
Abstract
Abstract
Background:
Bioactive proteins from milk fat globule membrane (MFGM) play extensive roles in cellular processes and defense mechanisms in infants. The aims of this study were to identify differences in protein compositions in human and caprine MFGM using proteomics and evaluate possible nutritional benefits of caprine milk toward an infant's growth, as an alternative when breastfeeding or human milk administration is not possible or inadequate.
Materials and Methods:
Human and caprine MFGM proteins were isolated and analyzed, initially by polyacrylamide gel electrophoresis, and subsequently by quadrupole time-of-flight liquid chromatography–mass spectrometry. This was then followed by database search and gene ontology analysis. In general, this method selectively analyzed the abundantly expressed proteins in milk MFGM.
Results:
Human MFGM contains relatively more abundant bioactive proteins compared with caprine. While a total of 128 abundant proteins were detected in the human MFGM, only 42 were found in that of the caprine. Seven of the bioactive proteins were apparently found to coexist in both human and caprine MFGM.
Results/Discussion:
Among the commonly detected MFGM proteins, lactotransferrin, beta-casein, lipoprotein lipase, fatty acid synthase, and butyrophilin subfamily 1 member A1 were highly expressed in human MFGM. On the other hand, alpha-S1-casein and EGF factor 8 protein, which are also nutritionally beneficial, were found in abundance in caprine MFGM. The large number of human MFGM abundant proteins that were generally lacking in caprine appeared to mainly support human metabolic and developmental processes.
Conclusion:
Our data demonstrated superiority of human MFGM by having more than one hundred nutritionally beneficial and abundantly expressed proteins, which are clearly lacking in caprine MFGM. The minor similarity in the abundantly expressed bioactive proteins in caprine MFGM, which was detected further, suggests that it is still nutritionally beneficial, and therefore should be included when caprine milk-based formula is used as an alternative.
Introduction
H
Caprine (or goat) milk, on the other hand, is less allergenic and has been reported to demonstrate higher buffering capability and digestibility than bovine milk.5,6 Due to these advantages, many researchers have recommended caprine milk as a substitute, particularly in the treatment of infants suffering from cow's milk protein allergy (CMPA). 7 However, caprine milk has been far less studied than bovine milk, despite being reported as the most consumed milk in developing countries worldwide. 8 When compared to bovine milk, caprine milk generally contains smaller fat globules and different casein composition, which apparently make it a lot more tolerable to infants compared to bovine milk.6,9
Milk fat globule membrane (MFGM) is the membrane surrounding droplets of lipids during their secretion in the alveolar lumen of a lactating mammary gland. There are increasing evidences to show that MFGM proteins contribute significantly to the cognitive development and immunity of developing infants.10,11 However, the powdered form of milk-based formulas is usually prepared from whey and skimmed milk concentrate, which do not contain cream, fat, or their MFGM proteins.2,12,13 To make up for the loss of essential fatty acids, milk formulas are usually supplemented with vegetable oils to provide an overall fatty acid profile that is similar to that of human milk.14,15 The aims of this study were to identify differences in human and caprine MFGM abundantly expressed protein compositions using proteomics analysis and evaluate possible nutritional benefits of caprine milk toward an infant's growth, as an alternative when breastfeeding or human milk administration is not possible or inadequate.
Materials and Methods
Ethics
The research protocol and consent procedure of this study have been approved by the Medical Ethics Committee of the University of Malaya (Ethical approval No. 435.18).
Human milk samples
Expressed human milk samples were collected from healthy donors between 26 and 40 years of age (n = 20, 50 mL per individual), who were in their 2nd–4th month of lactation. The donors were recruited through the assistance of the Malaysian Breastfeeding Support Centre (susuibu.com). Donors were verbally explained the purposes, procedures, and benefits of the study in their preferred language. They were given ample time to ask questions for clarification and to consider their participation in the research. Once verbal consent was acquired, the donor's written consent and information regarding the age of mother and child, frequency of breastfeeding, and health status were obtained through a set of questionnaires.
