
Abstract
Abstract
Background:
Chemokine C-C motif ligand 20 (CCL20) is implicated in the formation and function of mucosal lymphoid tissues. Although CCL20 is secreted by many normal human tissues, no studies have evaluated the presence of CCL20 in human milk or its production by oral keratinocytes stimulated by human milk.
Objective:
To evaluate the presence of CCL20 in breast milk and verify CCL20 secretion in vitro by oral keratinocytes stimulated with human and bovine milk, as well as its possible association with breast milk lactoferrin levels.
Materials and Methods:
The levels of CCL20 and lactoferrin were measured by enzyme-linked immunosorbent assay in human milk at three different stages of maturation from 74 healthy breastfeeding mothers. In vitro, oral keratinocytes were stimulated with human and bovine milk, and CCL20 was measured in their supernatant.
Results:
High concentrations of CCL20 were detected in the human breast milk samples obtained during the first week (1,777.07 pg/mL) and second week postpartum (1,523.44 pg/mL), with a significantly low concentration in samples at 3–6 weeks postpartum (238.42 pg/mL; p < 0.0001). Human breast milk at different weeks postpartum stimulated higher CCL20 secretion by oral keratinocytes compared with bovine milk (p < 0.05). Such stimulation had no association with breast milk lactoferrin concentration.
Conclusion:
CCl20 is present at high levels in human milk, predominantly in the first and second week postpartum, but at significantly lower levels at 3–6 weeks postpartum. Human milk is capable of stimulating CCL20 secretion by oral keratinocytes, and this induction had no association with breast milk lactoferrin concentration.
Introduction
T
Secretion of CCL20 could be stimulated by microbial factors, including lipopolysaccharide and inflammatory cytokines, such as tumor necrosis factor-alpha (TNF-α) 8 and IL-1β.9,10 Another possible candidate for the stimulation of CCL20 secretion is lactoferrin (Lf), with its cleavage products. 11 Lf is a major breast milk component and it plays an important role in unspecific immunity. Contrasted to this protector effect, studies have shown that some cleavage peptides of this protein have strong proinflammatory activity in mucosal membranes.12,13
CCL20 could play an important role in mucosal homeostasis and viral entry. CCL20 is a chemokine that attracts LCs, known as HIV target cells, which act predominantly in the initial transmission stage.1,14,15
No studies have evaluated the presence of CCL20 in human milk. Considering the importance of CCL20 in the formation and function of mucosal lymphoid tissues and chemoattraction of immature dendritic cells, we aimed to evaluate the presence of this chemokine in human milk, as well as its production, in vitro, by oral mucosal epithelial cells stimulated by human milk in different maturation stages and by bovine milk. We also verified the possible correlation between human milk Lf concentration and CCL20 secretion by oral keratinocytes.
Materials and Methods
Subject population
From January 2014 to January 2015, 74 nursing mothers were recruited during their postpartum visits at Vila Lobato Basic Health Unit and the Human Milk Bank of the Clinical Hospital of Ribeirão Preto School of Medicine, University of São Paulo, Brazil. The study protocol was approved by the local Research Ethics Committee (protocol no. 4616/2013), and all of the subjects signed written informed consent forms to participate.
The breastfeeding mothers were enrolled on the basis on the following criteria: ≥18 years old and not presenting pain, fissures, mastitis, or other alterations that would interfere with milk collection.
Collection of human breast milk samples
One breast milk sample was collected from each of the 74 lactating woman. The milk samples were classified according to the postpartum weeks 16 : week 1: 28 breast milk samples from lactating women in the first week postpartum; week 2: 16 milk samples from lactating women in the second week postpartum; weeks 3–6: 30 breast milk samples from lactating women at 3–6 weeks postpartum. Nurses collected the breast milk samples manually after lubrication of the skin and nipples by the subject's milk. The collected amount was ∼4 mL. The breast milk samples were obtained in sterilized polyethylene containers identified with a code and were transported on ice in an isothermal box. Subsequently, all of the samples were centrifuged (1000 g) for 5 minutes, and the whey fraction was separated in aliquots and stored at −80°C until processing.
Bovine milk was commercially obtained and centrifuged following an identical protocol used for human milk.
