
Abstract
Abstract
Background:
Some strains of Staphylococcus aureus can produce heat-stable enterotoxins that have been associated with gastritis and potentially necrotizing enterocolitis in preterm infants.
Objectives/Hypothesis:
To assess the impact of different storage temperatures on S. aureus growth and enterotoxin production in raw, Holder-pasteurized (HP) and ultraviolet-C (UV-C)-treated donated human milk (DHM).
Materials and Methods:
The milk samples from individual donors were pooled and divided into four equal portions. One portion was HP, the second was UV-C treated, the third was not treated, and the fourth was UV-C treated after being spiked with S. aureus. All samples were incubated at 37°C (18 hours) and 4°C (14 days). Bacterial colony count, enterotoxin A and B, and immune proteins were quantified.
Results:
At 37°C, the colony count increased in HP DHM and decreased in raw and UV-C-treated DHM. At 4°C, colony counts in HP DHM reduced and were not detected in raw and UV-C-treated DHM from day 8 of incubation. No bacteria were detected in samples that were inoculated before UV-C treatment. Enterotoxin A was only detected in HP-DHM at 37°C from the 9th hour onward. Enterotoxin B was detected in one sample at the 15th hour. Immune protein concentrations were similar in raw and UV-C DHM, and were reduced in the HP DHM.
Conclusion:
UV-C-treated milk reduces S. aureus growth with similar kinetics to raw milk making it a promising emerging technique to eliminate bacteria while retaining essential immune proteins in DHM.
Introduction
Mothers' own milk is the preferred nutrition for preterm infants due to its rich content of nutritional and immune proteins that protect them against infections as well as developing intestinal adaptation and maturation. 1 Depending on the energy requirements of the infant, human milk (HM) is often fortified to facilitate growth. When mothers do not have sufficient milk, donor human milk (DHM) is considered the next best alternative for preterm infants. 2 However, DHM may contain potential pathogens that increase the risk of harm to preterm infants. Many milk banks reduce these risks by heat treating DHM to minimize the risk of transmitting infections from DHM to preterm infants.
Many milk banks are currently using Holder pasteurization to eliminate bacteria in DHM, which is performed by heating the milk for 30 minutes at 62.5°C.3,4 This mode of pasteurization is capable of eliminating most bacteria from DHM. However, it leads to a significant loss of the immune protective components in DHM.5,6 Furthermore, Holder pasteurization is incapable of eliminating heat-stable enterotoxins that are produced by some strains of Staphylococcus aureus. 7
S. aureus in one of the most common potential pathogenic bacterial species found in DHM.8–11 Certain strains of S. aureus produce heat-stable enterotoxin, such as Staphylococcal enterotoxin A (SEA) and Staphylococcal enterotoxin B (SEB), and these are considered the most common enterotoxins capable of causing food poisoning and gastroenteritis. 12
Studies have shown that S. aureus does not produce a detectable enterotoxin in milk until it reaches a density of >10 6 colony-forming units per mL (CFU/mL).13,14 In addition, we have previously shown that, Holder-pasteurized (HP) DHM inoculated with S. aureus posttreatment produces enterotoxin only after 9 hours of incubation at 37°C, whereas no enterotoxin is produced in raw milk. 15 These results highlight the importance of the role of immune components in raw DHM in impeding proliferation of S. aureus and subsequently production of enterotoxin. Typically, there is a significant reduction in immune proteins in HP DHM compared with both raw milk and ultraviolet (UV)-C-treated DHM. 16
UV radiation is a nonthermal method of pasteurization. 17 UV-C radiation has a wavelength between 200 and 280 nm 18 and is effective at eliminating bacteria, viruses, and fungi due to the germicidal affect between 250 and 260 nm, with the maximum effect occurring at a wavelength of 254 nm. 18 The germicidal effect is due to cellular damage caused by alteration of the DNA structure. 19 The pyrimidine and purine bases of DNA absorb the UV-C energy, which leads photoproducts such as pyrimidine hydrates and dimers as well as protein crosslinkage. 20 This cellular inactivation and malfunction results in the inhibition of bacterial proliferation and cell death.