Caprine milk samples
Fresh caprine milk at mid-lactation stage was purchased from a small local farm rearing 30 goats of Shami breed in Kampung Sungai Lalah, Negeri Sembilan, Malaysia (GPS coordinates: 2.609098, 102.029539).
Preparation of MFGM proteins
MFGM proteins were isolated according to the previously described method of Pisanu et al. 16 Twenty milliliters of pooled milk samples from each respective species were centrifuged at 10,000 g for 15 minutes at 4°C to separate cream fraction from whey. Tubes were incubated on ice for 20 minutes and the cream was collected upon removal of aqueous fraction. The cream was subsequently washed twice using phosphate-buffered saline at 37°C for 10 minutes with slow agitation, and once using double-distilled water. Each washing step was followed by centrifugation at 5,000 g for 15 minutes at 4°C. Fat globules were obtained after removal of the aqueous layer and stored at 4°C overnight. They were then homogenized in double-distilled water using a hand homogenizer (TissueRuptor®; Qiagen, Valencia, CA) for 8 minutes at a frequency of 60 Hz. The homogenized fractions were heated for 30 minutes at 45°C followed by centrifugation at 10,000 g for 15 minutes at 4°C to separate fat from MFGM proteins. MFGM proteins in the pellet were solubilized in double-distilled water, estimated for their protein concentrations (Bio-Rad, Hercules, CA), and finally stored at −20°C.
Polyacrylamide gel electrophoresis in the presence of sodium dodecyl sulfate
Polyacrylamide gel electrophoresis in the presence of sodium dodecyl sulfate (SDS-PAGE) was conducted according to the method of Spertino et al. 17 The samples were reconstituted individually in SDS sample buffer and heated at 100°C for 5 minutes before electrophoresis. Subsequently, 60 μg of protein from each sample was loaded onto a 10% homogenous acrylamide gel. Each sample was run in triplicate and electrophoresis was performed at a constant voltage of 90 V. The gel was then stained with Coomassie Brilliant Blue R (Sigma-Aldrich).
In-gel trypsin digestion
Lanes of SDS-PAGE gels containing the samples were individually excised into 30 pieces. The gel slices were stored in clean microcentrifuge tubes containing double-distilled water before trypsin digestion using the protocol that was previously described by Seriramalu et al. 18 Tryptic digested samples were desalted using ZipTip® containing C18 reversed-phase resin (Millipore) according to the manufacturer's instruction. The samples were then lyophilized using a vacuum concentrator.
Nano liquid chromatography mass spectrometry quadrupole time-of-flight analysis and database search
Lyophilized digested samples were dissolved in 10 μL of 0.1% formic acid and separated using a nano liquid chromatography mass spectrometry quadrupole time-of-flight (LC/MS Q-TOF) system (1260 Infinity Nanoflow LC; Agilent, Santa Clara, CA), connected to a nano electrospray ionization source (Accurate-Mass Q-TOF 6550; Agilent) as described by Ashrafzadeh et al. 19 Samples were loaded into a large-capacity HPLC chip Column (Part No. G4240-62010, Agilent). Two mobile phases, mobile phase A (0.1% formic acid in H2O) and mobile phase B (0.1% formic acid in 100% acetonitrile), were used to form a 5–80% linear gradient (of mobile phase B) for 34 minutes (flow rate = 0.4 μL/minute). The overall MS run was performed in triplicates for the purpose of quantitative analysis and the data were acquired using Mass Hunter Workstation (version B.05.1; Agilent) according to the following parameters: MS scan range of 200–3,000 m/z; collision-induced dissociation of the two most intensive ions; and MS/MS scan range of 5–3,000 m/z.