Measurement of CCL20 in human breast milk
The breast milk whey fractions were diluted 1:10 in phosphate-buffered saline (PBS), and CCL20 was detected using a commercial solid-phase sandwich ELISA (enzyme-linked immunosorbent assay) kit (Quantikine; R&D Systems, Abingdon, UK), according to the manufacturer's recommendations. The assay range is 7.8–500 pg/mL with sensitivity of 0.87 pg/mL. The intra- and interassay coefficients of variation are 2.8% and 7.8%, respectively. The optical density was measured at 450 nm with wave correction of 570 nm in an iMark micro plate reader (Bio-Rad, Hercules, CA).
The measurement of CCL20 secretion by SCC-9 cells
The SCC-9 (American Type Culture Collection, Manassas, VA) cells, a human oral squamous carcinoma keratinocyte lineage of cells, were cultivated in Dulbecco's minimal essential medium (DMEM-F12 medium; Cultilab, Campinas, Brazil), supplemented with 10% fetal bovine serum (FBS) (Cultilab), 50 μg/mL of vancomycin, 50 μg/mL of gentamicin, and 0.4 μg/mL of hydrocortisone (Sigma-Aldrich, St. Louis, MO). In the stimulation experiments, the SCC9 cells were seeded onto a 12-well culture plate (TPP, Switzerland), at a density of 7.104 cells per well, and cultured for 2 days until 90–100% confluence was observed.
The SCC-9 cells were incubated with 100 μL of the whey fraction of bovine milk or 100 μL of the whey fraction of human breast milk samples diluted 1:10 in DMEM-F12 culture medium supplemented with 2% FBS. The control cells were treated with recombinant human IL1-β (Peprotech, Neuilly-sur-Seine, France) at a concentration of 25 ng/mL in DMEM-F12 culture medium and 2% FBS (the positive control) or DMEM-F12 culture medium supplemented with 2% FBS only (the negative control). After an overnight treatment at 37°C and 5% CO2, the secretion of CCL20 was quantified in the SCC-9 supernatant, using a commercial ELISA immunoassay (Quantikine; R&D systems), following the identical protocol of the CCL20 measurement in human breast milk. The lower limit of detection is 7.8 pg/mL, with sensitivity of 0.87 pg/mL. The intra- and interassay coefficients of variation are 3.1% and 9.1%, respectively.
The direct CCL20 detection in the breast milk whey and the supernatant was performed using the identical ELISA plate in a paired manner. To determine the CCL20 secretion by the SCC-9 cells, the CCL20 concentration in the breast milk whey was deducted from the CCL20 concentration in the supernatant.
The measurement of Lf in breast milk
The Lf was measured in human breast milk in different weeks postpartum by ELISA. A standard curve was prepared using different Lf concentrations from human milk (Sigma-Aldrich). In total, 100 μL of breast milk (diluted 1:10,000 in PBS) was distributed per well and incubated at 4°C overnight. Nonspecific binding was blocked by adding 200 mL of 10% albumin from chicken egg whites (Sigma-Aldrich) to each well, at 37°C for 2 hours. Human Lf was detected with rabbit anti-human lactoferrin antibodies (L3262; Sigma-Aldrich), incubated for 2 hours at 37°C, and peroxidase- conjugated anti-rabbit antibodies (Sigma-Aldrich), incubated for 40 minutes at the ambient temperature. Between these stages, four PBS washes with 0.05% Tween 20 (Sigma-Aldrich) were performed. We used o-phenylenediamine dihydrochloride (OPD; Sigma-Aldrich) as the substrate, and the optical density was measured at 492 nm in an iMark microplate reader (Bio-Rad). Each sample was evaluated in duplicate. The assay range was 39–2,500 pg/mL. The intra-assay of variation was calculated by finding the variation of results in four samples of known concentration tested on one plate. The interassay coefficient of variation was calculated by finding the variation of results in four samples of known concentration in separate assays (3 plates). The intra- and interassay coefficients of variation were 3.9% and 4.6%, respectively.
