
Abstract
Abstract
Objective:
Human milk administration in the early peritransplant period would lower intestinal inflammation after bone marrow transplant (BMT).
Materials and Methods:
Children 0–5 years undergoing BMT received either a ready-to-feed human milk preparation designed for these children (Prolacta Bioscience, Duarte, CA) or standard formula. Babies breastfeeding at the time of BMT were also enrolled on the human milk arm. Human milk was administered from day −3 until day +14 after BMT. Metagenomic shotgun sequencing and metabolomics of stool, plasma cytokines, and regenerating islet-derived 3α (REG3α) levels were measured at enrollment and day +14. Human leukocyte antigen-DR isotype (HLA-DR), CD38, and CD69 expression on T cells were evaluated at day +21.
Results:
Forty-six children were enrolled, 32 received human milk (donor milk n = 23, breastfeeding babies n = 9), and 14 were controls who received standard feeds supervised by a BMT dietician. Twenty-four patients received at least 60% of goal human milk and were evaluable. Plasma interleukin (IL)-8 (p = 0.04), IL-10 (p = 0.02), and REG3α (p = 0.03) were decreased in the human milk cohort. Peripheral blood CD69+ CD8+ T cells were higher in controls (p = 0.01). Species abundance of Adenovirus (p = 0.00034), Escherichia coli (p = 0.0017), Cryptosporidium parvum (p = 0.0006), Dialister invisus (p = 0.01), and Pseudomonas aeruginosa (p = 0.05) from stool was higher in controls. Stool alanine, tyrosine, methionine, and the ratio of fecal alanine to choline and phosphocholine were higher in controls (p < 0.05). No difference was observed in stool propionate and butyrate levels as measures of short-chain fatty acids between the two cohorts.
Conclusions:
Administration of human milk resulted in decreased markers of intestinal inflammation and could be a valuable adjunct for patients after BMT.
Introduction
Intestinal microbiota maintains gut homeostasis by stimulating cytokines and chemokine secretion from intestinal epithelial cells. 1 Mucosal injury caused by high-dose chemotherapy or radiation before bone marrow transplant (BMT) sets off a cascade of inflammatory cytokines such as interferon gamma (IFN-γ) and interleukin (IL)‐6. 2 These cytokines modify intestinal microbiota composition resulting in dysbiosis 3 and upregulate the expression of major histocompatibility complex and immune costimulatory molecules, initiating the pathophysiology of acute graft versus host disease (GVHD). 4 Increased intestinal permeability due to mucosal damage allows translocation of bacteria across the intestinal mucosa resulting in bloodstream infections (BSIs).5,6 Infections with specific pathogens early after BMT have been shown to lead to a higher incidence of acute GVHD. 6 Acute GVHD is the most significant complication of BMT and carries a mortality rate of 30–50%.7,8 BSIs after BMT are immediately life threatening and are a significant health care burden. 6 Intestinal dysbiosis is associated with both acute GVHD pathogenesis and incidence of BSI.9–11 If intestinal homeostasis could be restored or maintained after BMT, especially in the period of neutropenia lasting 2 to 3 weeks after stem cell infusion, we can potentially prevent acute GVHD and BSI.
Human milk oligosaccharides (HMOs) are nondigestible carbohydrates in breast milk 12 and the most abundant bioactive constituent of human milk. 12 Commensal bacterial metabolize HMO and produce short-chain fatty acids (SCFAs), which are the main source of energy for epithelial cells lining the colon creating a favorable environment for the growth of beneficial bacteria.12–15 Human milk also contains anti-inflammatory enzymes, anti-inflammatory cytokines, antiproteases, lactoferrin, and secretory immunoglobulin A, which stimulate maturation of the neonate's intestinal defenses.16,17 In breastfed preterm infants, breastfeeding has been associated with a reduced incidence of necrotizing enterocolitis, allergic diseases, and autoimmune diseases, suggesting that breast milk may modulate the development of the immune system.18–22
We hypothesized that children receiving human milk would have lower markers of intestinal inflammation and injury and a more diverse microbiome after a BMT. We tested our hypothesis by performing a clinical trial of a human milk-based product by administration to children after a BMT and observed these children for development of acute GVHD and BSI.