The acquired MS/MS data were extracted and searched against Swiss-Prot databases using Spectrum Mill MS Proteomics Workbench software (Rev B.04.00.127; Agilent). Two databases were used, caprine (31,207 entry sequences and Homo sapiens [168,628 entry sequences), both updated on 3rd of December 2015). Mass tolerance of precursor and product ions was set to ±20 and ±50 ppm, respectively. Carbamidomethylation of cysteine was set as a fixed modification and oxidized methionine was set as a variable modification. Precursor mass shift was fixed between −18and 177 kDa to consider variable modifications such as the presence of sodium and potassium adducts. Identified proteins or peptides were validated using the same software based on the default settings, with the following inclusion criteria: protein score >20, peptide score >10, and scored peak intensity (% SPI) >70%. Proteins that shared at a minimum of one peptide were grouped together and the identified proteins were filtered to attain a false discovery rate (FDR) of <1% for the peptide-spectrum matches.
Data analysis and classification of MFGM proteins
Validation of MS/MS-based peptide and protein identifications from Spectrum Mill was carried out using Scaffold software (version 4.4.1; Proteome Software, Inc.) and protein identification was searched using X! Tandem algorithm (The GPM, thegpm.org; version CYCLONE; 2010.12.01.1). 20 Proteins sharing significant peptide evidence were grouped and annotated with gene ontology (GO) terms from NCBI (downloaded October 27, 2015).
Label-free relative quantification analysis
Mass Profiler Professional (MPP) software (version 13.0; Agilent) was used to analyze all data files of identified and validated proteins that were expressed in both human and caprine milk, as described by Thio et al. 21
Results
Representative patterns of human and caprine MFGM proteins isolated from pooled milk samples that were resolved in 10% SDS-PAGE gels are shown in Figure 1. Sixty micrograms of total MFGM protein from each pooled sample was loaded into the gel. Lane 1 demonstrates the abundantly expressed human MFGM protein bands, whereas caprine MFGM abundant protein bands are displayed in Lane 2. Prominent differences were detected between the SDS-PAGE profiles of abundant MFGM proteins sourced from human and caprine milk. Similar protein bands were seen at regions of around 50, 70, 70–100, and 140 kDa in both the MFGM protein samples. While 11 prominent bands appeared to have been detected in the human MFGM protein sample, the caprine MFGM protein sample demonstrated a total of nine well resolved bands, but with relatively higher content of low molecular weight proteins ranging between 25 and 35 kDa.