UV-C is a treatment commonly used by the food industry. 21 However, the effectiveness of using UV-C to treat milk has been limited due to the opaque nature of milk; its absorption coefficient is high (300 cm−1) compared with water (0.1 cm−1) 20 or pineapple juice, which has the highest absorption coefficient of all juices of 73 cm−1. 22 Juices with an opaque nature need to be fully exposed to the UV-C by increasing the flow rate of the juice inside the UV-C reactor to make sure that all parts of the liquid are exposed to the irradiation resulting in a higher bacterial inactivation rate. 23 Furthermore, the HM absorption coefficient also increases with an increasing total solids concentration, which is variable in HM, further limiting UV-C penetration. 24 UV-C has been used commercially to treat bovine milk and it was successful in treating milk by rapid turbulent flow to expose all parts of the milk to the UV-C. 25 Thus, treating HM with UV-C requires special equipment to allow adequate application of UV light to a volume of milk. This can be achieved using equipment producing a low vortex in the milk forming a thin film on a transparent surface allowing UV-C exposure to all parts of the milk. As such UV-C has been used to successfully eliminate bacteria in HM.24,26
For UV-C to be seriously considered as an alternative treatment option for Human Milk Banks, it is important to determine whether UV-C treatment of HM induces increased bacterial toxin production during the inactivation process, or alternatively, whether it inhibits the production of bacterial toxin by inhibiting the bacterial proliferation in HM.
The aim of this study was to investigate the ability of S. aureus to produce enterotoxin in DHM treated with UV-C compared with HP DHM and raw milk. We hypothesize that UV-C-treated DHM will inhibit bacterial proliferation due to the higher retention of HM bactericidal immune proteins and, therefore, enterotoxin production will be minimal.
Materials and Methods
Sample source
Milk samples were obtained from four donors who donated milk to the Perron Rotary Express Milk Bank (PREM Bank), King Edward Memorial Hospital for Women, Subiaco, Western Australia. All donors had given prior consent for their milk to be used in research (Western Australian Health Human Research Ethics Committee; 2014127EW).
The milk samples from individual donors were pooled and divided into four equal portions. One portion was HP, the second was UV-C treated, the third was not treated, and the fourth was UV-C treated after being spiked with S. aureus.
Holder pasteurization
Milk samples (400 mL; n = 4) were pasteurized using the PREM Bank pasteurizer (Sterifeed S90; Medicare Colgate Ltd., United Kingdom) according to the PREM Milk Bank guidelines. The milk reached a temperature of 63°C ± 0.5°C for 30 (−0/+7) minutes and the temperature profile was recorded using a thermocouple (NATA) to confirm the heating profile.
UV treatment
Each pooled milk sample (400 mL; n = 4) was transferred into a 600 mL PYREX glass beaker. A germicidal UV-C lamp (GPH287T5L; Infralight Pty Ltd, Helensburgh, NSW, Australia) was placed diagonally in the beaker. The uncovered part of the lamp was in contact with the milk (65 mm/211 mm), resulting in a UV output of ∼3.68 W. The milk was stirred with a 8 × 40 mm magnetic stirrer bar at 600 rpm (IEC CH2090-001–Industrial Equipment & Control, PTY.LTD, Australia) during the treatment to create vertical flow to expose all milk evenly to the UV-C. Dosage was measured using a UV meter (Gigahertz optic x911–Gigahertz-Optic, Germany) 5 cm from the top of the beaker. Treatment duration was 15 minutes, giving a dosage of 1,510 J/L. Dosage was also calculated as the product of treatment time (seconds) and UV-C power (W) divided by the treated volume (L), and was 2,259 J/L.
Milk bacterial content pre- and posttreatment
All raw milk samples had ≤103 CFU/mL coagulase-negative Staphylococcus. All HP- and UV-C-treated samples were confirmed to be free of bacteria before inoculation.
Spiked milk followed by UV treatment
Each raw milk sample (400 mL; n = 4) was transferred into a 600 mL PYREX glass beaker inoculated with 103 CFU/mL of enterotoxin A and B producing S. aureus and treated as mentioned previously.