Statistical analysis
The results are shown as the mean ± standard deviation. The statistical analysis was performed using GraphPad Prism software (San Diego, CA). The normality of data distribution was examined using Kolmogorov–Smirnov test. Outliers were not removed from a data analysis. The Kruskal–Wallis test and Dunn's post-test were used to compare the Lf and CCL20 concentrations in the different breast milk maturation stages and the CCL20 concentrations in the cellular supernatant resulting from the bovine milk stimulation, positive control, negative control, and breast milk at different maturation stages. The correlations of the CCL20 and Lf concentrations with the number of postpartum days as well as between the Lf concentration in breast milk and the CCL20 secretion by the SCC-9 cells were verified using Spearman's test; p < 0.05 was considered statistically significant in all of the tests.
Results
The demographic characteristics of the 74 breastfeeding women, with a median age of 30 years (ranged 18–40 years), enrolled in the study are shown in Table 1.
Group 1: Lactating women in the first week postpartum; Group 2: Lactating women in the second week postpartum; Group 3: Lactating women at 3–6 weeks postpartum.
A high concentration of CCL20 was found in human breast milk
Human breast milk has high CCL20 concentrations in breast milk samples obtained during the first 1 to 2 weeks postpartum, at 1,777 (±1,547) pg/mL and 1,523 (±1,083) pg/mL respectively, and the concentration is gradually reduced at 3–6 weeks postpartum, at 238 (±437) pg/mL (p < 0.0001, the Kruskal–Wallis test).
The CCL20 breast milk concentration had a negative correlation with the infancy postpartum days (r: −0.6840; CI: −0.7918 to −0.5350; p < 0.0001, Spearman's coefficient), as shown in Figure 1.
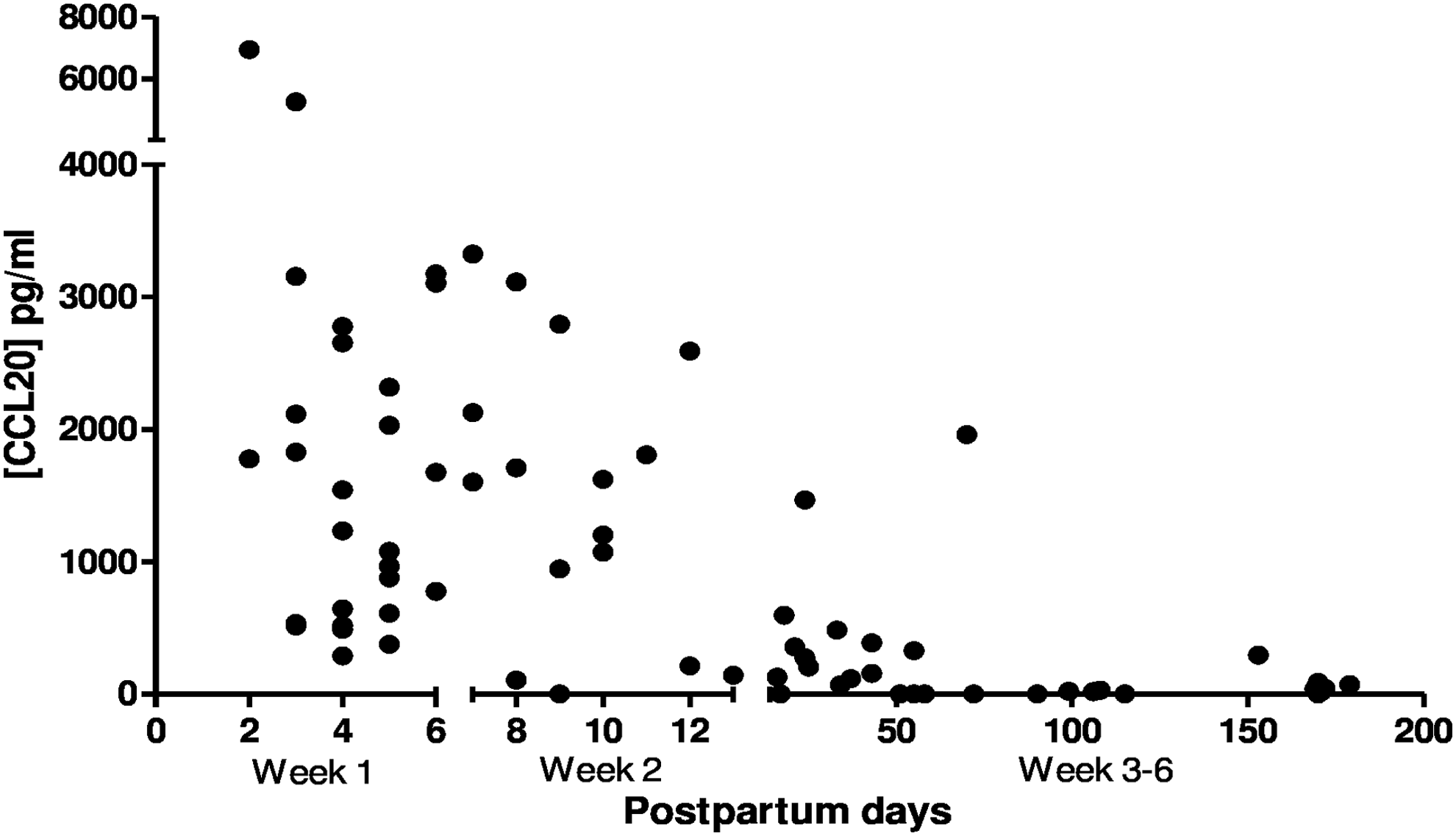
The correlation between the postpartum days and CCL20 concentration in breast milk. r: −0.6840; IC: −0.7918 to −0.5350; p < 0.0001, Spearman's coefficient. Week 1: 28 breast milk samples from lactating women in the first week postpartum; Week 2: 16 milk samples from lactating women in the second week postpartum; Weeks 3–6: 30 breast milk samples from lactating women at 3–6 weeks postpartum. CCL20, chemokine C-C motif ligand 20.
Breast milk Lf measuring
As with CCL20, high concentrations of Lf were detected in the initial stages of breast milk maturation, correlating negatively with the postpartum days (r: −0.6350; CI: −0.7541 to −0.455; p < 0.0001, Spearman's coefficient) (Fig. 2). The breast milk from first and second week postpartum presented mean Lf concentrations of 12,754 (±1,960) μg/mL and 10,366 (±1992) μg/mL, respectively, whereas the levels decreased to 7,285 (±3,383) μg/mL at 3–6 weeks postpartum (p < 0.0001—the Kruskal–Wallis test).