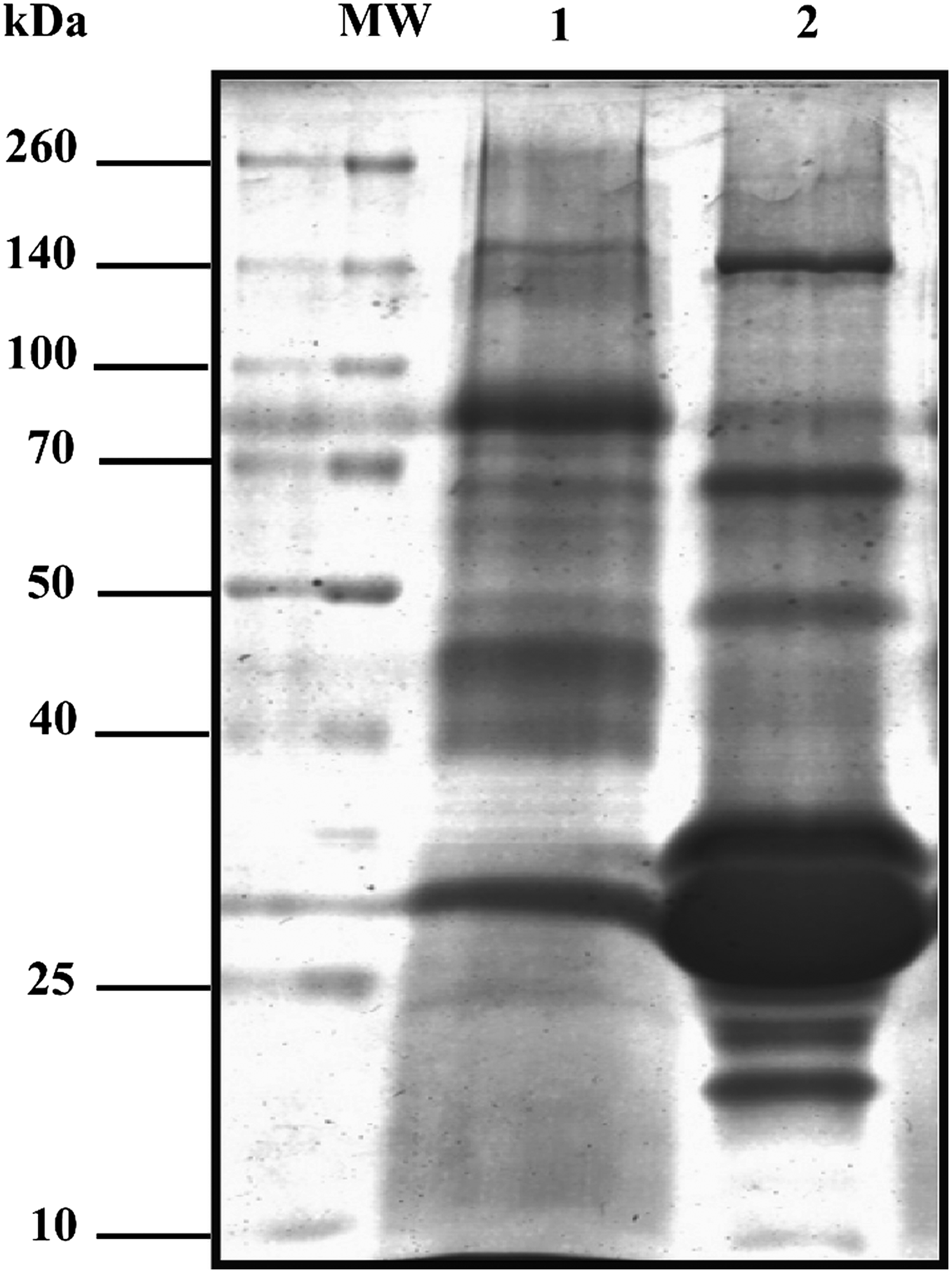
Representative SDS-PAGE patterns of MFGM proteins extracted from human and caprine milk. MFGM proteins were separated in 10% resolving gel and stained with Coomassie blue stain. MW denotes broad-range molecular weight marker. Lane 1 represents human MFGM proteins, while Lane 2 refers to caprine MFGM proteins. MFGM, milk fat globule membrane; SDS-PAGE, polyacrylamide gel electrophoresis in the presence of sodium dodecyl sulfate.
When the MFGM protein samples were subjected to trypsin digestion and analyzed by label-free relative quantification analysis using nano LC/MS Q-TOF and database search, a total of 128 abundantly expressed proteins were detected in the MFGM of humans (Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/bfm) and 42 were found in that of caprine (Supplementary Table S2). Among the predominant human MFGM proteins detected by our label-free quantification analysis were beta-casein, lysozyme, lactotransferrin, kappa-casein, carboxylic ester hydrolase, xanthine oxidase/dehydrogenase, mucin-1, EGF factor 8, butyrophilin subfamily 1 member A1, and apolipoprotein D (Table 1). However, caprine MFGM appeared to contain high abundance of EGF factor 8, alpha-S2-casein, isoform 2 of alpha-S1-casein, CSCN1, beta-casein, kappa-casein, serum albumin, glycosylation-dependent cell adhesion molecule 1, CD36, and beta-lactoglobulin (Table 1).
MFGM, milk fat globule membrane.
Among these proteins, only seven, namely alpha-S1-casein, beta-casein, butyrophilin subfamily1 member A1, fatty acid synthase, lactotransferrin, lipoprotein lipase, and EGF factor 8 protein (fragment), were commonly detected in both human and caprine MFGM (Table 2). In human milk, it was observed that the MFGM contains ∼6.8-fold higher abundance of lactotransferrin, 4.0-fold abundance of beta-casein, and 3.3-, 2.2-, and 1.6-fold abundance of lipoprotein lipase, fatty acid synthase, and butyrophilin subfamily 1 member A1, respectively (Table 3). On the contrary, caprine MFGM showed relatively higher abundance of EGF factor 8 and alpha-S1-casein, with the latter protein indicating more than 4.8-fold difference compared to that present in human MFGM.
