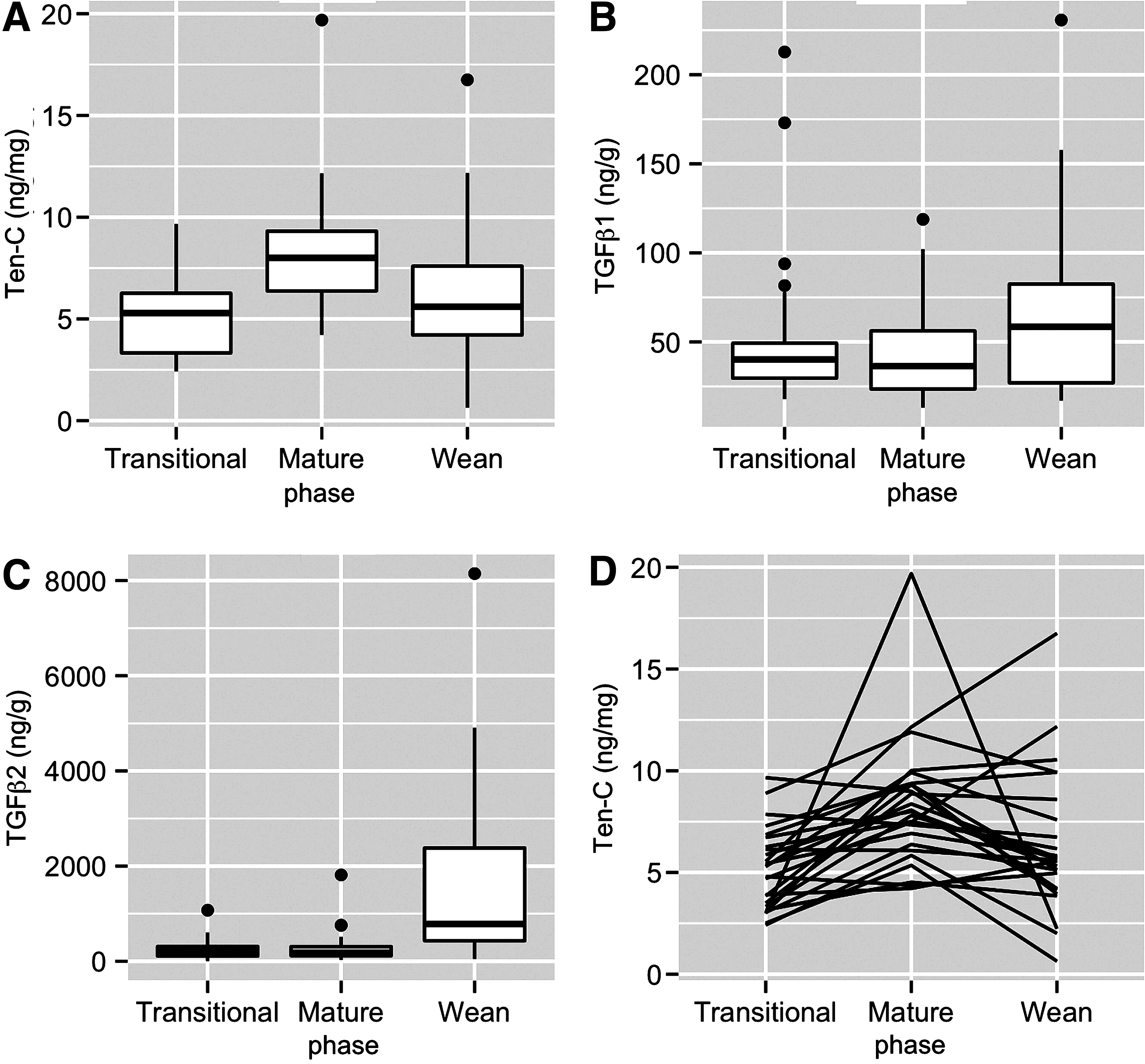
Tenascin-C was detectable in all milk samples (Table 2). The highest mean levels were found in whole milk, whether considering concentration per unit volume or controlling for total protein. Change in tenascin-C expression during lactation was highly variable, with the highest levels found in transitional milk for 3 women, in whole milk for 16 women, and in wean milk for 6 women.
Ten-C, tenascin-C; TGFβ, transforming growth factor β.
TGFβ1 and TGFβ2 were detectable in all milk samples. Mean TGFβ1 and TGFβ2 levels were highest during breast weaning (Table 2). Whereas mean TGFβ1 levels were 26% higher in wean than transitional milk, mean TGFβ2 levels were over fivefold higher in wean than in transitional samples. The summary pattern of tenascin-C, TGFβ1, and TGFβ2 expression in this cohort is demonstrated in Figure 1.