Materials and Methods
Clinical trial
We conducted a prospective clinical trial at Cincinnati Children's Hospital Medical Center. This trial was registered on clinicaltrials.gov (NCT02470104). The original intent was to conduct an unblinded randomized clinical trial and enroll 42 patients. We perform ∼100 BMTs at our center every year. About 20% of these patients are ≤5 years of age. We wanted to complete our pilot study in 2 years and based on the amount of donor human milk available to us, we estimated that we would be able to enroll 42 patients successfully in 2 years We intended to enroll and randomize patients 2:1 to receive either donor human milk preparation formulated specially for the study (Prolacta Bioscience, Duarte, CA) or standard feeding with formula. Some of these 42 children being enrolled on study were receiving mothers' own milk at the time of the study and were enrolled on the human milk arm without any randomization. Controls received standard cow's milk-based nutritional supplementation through enteral tubes, if indicated. Babies breastfeeding at the time of BMT were assigned without randomization to the human milk arm. The number of breastfeeding babies to be enrolled on the human milk arm without randomization was not predetermined at study onset. No control was receiving human milk immediately before BMT (Fig. 1).
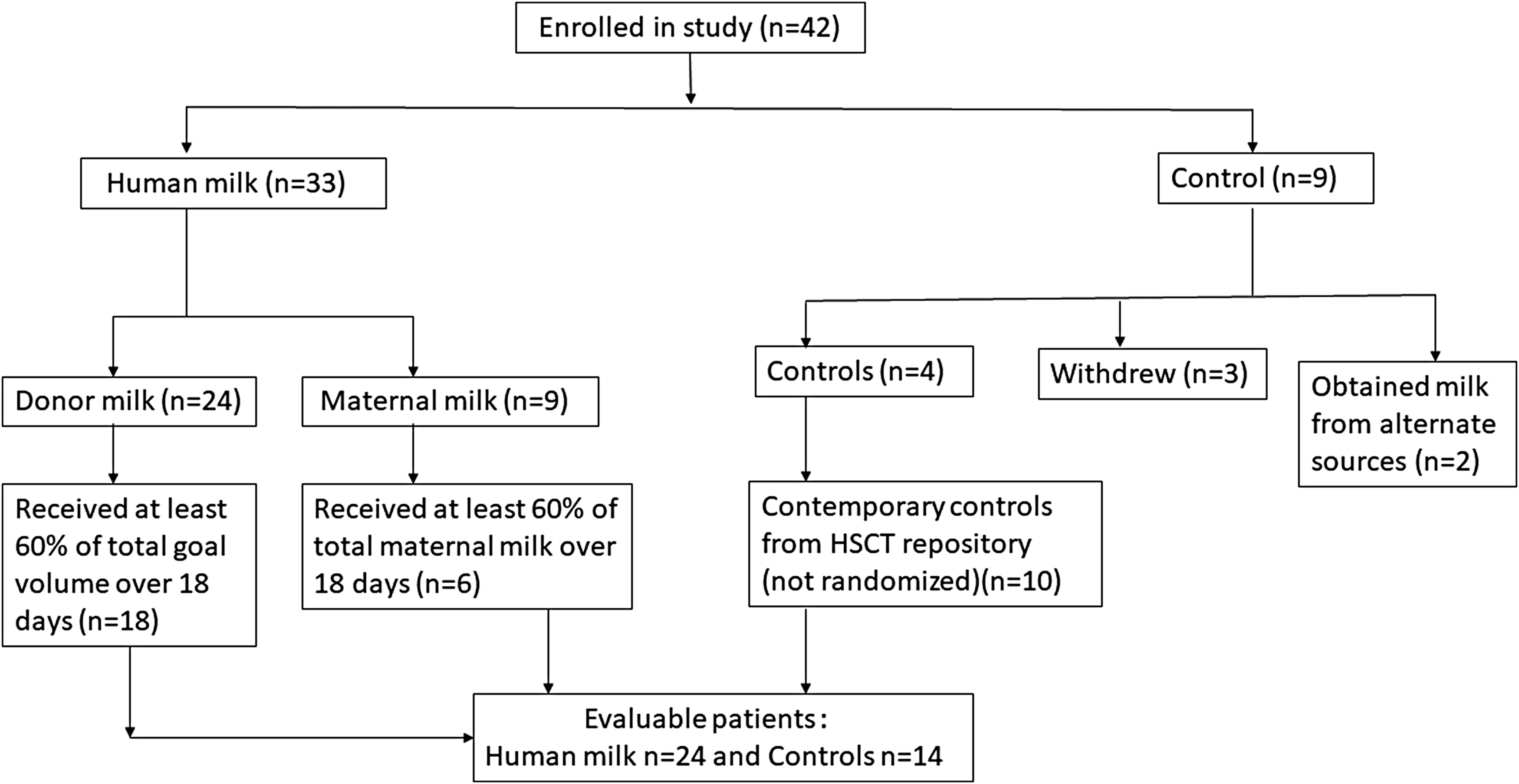
Trial profile.