(+), Denotes higher abundance in human milk relative to caprine milk; (
GO, gene ontology.
To provide further insight into their involvement in biological processes, the human MFGM abundant proteins identified were subsequently analyzed using the Scaffold software version 4.4. Figure 2 demonstrates the results of the GO analysis, in which the proteins were categorized into six biological processes of gene annotations. Forty-nine proteins were found to be involved in “metabolic process,” while 48 were involved in “response to stimulus.” This was followed by 27 proteins in “developmental process” and 23 proteins in “immune system process.” Thirteen of the proteins were apparently involved in “multicellular organismal process,” while “reproductive process” had three proteins involved (Supplementary Table S3).
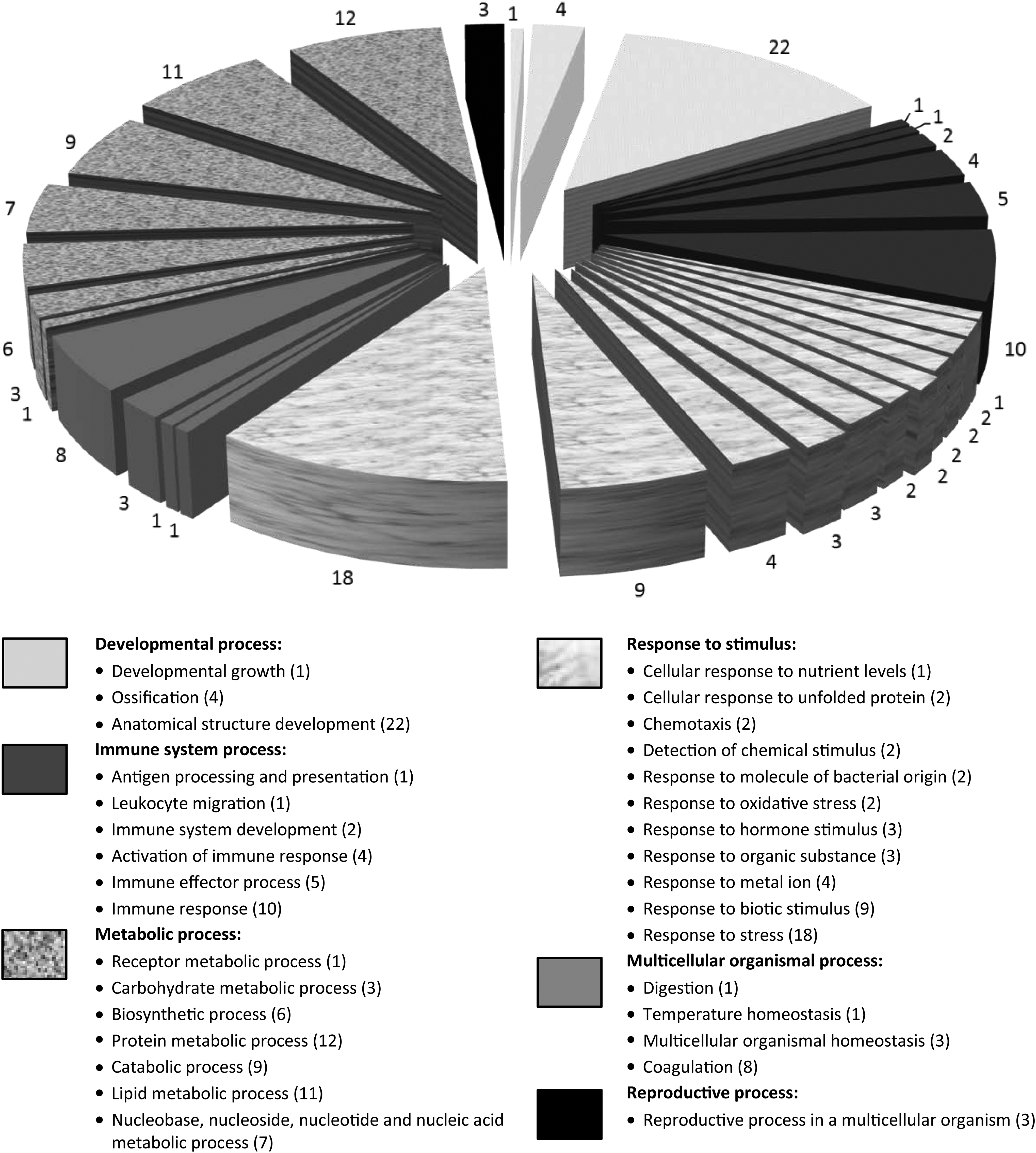
Gene ontology analysis of human MFGM proteins carried out using Scaffold software. A total of 128 proteins were analyzed and classified into 6 different gene annotations based on their biological roles. Color-coded keys represent gene annotation groups and the number of proteins involved in the process.