Inoculum preparation
Enterotoxin A- and B-producing S. aureus (UWA culture collection 2484) were kindly provided by the School of Biomedical Sciences, University of Western Australia. The bacteria were grown on blood agar (BA) plates (PathWest media, Perth, Western Australia) overnight at 37°C. Several distinct colonies were transferred to 5 mL sterile Luria-Bertani (LB) broth. The suspension was measured in a spectrophotometric plate reader at O.D.450 (EnSpire 2300; PerkinElmer, MA). The culture was diluted with sterile LB broth to have a density of 10 6 CFU/mL and this was confirmed by duplicate plate counts (spreading 100 μL of culture).
Inoculation and incubation
The diluted inoculum was added to each milk sample (raw, HP and UV-C treated) at a volume ratio of 1:1,000 (bacterial broth:milk) resulting in an inoculum of ∼103 CFU/mL. The bacterial inoculum was also inoculated in LB broth at 1:1,000 and used as a growth control.
Sample testing
Bacterial growth and toxin production were measured in LB broth, raw milk (with ≤103 coagulase-negative staphylococcus), and HP DHM (62.5°C for 30 minutes), UV-C treated and spiked DHM then UV-C treated. The samples were incubated at 37°C for 18 hours or at 4°C (refrigerator) for 14 days. An aliquot was collected every 3 hours of incubation from the milk samples incubated at 37°C, and every 24 hours from milk samples incubated at 4°C. Bacterial identification and growth were determined using plate culture (CFU/mL), and toxin was detected using a Staphylococcal enterotoxin reversed passive latex agglutination test (SET-RPLA) (Oxoid, United Kingdom).
Bacterial colony counts
S. aureus growth kinetics were determined using the single-plate serial dilution spotted method and all dilutions (neat to 10−8) were assessed in duplicate on BA plates. Ten microliters of each dilution was spotted in triplicate spots on a duplicate BA plate and incubated overnight at 37°C. The number of bacterial colonies present was determined and averaged, then multiplied by the dilution factor to calculate the number of viable cells, then transformed to a base 10 logarithm. To accurately verify the number of viable bacteria in the samples incubated more than 5 days at 4°C and 6 hours at 37°C, which were expected to have lower viable counts (<100 CFU/mL), 100 μL of each sample was spread plated on BA plates in duplicate, and incubated overnight at 37°C. The number of the colonies was averaged and multiplied by the dilution factor to calculate the number of viable bacteria in each sample, then transformed to base 10 logarithm. The limit of detection for this assay was 10 CFU/mL.
Enterotoxin standard curves
A standard curve was prepared from SEA and SEB (Sigma-Aldrich) to make a series of enterotoxin concentrations from 0.5 to 100,000 ng/mL.
Enterotoxin detection
Assays for enterotoxins A and B were carried out with a SET-RPLA TD900 Kit (Oxoid) according to the manufacturer's instructions.
Results were scanned using a document scanner (Epson V700–Epson, Japan). ImageJ software 27 was used to evaluate the generated images and semiquantitative results were acquired by measuring the pixels for each result in comparison to negative controls and standard curve toxin results to determine the amount of toxin present. Enterotoxin measurements from all raw samples and UV-treated samples were below the limit of detection; therefore, the toxin comparison was only performed with the HP milk samples.
Measurement of lactoferrin and secretory Immunoglobulin A
The lactoferrin content of DHM samples was quantified for all samples incubated at 37°C by the sandwich enzyme-linked immunosorbent assay method as previously described by Czank et al. 5 All antibodies and standards were purchased from MP Biomedicals Australasia (New South Wales, Australia). The recovery assay for lactoferrin was 100.5% ± 3% (n = 6) and for sIgA was 101% ± 4% (n = 7).
Measurement of lysozyme
Determination of lysozyme concentration for all samples incubated at 37°C was carried out using a modified turbidimetric assay. 28 Standards were prepared by diluting hen egg white lysozyme (Sigma, St. Louis, MA) 0.0005–0.01 mg/mL. Skim milk samples were diluted 10-fold in 0.1 M Na2 HPO4/1.1 mM citric acid (pH 5.8) buffer.