An expert BMT dietician supervised both arms. Human milk product was started on day −3 and continued until day +14 after BMT, the period of maximal intestinal inflammation. All patients, regardless of randomization, also received parenteral nutrition in addition to enteral nutrition. We determined that human milk should account for at least 40% of calories based on early work done in children where we observed tolerability of this volume of feeds around BMT and also based on the fact that human milk should not be the sole source of calories in children >6 months of age. In children who were receiving donor milk product and who were receiving expressed maternal breast milk, study protocol specified that “adequate” exposure to human milk to observe biologic effects and be included in analysis would be at least 60% of goal human milk volume intake. In infants who were breastfeeding at the time of BMT, goal breastfeeding was defined as feeding at least five times a day, ∼10 minutes per side. An “adequate” exposure of human milk in breastfeeding babies was defined as receiving 60% of goal breastfeeding.
Problems with randomization
We enrolled 33 patients on the human milk arm and 9 controls. Three subjects withdrew from the study when assigned to the control arm. Two controls obtained human milk surreptitiously from alternate sources informing study staff 1 year after study completion. Four controls were eventually evaluable on the planned randomized study. We then altered our randomized trial study design to a pilot clinical trial with historic controls as a comparison group, enrolling 10 nonrandomized contemporary controls who would have met study eligibility to augment the control arm and had available stored samples for analysis (Fig. 1). These 10 contemporary nonrandomized controls were enrolled on our BMT repository in the year before initiation of the study and would have met enrollment criteria of the study and had stool and plasma samples collected at the desired time points.
The study was approved by the Institutional Review Board of Cincinnati Children's Hospital Medical Center (CCHMC), and written informed consent was obtained from parents of participating children.
Prolacta Bioscience, Inc.® manufactured the donor human milk product specially for this study. The product had 2.15 g protein and 97.7 Kcal/dL and was rigorously screened to eliminate pathogens, especially Bacillus cereus, and pasteurized through two heat treatments.
Incidences of acute GVHD and BSI and cytomegalovirus (CMV), adenovirus, and Epstein–Barr virus (EBV) DNA loads were documented until day +100. Acute GVHD was graded based on consensus criteria.
Analyses
We performed an “as-treated” analysis of our data to test the biologic effect of our intervention in addition to an intention-to-treat analysis of primary study end points.
Pro- and anti-inflammatory cytokines and regenerating islet-derived 3α (REG3α) were measured at enrollment and day +14. Markers of T cell activation CD69, CD38, and Human leukocyte antigen-DR isotype (HLA-DR) were measured at day +21 after BMT, stool microbiome and metabolomics were evaluated at baseline and day +14 after BMT.
Plasma cytokine and REG3α analyses
Plasma cytokines were measured on a Luminex 200 Instrument (Luminex Corporation, Austin, TX) according to the manufacturer's instructions with the Human Cytokine Magnetic 10-Plex Panel (Invitrogen, Carlsbad, CA). Cytokines assessed included IL-1β, IL-2, IL-4, IL-5, IL-6, IL-8, IL-10, IFN-γ, tumor necrosis factor-α, and granulocyte macrophage-colony stimulating factor (GM-CSF). We measured IL-8 on day +7 on all patients by ELISA (enzyme-linked immunosorbent assay) (R&D Systems, Inc., Minneapolis, MN) per manufactures instructions. REG3α levels from plasma were measured at enrollment and day +14 after BMT by ELISA (R&D Systems, Inc.) per manufacturer's instructions. We compared the ratio of day +14: baseline and day +7: baseline of all cytokines between the two cohorts by Mann–Whitney U test. If patients experienced clinical events known to be associated with cytokine elevations (e.g., engraftment syndrome or sepsis) on day +7 or day +14, we a priori excluded them from cytokine analyses. We also compared the ratio of day +14: baseline REG3α between the two cohorts.
CD69, HLA-DR, and CD38 expression on T cells
Cryopreserved peripheral blood mononuclear cells were evaluated. Cells were incubated with fluorochrome-conjugated monoclonal antibodies directed against CD3 and CD69 (Biolegend, San Diego, CA), CD8 and HLA-DR (BD Biosciences, San Jose, CA), CD38 (eBioscience, San Diego, CA), and analyzed on a BD FACS Canto II flow cytometer (BD Biosciences). Patients were evaluable for analyses if they had at least 200 CD4+ and CD8+ T cell events.