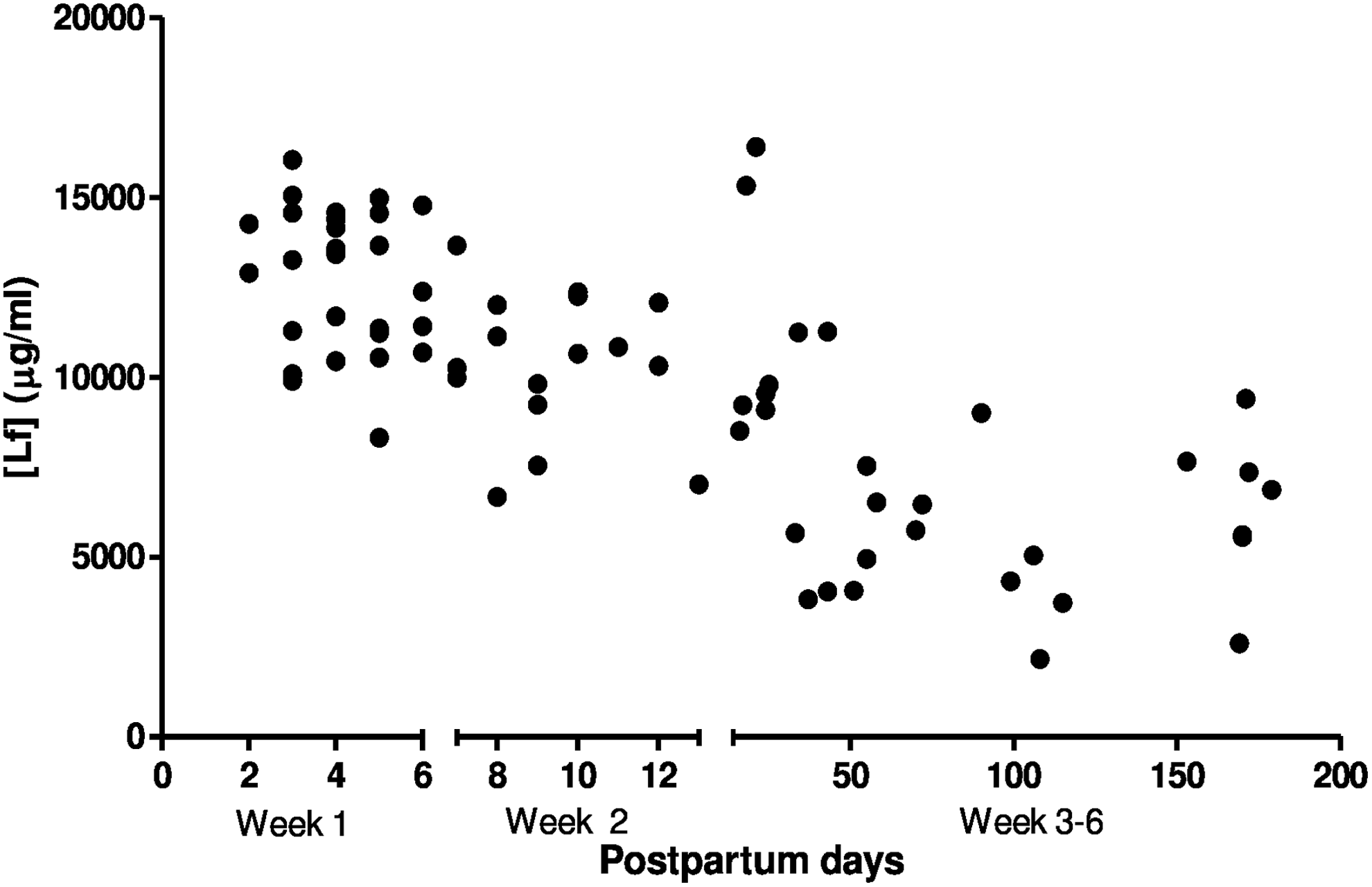
The correlation between the postpartum days and Lf concentration in breast milk. r: −0.6350; IC: −0.7541 to −0.455; p < 0.0001, Spearman's coefficient. Week 1: 28 breast milk samples from lactating women in the first week postpartum; Week 2: 16 milk samples from lactating women in the second week postpartum; Weeks 3–6: 30 breast milk samples from lactating women at 3–6 weeks postpartum. Lf, lactoferrin.
Human breast milk promotes a greater induction of CCL20 secretion by SCC-9 cells when compared to bovine milk and a negative control
As shown in Figure 3, there was no CCL20 basal secretion by the nontreated SCC-9 cells (negative control). The positive control (IL1-β) was responsible for a mean CCL20 secretion of 261.44 ± 89.89 pg/mL (p < 0.05). The fractions of the human breast milk whey in different weeks postpartum stimulated CCL20 secretion by the SCC-9 cells in comparison to the nontreated cells (p < 0.05). The breast milk from the first week stimulated the CCL20 secretion by the SCC-9 cells in 136.60 (±168.4) pg/mL, and the milk from the second and 3–6 weeks postpartum stimulated this production in 111.23 (±105.25) pg/mL and 84.11 (±58.81) pg/mL, respectively, (p > 0.05).