Twenty-five microliters of standards or diluted skim milk samples were placed into the wells of a 96-well plate (Greiner Bio-One, Frickenhausen, Germany). One hundred seventy-five microliters of Micrococcus lysodeiltikus suspension (0.075% w/v, ATCC No. 4698; Sigma, St. Louis, MA) was added into each well and the plate was incubated with shaking at RT for 1 hour. The absorbance was measured at 450 nm. Recovery of lysozyme was 102% ± 2.5% (n = 6).
Statistical analysis
Results are expressed as mean ± SD, unless stated otherwise. The growth of S. aureus in raw, HP, UV-C, and spiked then UV-C-treated DHM between 0 to 18 hours was visualized graphically.
When bacterial growth was observed, a logistic growth model was used to explore the relationship between bacterial colony count (log base 10) and incubation time. Segmented regression was used to explore the relationship between bacterial colony count (Log10) and the concentration of enterotoxins.
Linear regression was used to explore the relationship between observed protein levels and time of incubation, treatment, and sample.
The rate of bacterial growth was calculated as the log of bacterial growth at 18 hours normalized by the starting (log) bacterial growth converted to an hourly rate. The rate of bacterial growth was compared with the retention of lysozyme, lactoferrin, and sIgA using a simple linear regression. Significance was set at the 5% level and data were analyzed using the R environment 29 for statistical computing with the package segmented used for fitting the segmented regression models.