Microbiome analyses
Metagenomic shotgun sequencing was performed for stool samples collected at enrollment, day 0, day +7, and day +14 after BMT. DNA was extracted from stool samples using the PowerFecal kit from Qiagen per kit instructions. DNA concentrations were measured using Qubit. Library generation was performed using Illumina Nextera XT adapters and metagenomic shotgun sequencing was performed on a NextSeq500 platform in the Precision Metagenomic Core Research Laboratory (PML) at CCHMC. Raw sequencing data underwent quality control measures followed by read alignment to species using a custom-built and validated pipeline by the PML. Statistical analyses of diversity, principal component analysis, and partial least squared discriminant analysis (PLS-DA) were all performed using R software. Alternative multiple response permutation procedures (MRPP) was used to compare overall species abundance composition between groups. Pairwise comparison between the two cohorts was performed by the Wilcoxon rank sum test. Stool samples at all time points between the two cohorts were grouped for comparison.
Metabolomics
Stool samples were lyophilized and weighed (80 ± 3 mg) into 2 mL bead tubes containing 2.8 mm ceramic beads (VWR International LLC, Radnor PA) before sample processing. Owing to limited volumes of sample collected, 40 ± 2 mg sample size was used for four of the samples. Each sample preparation was performed in a batch of 12 to 13 test samples, a blank, and a QC control material. Pooled dried fecal control material prepared in-house was used in this study to ensure reproducibility and accuracy of results. On the day of processing, 1 mL of cold phosphate buffered saline was added to the sample, homogenized for 30 seconds at 5,000 rpm using Minilys (Berten Tech.). Samples were then transferred into a 2 mL microcentrifuge tube and centrifuged at 10,000 × gn at 4°C for 10 minutes. Supernatants (400 μL) were then filtered at 12,000 × gn for 60–90 minutes at 4°C using prewashed 3 kDa spin filters (NANOSEP 3K; Pall Life Sciences). Nuclear magnetic resonance buffer containing 100 mM phosphate buffer in D2O, pH 7.3, and 1.0 mM TMSP (3-trimethylsilyl 2,2,3,3-d4 propionate) was added to 198 μL of fecal filtrate. The final sample volume was 220 μL and final TMSP concentration in each sample was 0.1 mM.
Experiments were conducted using 200 μL samples in 103.5 × 3 mm NMR tubes (Bruker). One-dimensional (1D) 1 H NMR spectra were acquired on a Bruker Avance II 600 MHz spectrometer. All data were collected at a calibrated temperature of 298 K using the noesygppr1d pulse sequence in the Bruker pulse sequence library. All free induction decays were subjected to an exponential line broadening of 0.3 Hz. On Fourier transformation, each spectrum was manually phased, baseline corrected, and referenced to the internal standard TMSP at 0.0 ppm using Topspin 3.5 software (Bruker Analytik, Rheinstetten, Germany). For a representative sample, two-dimensional (2D) 1 H– 13 C heteronuclear single quantum coherence (HSQC) data were collected for metabolites assignment. Chemical shifts were assigned to metabolites based on 1D 1 H, 2D TOCSY and HSQC NMR experiments with reference spectra found in Chenomx® NMR Suite profiling software (Chenomx, Inc., version 8.1). Concentrations of metabolites present in the filtered fecal water were calculated using Chenomx software based on the internal standard, TMSP. Thirty-four metabolites were quantified (μM) and concentrations were normalized to the original dry tissue weight (mg) before the statistical analysis.
Results
Forty-two children were prospectively enrolled. Twenty-four nonbreastfeeding children were randomized to receive the donor human milk-based product and nine were randomized to receive standard nutrition per routine clinical care. Four of nine randomized controls completed study protocol (Fig. 1). Nine babies breastfeeding at the time of BMT were allocated to the human milk arm without randomization (Fig. 1). Two babies received expressed breast milk, three were breastfed, whereas four babies received a combination of expressed breast milk and breastfeeding. None of the nine breastfed infants at the time of BMT required any donor human milk supplementation during the trial. Ten nonrandomized contemporary controls were added to the control arm due to the unexpected attrition of patients on the control arm. No additional oral nutritional supplementation was administered to patients on either arm. All patients (human milk, randomized controls, and nonrandomized contemporary controls) received antiviral, antifungal, and Pneumocystis jirovecii prophylaxis as part of standard of care. Cefepime was the antibiotic of choice for febrile neutropenia.