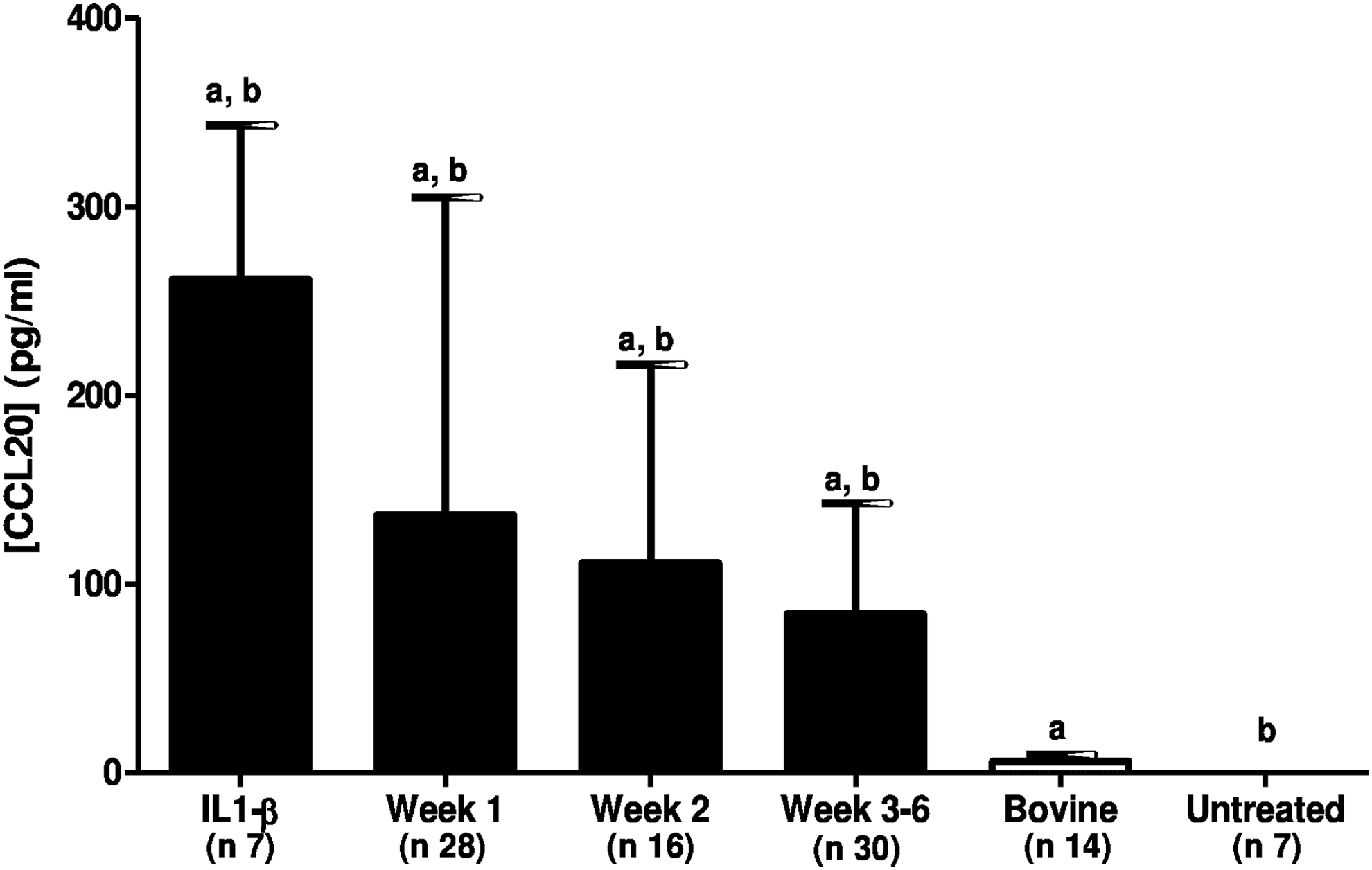
The CCL20 production by SCC-9 cells, after overnight stimulation with IL1-β (the positive control), human breast milk whey in three different stages of maturation, bovine milk whey, and nontreated cells. aThe statistically significant difference between the CCL20 concentration of human milk in different weeks and bovine milk. bThe statistically significant difference between the CCL20 concentration in different stages of human milk and the negative control. n = number of experiments. Week 1: breast milk samples from lactating women in the first week postpartum; Week 2: breast milk samples from lactating women in the second week postpartum; Weeks 3–6: breast milk samples from lactating women at 3–6 weeks postpartum. CCL20, chemokine C-C motif ligand 20.
The CCL20 secretion by bovine milk was 5.9 (±3.8) pg/mL, which was similar to that in the nontreated cells (p > 0.05) because of less stimulation compared with the CCL20 production by human milk (p < 0.05). The human breast milk from the first, second, and 3–6 weeks postpartum were responsible for a higher CCL20 production than the bovine milk (p < 0.05), as shown in Figure 3.
The induction of CCL20 secretion by SCC-9 cells had no correlation with the human breast milk Lf concentration
There was no correlation between the breast milk Lf concentration and the CCL20 secretion by SCC-9 cells. The Lf concentrations and its capacity of induce CCL20 secretion by the SCC-9 cells are shown in Figure 4 (r: 0.022; CI: −0.213 to 0.256; p = 0.849, Spearman's coefficient).