Discussion
In this study, we have compared the composition of selective abundantly expressed proteins in human and caprine MFGM using proteomics and software analyses. Our data showed that the human MFGM contained more abundantly expressed proteins compared with the caprine. While 128 abundant MFGM proteins were detected in human milk, only 42 were found in caprine milk. In addition, label-free quantitative protein analysis using LC/MS Q-TOF was carried out to determine the levels of abundances of these proteins. The 10 most abundant proteins measured in human MFGM were compared to those measured in caprine MFGM. Generally, the differences in these profiles suggested that MFGM contains important species-specific proteins expressed at different levels according to their functions that are specifically tailored to support the optimal physiological function and growth of human infants and goat's kid.
Our GO analysis revealed that human MFGM harbors a large number of abundantly expressed proteins involved in the metabolic and developmental processes. Altogether, there were 49 human MFGM proteins categorized under “metabolic process” and 27 were involved in “developmental process.” The term “metabolic process” is defined as the chemical reactions and pathways by which living organisms transform chemical substances, which include macromolecular processes such as “DNA repair and replication” and “protein synthesis and degradation.” Further analysis of the MFGM abundantly expressed proteins involved in “metabolic process” showed that they were mainly involved in protein and lipid metabolisms (Metabolic process in Supplementary Table S3), the former of which had 12 and the latter had 11 proteins, respectively. The gastrointestinal system of full-term infants is fully matured for digestive, protective, and absorptive functions. 22 However, certain functions such as the production of pancreatic enzymes are still underdeveloped, which require bioactive factors from colostrum and mature milk.22,23 The detection of many proteins and enzymes involved in protein and lipid metabolisms in this study supports the above observation, where they are crucial to compensate the infant's low digestive capacity.