Twenty-four patients in the human milk arm (18 who received the human milk preparation and six who received maternal milk) received at least 60% of total goal feeds for the study period and were evaluable for “as-treated” analyses (Fig. 1). All controls received at least 60% of their goal standard formula from day −3 to day +14.
Supplementary Table S1 shows transplant characteristics. Median age was similar between children who received human milk (1.25 years; range 0.3–4.9 years) and controls (1.36 years; range 0.5–4.9 years).
“As-treated” and “augmented controls” analyses to assess biologic effect of human milk
Five patients in the human milk arm were excluded from cytokine analysis as they experienced gram-negative sepsis (n = 1) or engraftment syndrome (n = 4) at day +14. We measured day +7 IL-8 levels in all patients because IL-8 is influenced by neutrophil engraftment, typically occurring around day +14, confounding levels at that day.
Ratios of day +14 to baseline REG3α (p = 0.03), IL-6 (p = 0.04), IFN-γ (p = 0.04), and IL-10 (p = 0.004) were significantly elevated in the randomized controls compared with the human milk arm. The ratio of day +7 to baseline IL-8 (p = 0.04) was elevated in the randomized controls compared with the human milk arm (Fig. 2A–E). After augmentation of our 4 randomized controls with 10 contemporary nonrandomized controls, the ratio of day +14 to baseline IL-10 (p = 0.02) was significantly elevated in controls compared with the human milk arm (Fig. 2G) as was the ratio of day +14 to baseline REG3α (p = 0.03) (Fig. 2H) and day +7: baseline IL-8 (p = 0.04) (Fig. 2I). We were unable to perform IFN-γ analyses on the 10 contemporary nonrandomized controls due to exhaustion of available samples on our repository.

Ratio of day +14 IL-6, IL-10, IFN-γ, and REG3α and day +7 IL-8 to baseline in human milk cohort compared with randomized controls or randomized controls + contemporary nonrandomized controls.