Results
Bacterial growth at 37°C incubation
DHM from four donors was treated by Holder pasteurization, UV-C treatment, or was left untreated. A total of 103 CFU/mL of S. aureus was inoculated and bacterial growth was assessed every 3 hours for 18 hours.
The colony count of S. aureus in the HP DHM consistently increased until it reached the stationary phase after 12 hours when incubated at 37°C. The growth rate was similar to the growth of S. aureus in the LB broth (positive control) (Fig. 1a).
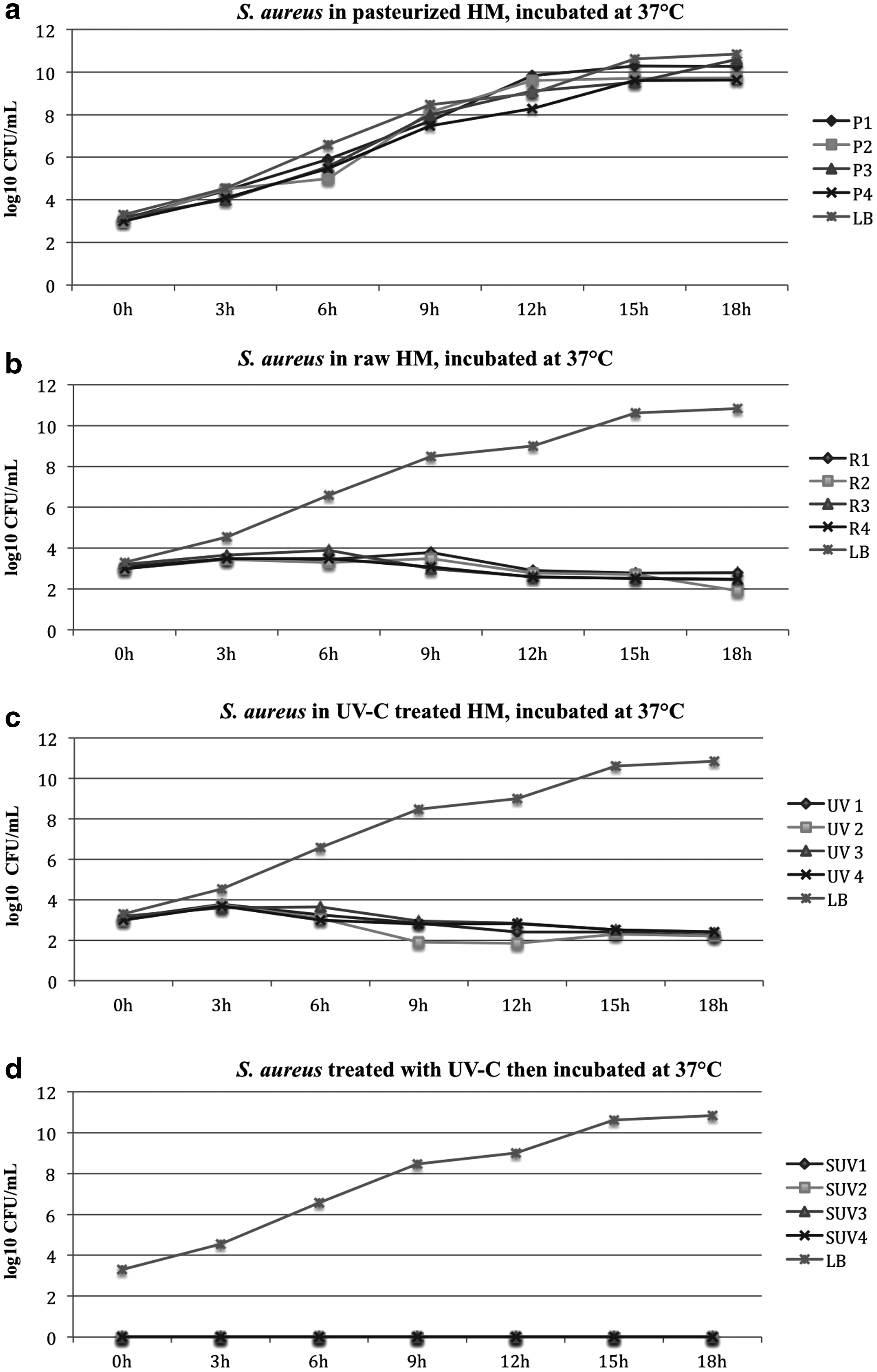
Growth of Staphylococcus aureus in raw, Holder-pasteurized, UV-C and spiked then UV-C treated DHM incubated at 37° C for 18 hours.
The colony count of S. aureus in raw DHM remained stable then decreased minimally when incubated in 37°C (Fig. 1b).
The colony count of S. aureus in the UV-C treated DHM decreased slightly when incubated in 37°C (Fig. 1c).
Bacteria that were inoculated before UV-C treatment were not detected after UV-C treatment and were not detected after 18 hours of incubation at 37°C (Fig. 1d).
Bacterial growth at 4°C incubation
DHM from four donors was treated by Holder pasteurization, UV-C treatment, or was left untreated. A total of 103 CFU/mL of S. aureus was inoculated and incubated at 4°C for 14 days. Growth was assessed every 24 hours.
S. aureus in the HP milk samples incubated at 4°C showed a decrease in the colony count by three-fold within 14 days of incubation (Fig. 2a).