In this study, it is interesting to note that there were only seven abundantly expressed proteins that were commonly present in both human and caprine MFGM. Even then, the levels of these proteins, which included alpha-S1-casein, beta-casein, butyrophilin subfamily1 member A1, fatty acid synthase, lactotransferrin, lipoprotein lipase, and EGF factor 8 protein (fragment), were actually different. In general, human MFGM contained substantially higher levels of lactotransferrin, beta-casein, lipoprotein lipase, fatty acid synthase, and butyrophilin subfamily1 member A1 compared to that in caprine MFGM. Lactotransferrin was among the predominantly expressed proteins in human MFGM, compared to caprine. As a member of transferrin family, this protein is responsible for binding and transferring of ferric ions. 24 It is involved in conferring the first line of defense in infants by preventing virus and bacteria from growing and multiplying in the intestinal mucosa through absorption of the iron that is required by the pathogens.25,26 The relatively lower level of lactotransferrin in caprine MFGM demonstrated in this study has also been previously reported, where the concentration of this protein in other ruminant milk is 10–100 times lower compared to human milk. 27 Apparently, the structure of caprine lactotransferrin is highly homologous to human lactotransferrin, and therefore, this protein has been suggested as a bioactive candidate for infant formula supplementation. 27
Caprine MFGM, on the other hand, appeared richer in alpha-S1-casein and EGF factor 8 (also known as lactadherin). These two proteins were also present in human MFGM, but at relatively lower abundances. While alpha-S1-casein is considered a minor protein in human MFGM, this study also demonstrated the presence of beta-casein, as the major casein constituent in human MFGM. Beta-casein, which is structurally unique among caseins due to the existence of multiple amino acids along its backbone, generates smaller casein phosphopeptides upon digestion to aid the absorption of calcium. 28
Not much is known about the specific function of EGF factor 8 protein in caprine, but it is generally involved in the maintenance of intestinal epithelial homeostasis and the promotion of mucosal healing. A study by Kvistgaard et al. showed that human EGF factor 8 inhibited rotavirus infection in vitro through protein–virus interaction. 29 In contrast, there are numerous studies on alpha-S1-casein, which is the most abundant casein in bovine and caprine milk. Caseins interact with calcium phosphate to form micellar complexes that provide sufficient calcium for the growth of ruminant neonates. 30 In humans, caseins have been thought to be one of the sources of amino acids, as well as improving the infant's intestinal absorption of trace elements and calcium. 31 Alpha-S1-casein had been shown to be expressed in human monocytes and exhibited an immunomodulatory role. 31 However, this subunit of casein is known to be a major allergen in children with CMPA. 32 The amino acid sequence of this protein in bovine milk is markedly different from the sequence of the same protein detected in caprine milk, and according to Gupta et al., a protein's primary structure determines its chemical and biological attributes.32,33 In view of these differences, it has been suggested that caprine milk could serve as a hypoallergenic alternative formula to infants with CMPA, when breastfeeding was not possible. 32
However, it is clear that until substantial evidences are obtained from both in vitro and clinical studies, the use of caprine milk in infants with CMPA cannot be considered fully safe. 34 European Food Safety Authority's Panel on Dietetic Products, Nutrition and Allergies had published their opinion regarding the substitution of bovine milk-based infant formula with caprine milk-based infant formula for infants with CMPA. 34 The Panel noted that it is possible for caprine milk protein to also induce allergic reaction, but the data to prove this notion are still inadequate. 34
Out of 128 proteins detected in human MFGM, 7 proteins, or 5.8%, were also detected in caprine MFGM. The percentage may be small; nonetheless, these bioactive proteins also contributed to important roles in infant growth. Human milk has always been considered superior than those of ruminants, thus retaining that the beneficial MFGM in caprine milk-based formula may bring health and nutritional value to the infant. Nevertheless, it should be kept in mind that the information, production, and use of non-bovine milk-based formula are not as comprehensive as bovine milk-based formula, and therefore, there is a need for more research to fill in these gaps of knowledge.
Conclusions
When taken together, this study showed that human MFGM was different from caprine MFGM, where the human MFGM appeared far more superior as it contained 128 abundantly expressed proteins that were generally not detected in caprine MFGM. The minor similarity in the abundantly expressed bioactive protein composition of caprine MFGM that was detected in this study further suggests that it was still nutritionally beneficial, and therefore should be included when caprine milk-based formula is used as an alternative when breastfeeding or human milk administration is not possible or solely adequate for consumption.
Footnotes
Acknowledgments
We acknowledge the assistance of Puan Nor Kamariah Mohamad Alwi from ![]() and breastfeeding mothers who voluntarily contributed their precious milk toward this study. This work was supported from the funding of Fundamental Research Grant (FP 092/2010A) from the Ministry of Higher Education, Malaysia, and Postgraduate Research Grant (PG 082/2012B) granted by University of Malaya, Malaysia.
and breastfeeding mothers who voluntarily contributed their precious milk toward this study. This work was supported from the funding of Fundamental Research Grant (FP 092/2010A) from the Ministry of Higher Education, Malaysia, and Postgraduate Research Grant (PG 082/2012B) granted by University of Malaya, Malaysia.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.