Eight human milk recipients and 13 controls had at least 200 CD4+ and CD8+ T cell events. Median percentage expression of CD69 (an early marker of activation) on CD8+ T cells in the milk recipients was 3% (range 1–55%) compared with 23% (range 1–86%) in controls (p = 0.01). Median expression of CD69 on CD4+ T cells and CD38 and HLA-DR (later markers of T cell activation) on all T cells was not different (Fig. 3A).
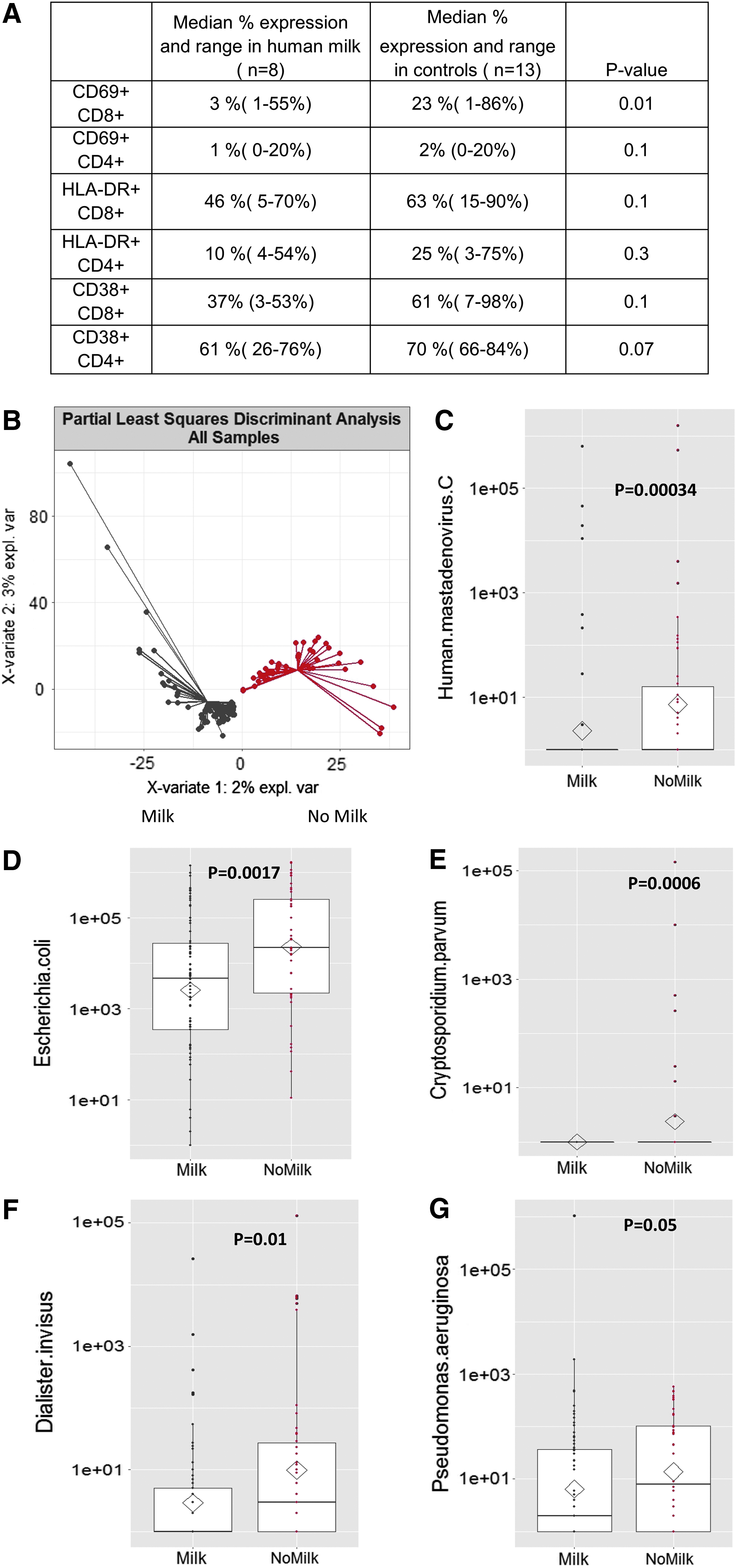
We compared stool samples between 24 human milk patients and 14 controls (4 randomized controls and 10 nonrandomized contemporary controls). Principal co-ordinate analysis comparing intestinal microbiome species between the cohorts by MRPP showed differences between the two cohorts (p = 0.033). Supervised analysis (PLS-DA) further resolved the microbiomes between the two groups (Fig. 3B). Feature selection of species composition revealed controls had a higher species abundance of Mastadenovirus C. (p = 0.00034), Escherichia coli (p = 0.0017), Cryptosporidium parvum (p = 0.0006), Dialister inversis (p = 0.01), and Pseudomonas aeurginosa (p = 0.05) (Fig. 3C–F).
Greater intestinal inflammation should cause more intestinal epithelial shedding in stool. 23 We observed higher human DNA in control stool controls compared with milk recipients (p = 0.0067).
Twenty stool samples of human milk recipients and five controls (randomized + contemporary nonrandomized controls) were evaluated at baseline and day +14 for metabolomic analyses. Levels of fecal alanine, methionine, and tyrosine were lower in human milk patients (Fig. 4A–C). Stool propionate at day +14 (Fig. 4D) and stool butyrate, as measures of SCFAs, were not different between the two cohorts. Ratios of alanine to phosphocholine and alanine to choline were significantly higher in controls (p < 0.01) (Fig. 4E, F). All other metabolites tested were not significantly different between the two cohorts (data not shown).