Tenascin (Ten)-C
Levels of TGFβ1 and TGFβ2 were strongly correlated and statistically significant at every stage. Tenascin-C and TGFβ1 were weakly inversely correlated at all three milk collection time points; the inverse correlation between tenascin-C and TGFβ2 was stronger in wean milk (Table 2). Tenascin-C levels in wean milk decreased (Fig. 2) as the length of breastfeeding increased (p = 0.048). When all three milk collection time points were combined, the correlations between tenascin-C and both TGFβ1 and TGFβ2 were negative and of borderline significance (Table 2). Considering the sample size of this study, this level of significance is deemed to be clinically meaningful.
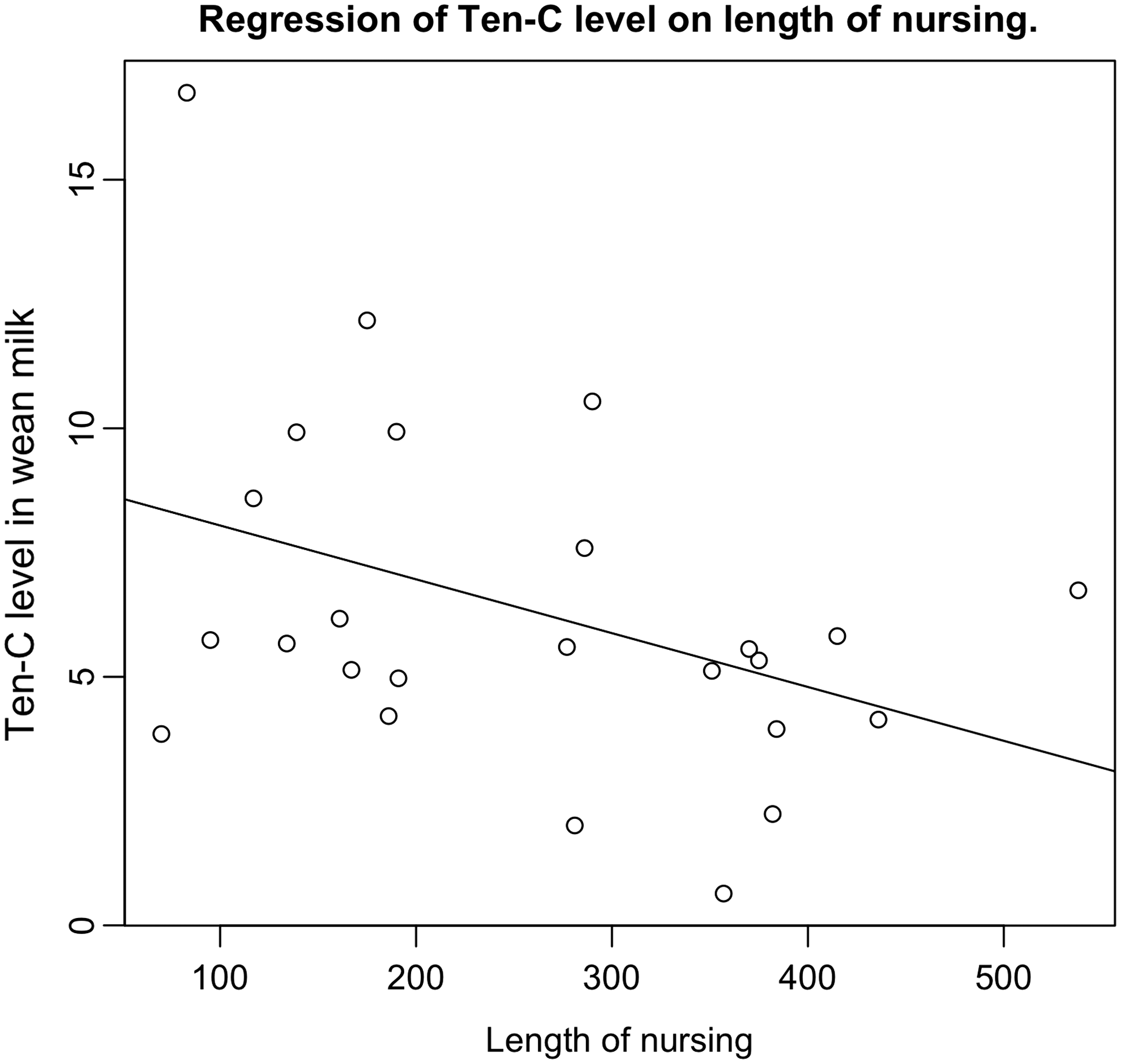
Ten-C level (ng/mg) in wean milk decreased as the length of nursing (days) increased.