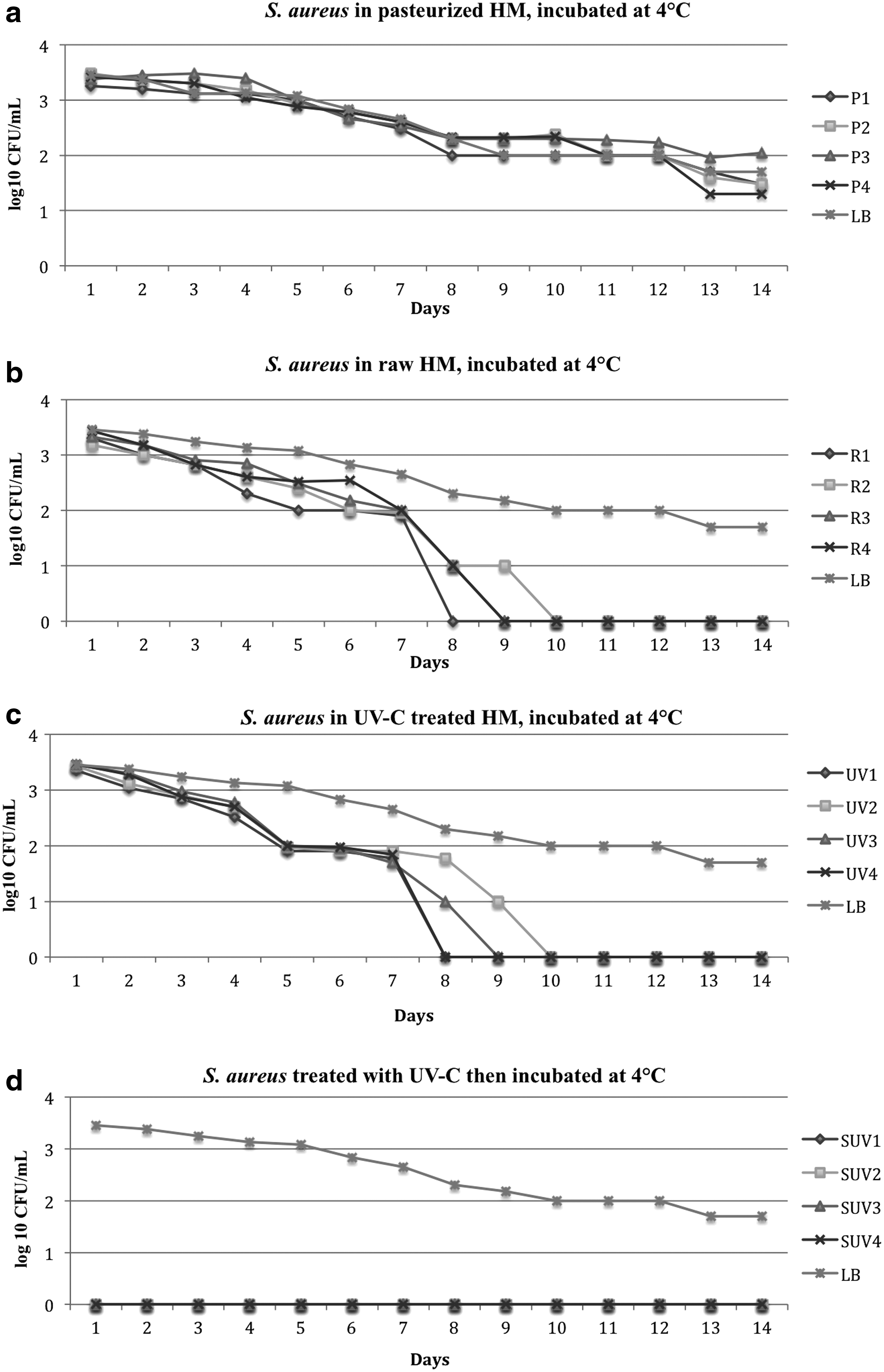
Growth of S. aureus in raw, Holder-pasteurized, UV-C and spiked then UV-C-treated DHM incubated at 4°C for 14 days.
The colony count of S. aureus in raw DHM decreased by threefold (under the limit of detection) between day 8 and 10 of incubation (Fig. 2b).
The colony count of S. aureus in the UV-C-treated DHM (bacteria introduced after treatment) decreased until it was below the limit of detection between day 8 and 10 of incubation (Fig. 2c).
In the milk that was inoculated with S. aureus before UV-C treatment, no bacterial colonies were detected after UV-C treatment and until the end of incubation after 14 days at 4°C (Fig. 2d).
Lactoferrin, sIgA, and lysozyme retention in DHM samples
Protein levels remained constant over the incubation time at 37°C for all three proteins.
The levels varied by sample (all p < 0.001) and treatment (all p < 0.001). HP DHM had a significantly lower concentration of lactoferrin, lysozyme, and sIgA compared with raw milk and UV-C (all p < 0.001) (Table 1).
Concentration of Immunological Proteins in Raw, Holder-Pasteurized, and Ultraviolet-C-Treated Donated Human Milk Samples (Mean ± SD, n = 4)
p-Value <0.001.
UV-C, ultraviolet-C.
There was no significant difference in sIgA, lactoferrin, and lysozyme variation between samples. Bacterial proliferation rate per hour was lower with higher retention of lactoferrin, lysozyme, and sIgA (all p < 0.001) when HM was incubated at 37°C (Fig. 3).