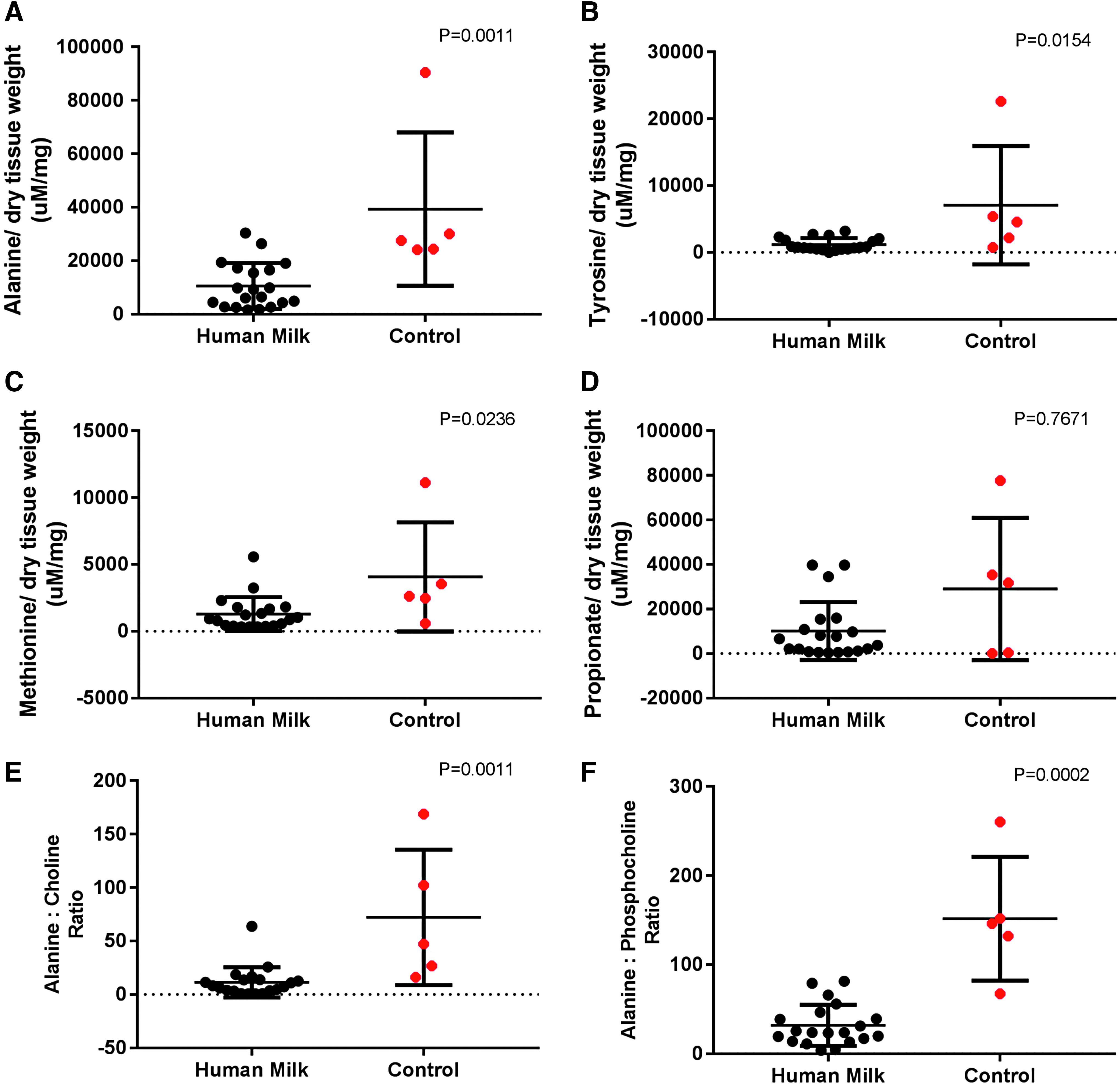
Stool metabolomic analyses of amino acids at day +14.
CMV, EBV, and adenovirus viremia by day +100 after BMT were not significantly increased in children who received human milk compared with controls (Fig. 5A). There were no B. cereus infections or other infections attributable to the milk product.

Four of 23 children within the human milk arm who received an allogeneic BMT had grades I–IV acute GVHD, whereas 3 of 4 randomized controls had grade I–IV GVHD (p = 0.04). Two of 23 human milk recipients had grade II–IV GVHD compared with 2 of 4 randomized controls with grade II–IV GVHD (p = 0.09). Two of 23 human milk recipients had gastrointestinal (GI) GVHD compared with 2 of 4 randomized controls (p = 0.14). When we added 10 contemporary nonrandomized controls to the analysis, 6 of 14 controls had acute grade I–IV GVHD (p = 0.13) and 4/14 had grade II–IV GVHD (p = 0.17). Three of six patients with acute GVHD had intestinal involvement (p = 0.65) (Fig. 5B).
The incidence of BSI was 3 per 100 central venous catheter days in the human milk arm compared with 9 per 100 central venous catheter days (p = 0.06) in the randomized control arm and 11 per 100 central venous catheter days (p = 0.2) in the randomized control plus contemporary nonrandomized control arm.
We also performed an intent-to-treat analysis of primary end points of the study on the original cohort of 42 patients (33 randomized to receive human milk and 9 randomized controls). Five of 9 controls versus 5/32 human milk recipients had acute GVHD (p = 0.03), whereas 3/9 controls versus 3/32 human milk recipients had GI GVHD (p = 0.11). Four of 9 controls versus 7/33 human milk recipients had a BSI by day +100 (p = 0.19).