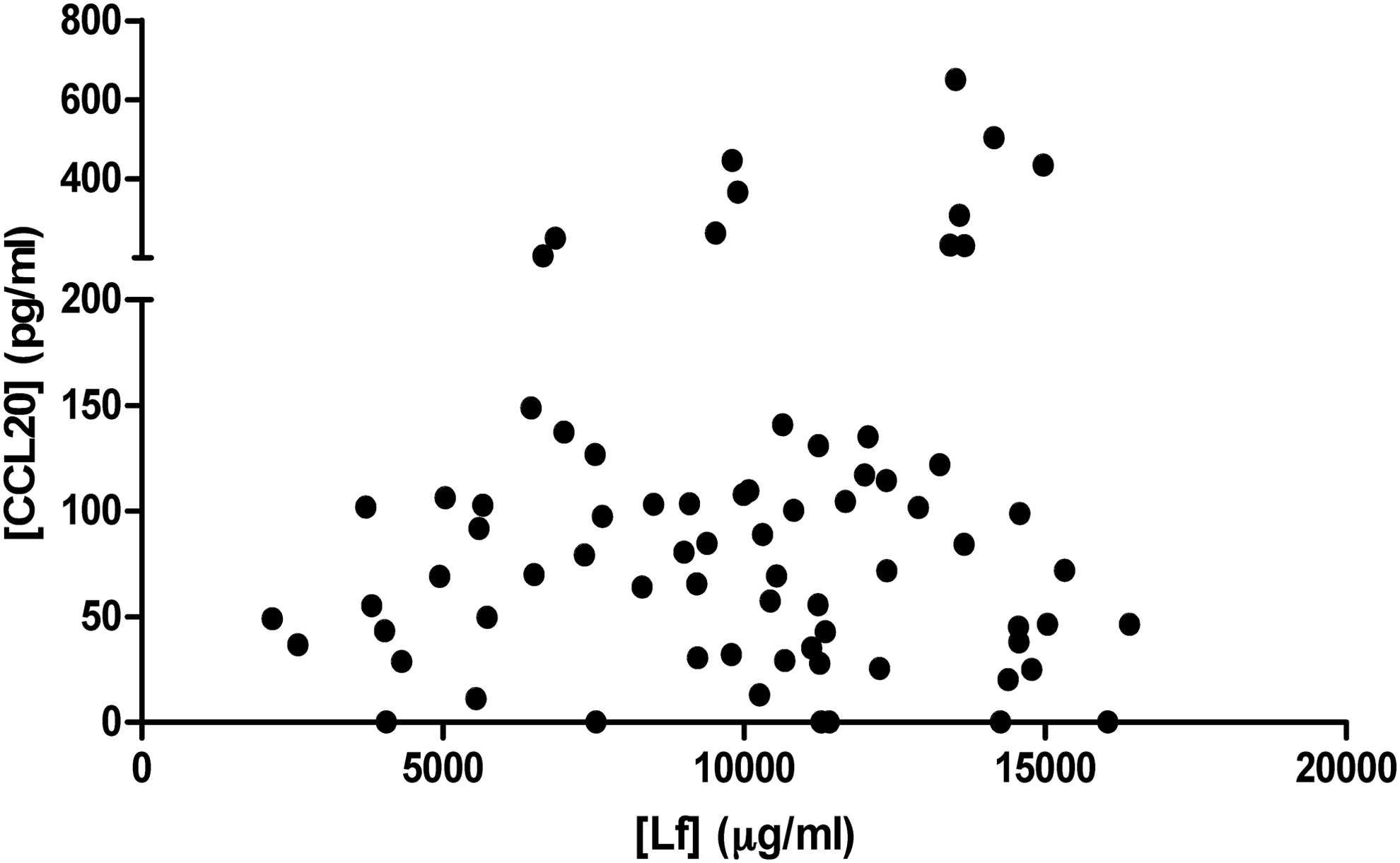
The correlation between the breast milk Lf concentration and CCL20 secretion by SCC-9 cells. r: 0.022; IC: −0.213 to 0.256; p = 0.849, Spearman's coefficient. CCL20, chemokine C-C motif ligand 20; Lf, lactoferrin.