Discussion
Tenascins are glycoproteins found in the ECM. Of the four vertebrate tenascins (R, C, X, and W), expression of tenascin-R is the most restricted, being found only in the central nervous system, 13 whereas tenascin-X is constitutively expressed in connective tissues. Tenascin-C and Tenascin-W expression varies in response to stimuli. 13 Involution studies in rats demonstrate increased expression of tenascin-C in the involuting mammary gland compared to the nulliparous gland. 5 Two reports document tenascin-C expression by mass spectrometry in human milk early in lactation. The first study, which evaluated milk samples from four women, documents what we feel is a clinically significant drop in tenascin-C during the first month of lactation. 11 A second study evaluated breast milk from 17 preterm mothers during the first 2 months postpartum and 8 term mothers during the first month postpartum. 12 For both preterm and term mothers, tenascin-C decreased during the first month and then leveled off during the second month.
We report, for the first time to our knowledge, tenascin-C levels in human breast milk collected during breast weaning/early breast involution. We observed the highest mean tenascin-C expression in whole milk, although for an individual participant, the change over time was quite variable, with 24% (6/25) of women having the highest expression of tenascin-C in their wean milk sample (Table 1 and Fig. 1D). We did not observe a unique characteristic, such as age, family history of breast cancer, BMI, or age at first full-term pregnancy, which differentiated these six from the other 19 participants. We searched for other characteristics (sleep habits and diet), which might influence tenascin-C levels. We were unable to find a publication linking tenascin-C to sleep habits. We found an animal study 14 suggesting that a high-fat diet may influence tenascin-C expression. We were unable to find a human study documenting this.
We observed a trend toward increasing tenascin-C in whole milk compared to transitional milk, unlike two prior publications,11,12 which observed a decrease during the first month and then a leveling off between months 1 and 2 after childbirth. Possible explanations for the differences include random variability due to the small size of the studies, the different method of breast milk analysis (the first two studies by mass spectrometry, whereas we used an immunoassay), and the time of collection (the two studies collected milk very early after childbirth, whereas our transitional milk collection was generally between days 5 and 10 after childbirth). In contrast, we identified a significant inverse correlation between tenascin-C and length of nursing (Fig. 2). Epidemiologic findings are mixed regarding the influence of nursing length on future breast cancer risk, with some 15 but not all 16 studies supporting a protective effect with increasing length of nursing. Our findings are consistent with a protective effect of longer nursing length.
TGFβ is linked to breast cancer in preclinical models 17 and to prognosis in human breast cancer. 18 TGFβ levels increase during mammary gland lactation and involution, 17 and it mediates proapoptotic effects during involution. 19 Transgenic mice that overexpress TGFβ show increased apoptosis throughout mammary development. 20
Tenascins are known to activate TGFβ isoforms, 21 which are important mediators of breast involution and breast cancer progression. 19 Most reports documenting an association between TGFβ isoforms and tenascins have evaluated TGFβ1. We evaluated both TGFβ1 and TGFβ2 in the same milk samples as tenascin-C. We observed a stronger association between tenascin-C, a protein linked to breast cancer, and TGFβ2 than TGFβ1. This observation is consistent with our earlier observations of TGFβ2 upregulation in the cancer-containing breast of nursing women compared to their matched normal breast 22 and in wean milk compared to milk collected earlier in lactation, 23 as during breast weaning and involution, the breast microenvironment is tumor promotional. 5 Our results suggest an inverse correlation between tenascin-C and TGFβ. The lack of significance between tenascin-C and TGFβ2 in wean milk, with a correlation coefficient of −0.37, may be due to the limited sample size. It is possible that tenascin-C is turned off with the activation of TGFβ or that the tumor promotional properties of tenascin-C compete with the apoptosis-inducing properties of TGFβ. 19
Conclusion
Tenascin-C is expressed in human breast milk throughout lactation, including during breast weaning/early involution. Levels during involution are inversely correlated with TGFβ2, suggesting a possible interaction. Since tenascin-C is known to promote the progression of breast cancer and TGFβ2 is thought to induce cellular apoptosis during involution, further study of this interaction is warranted.
Footnotes
Acknowledgments
Benji Hawkins assisted with collection of all milk samples. Funding for this project was provided by the Avon Foundation for Women (Grant No. 02-2012-090).
Disclosure Statement
No competing financial interests exist.