Discussion
We show that administration of human milk to children is feasible after BMT. IL-8, IFN-γ (proinflammatory), and IL-10 (anti-inflammatory) levels were reduced 14 days after BMT in children who received human milk. IL-10 is an anti-inflammatory cytokine 24 that typically increases in response to a proinflammatory cytokine surge after BMT to maintain homeostasis. Plasma REG3α levels, a marker of intestinal injury 25 and GI GVHD, 26 were elevated in controls suggesting that human milk protected from intestinal injury. Similarly, human DNA was increased in the stool of controls. Expression of CD69, an early T cell activation marker 27 implicated in acute GVHD pathogenesis, was higher on CD8+ T cells in controls compared with children who received human milk, supporting inflammation in controls. CD38 or HLA-DR on CD4+ or CD8+ T cells were not significantly upregulated due to the early time point (day +21) of assessment in our patient population. Activation of CD38 and HLA-DR31 typically occurs at later time points.
Mastadenovirus, a common pathogen after BMT, was reduced in stool of milk recipients compared with controls, supporting a role for human milk in modifying the intestinal microbiome.28,29 Our observation of lower detection of E. coli and Pseudomonas aeruginosa from the stool of human milk recipients, bacteria associated with acute GVHD, supports our trend toward lower incidences of BSI and acute GVHD.4,9,30,31 We did not observe a difference in bacterial diversity between the two cohorts but believe that intestinal inflammation is impacted by mechanisms beyond bacterial diversity, such as impact of intestinal viruses, fungi, and bacteriophages. Since analyses of the human microbiome are challenged by false discovery related to multiple comparisons, we used a systems biology approach by coupling microbiome analyses with NMR to examine the corresponding metabolic features of the microbiome. In addition, we have previously shown a progressive decline in stool butyrate and propionate in children with imminent acute GVHD, 32 increasing our enthusiasm for performing metabolomic analyses in this study. Our data show lower levels of fecal choline, as have been observed in patients with active inflammatory bowel disease compared with controls.33–36 Our finding of higher alanine:choline ratios in controls suggests that lower choline levels in controls could likely be due to microbial shifts leading to increased conversion to trimethylamine, which in turn contributes to an inflammatory milieu in the intestine. We did not observe differences in SCFAs as measured by stool butyrate and propionate between our two cohorts, likely due to the low abundance of these in the stool in children at day +14 after BMT (unpublished observation, Romick-Rosendale laboratory).
Pathogens observed to be conveyed by breastfeeding to the BMT population, for example, CMV, or through pasteurized donor milk, for example, B. cereus, were not higher in children who received human milk compared with controls.
Components in human milk 37 are well known to impact the host immune system. HMOs influence the intestinal microbiome and promote growth of healthy bacteria in the intestine. 16 A healthy intestinal microbiome after BMT has been associated with lower incidences of acute GVHD and BSI in several studies.3,4 Lactoferrin promotes growth of commensal bacteria and reduces inflammatory cytokine production,38,39 which could explain lower IFN-γ levels in human milk recipients. Lastly, human milk itself contains anti-inflammatory cytokines40,41 such as IL-10 and transforming growth factor-β that act directly on enterocytes and reduce IL-8 production.
Our study has several limitations. The study was not blinded that led to failure of study randomization. We recognize the obvious criticism of selection bias of contemporary nonrandomized controls but minimized this by selecting consecutive controls that met trial eligibility criteria. We combined donor human milk and breastfeeding babies in a single group for comparison when there could be important differences between these two groups in terms of mechanism of how human milk reduced intestinal inflammation. Given the human biodiversity as well as the biodiversity of human milk and many potential cofounding variables, larger studies are needed before human milk or human milk products can be used for the benefit of BMT recipients in infants and young children.
In summary, our pilot study shows that human milk administration after BMT is feasible and safe in young children, and might reduce intestinal inflammation and injury, supporting further clinical trials.
Footnotes
Acknowledgments
The authors thank all the clinical staff at Cincinnati Children's Hospital and all patients and families. Donor human milk product used in this study was kindly provided by Prolacta Bioscience. Prolacta Bioscience had no role in data acquisition, interpretation of results, or decision to publish this study. Results of this study were presented at the American Society of Bone Marrow Transplantation Annual Meeting, 2018.
Ethics Committee Approval
This study was approved by the Cincinnati Children's Hospital Medical Center Institutional Review Board.
Disclosure Statement
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.