Discussion
To the best of our knowledge, this study is the first evaluation of the detection of CCL20 in human breast milk and its production by oral epithelial cells under stimulation by human milk. We found high CCL20 concentrations in breast milk in the first and second week postpartum with decreasing concentrations at 3–6 weeks. In addition, we verified that human milk is capable of stimulating higher CCL20 secretion by oral keratinocytes in a comparison with bovine milk; however, the induction of CCL20 secretion had no direct correlation with the Lf concentration present in human milk.
Our results, which showed the decline of CCL20 concentration in mature stages of breast milk, are similar to those of previous studies that verified high concentrations of other cytokines and chemokines in colostrum when compared with mature milk.17,18 The presence of high levels of CCL7 and CCL27 chemokines in paired breast milk samples was verified for the first time by Radillo et al., in 2013. 19 According to the authors, colostrum contained higher CCL7 and CCL27 concentrations when compared with mature milk. Although only one breast milk sample was obtained from each lactating mother in our study, we were also able to verify higher CCL20 levels in the early stages of breast milk maturation.
As with CCL20, high concentrations of Lf were detected in breast milk in the first and second week postpartum, but its levels decreased at 3–6 weeks after delivery. These results are consistent with those of other studies that also verified low levels of Lf in breast milk shortly after postpartum.20,21
The important role of breast milk cytokines and chemokines in infants' immune system regulation and development has been well documented.17,22 High levels of immunological components have been found in breast milk, predominantly in colostrum. Colostrum also contains relatively low concentrations of lactose and glucose, 23 indicating that colostrum plays both an important immunological and nutritional role. 24
In this study, the CCL20 secretion had no direct correlation with the Lf concentration present in breast milk, which differs from the 2014 findings of Lourenço et al. 11 in a study of CCL20 production by vaginal epithelial cells stimulated by seminal plasma, which suggested that this chemokine production could be induced by other breast milk constituents. There is evidence that cleaved inflammatory Lf peptides are responsible for the induction and production of IL-6, MCP-1, and IL-8 cytokines by oral epithelial cells and Lf cleavage products with different molecular weights could be induced in mastitis or other inflammatory processes for proteolysis.12,13 In this study, the participants were healthy breastfeeding mothers without mastitis, and the intact human Lf did not generate cleaved inflammatory Lf peptides on its own.
Other cytokines recognized as capable of stimulating CCL20 production by epithelial cells, such as IL-1β and TNF-α,8–10 have been found in human milk.17,25 High levels of these cytokines are present in colostrum when compared with mature breast milk;17,18 however, additional studies are necessary to verify breast milk components responsible for stimulating CCL20 secretion.
With results that differed from those found in our study, which utilized oral epithelial cells, Lane et al. 26 verified that gut epithelial cells could increase CCL20 expression with bovine milk oligosaccharides in comparison with gut epithelial cells stimulated by human milk oligosaccharides. We analyzed CCL20 production in human and bovine total milk serum, and recent studies have shown that human milk serum has a site-specific microbiota, 27 mostly composed of lactobacilli, lactococci, and enterococci. 28 Some of these microorganisms (e.g., lactobacilli) can induce CCL20 production by epithelial cells, 29 which could explain the higher CCL20 secretion by oral epithelial cells in human milk, once commercially available pasteurized bovine milk was utilized.
CCL20 is involved in the inflammatory process as a mediator in different diseases. 30 A potential contribution of our study may be its applicability in HIV transmission studies. Several works have postulated a correlation between CCL20 and HIV sexual transmission.3,14,15,31 The presence of CCL20 in human milk in the oral cavity of infants or CCL20 secretion by oral epithelial cells might facilitate HIV entrance because of the strong chemoattraction role of LCs, 1 which are known to be HIV target cells. 32
Conclusions
Our results showed that CCL20 is present in high concentrations in human milk, predominantly in breast milk during the first and second week postpartum, and there is a significant decline in the concentration at 3–6 weeks postpartum.
Human milk is capable of stimulating higher CCL20 secretion by oral keratinocytes when compared with pasteurized bovine milk, and this induction had no direct correlation with Lf concentration in breast milk.
Footnotes
Acknowledgments
We are grateful to all mothers who participated in the study. We also acknowledge Professor Paulo Tambasco (University of São Paulo) for providing the SCC-9 cells. This study was supported by funds from Coordination of Improvement of Higher Level Personnel (CAPES).
Disclosure Statement
No competing financial interests exist.