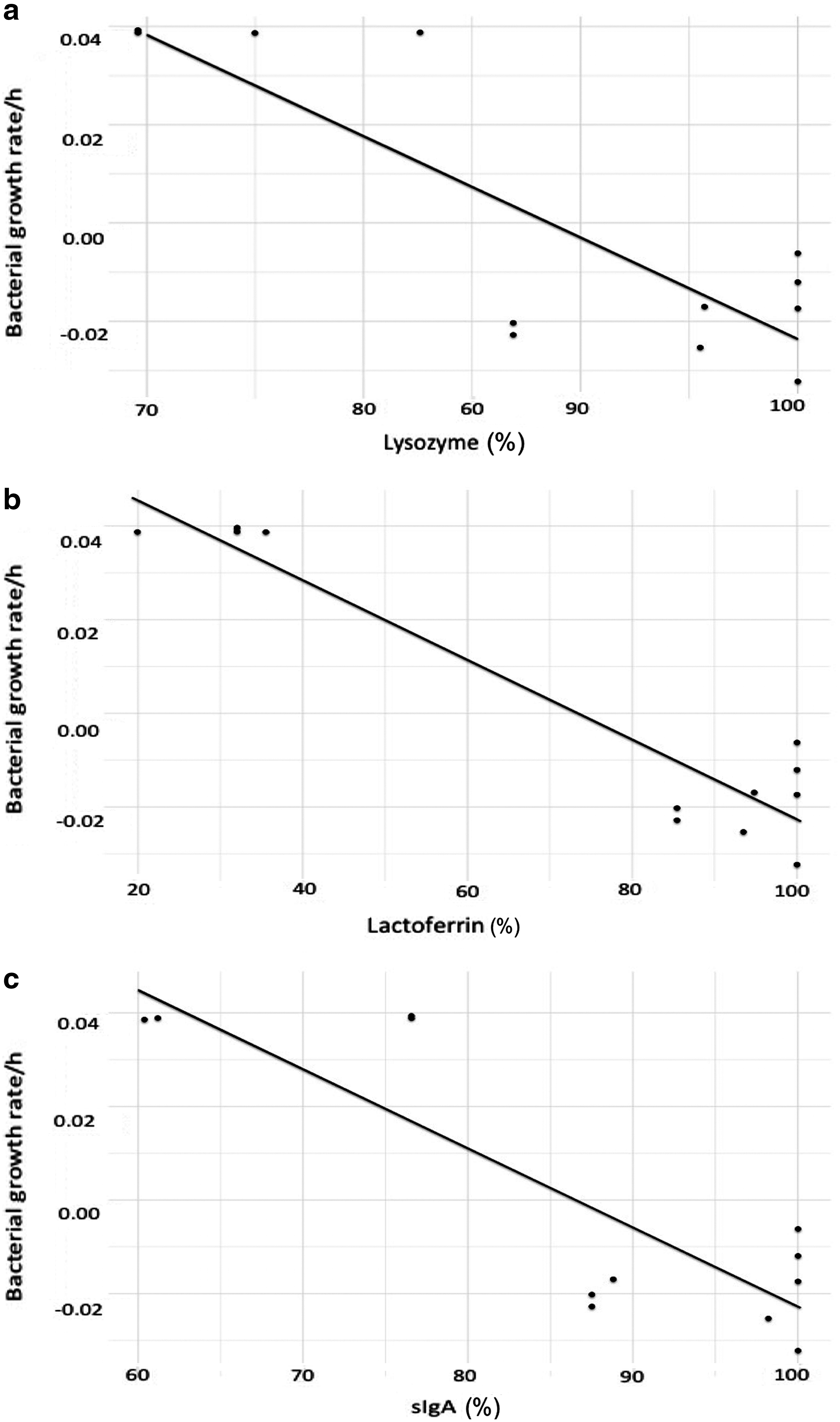
Relationship between S. aureus proliferation rate per hour and retention of
Enterotoxin in DHM
SEA was only detected in HP DHM inoculated with S. aureus and incubated at 37°C from the ninth hour of incubation onward. At this point, the growth was nearing stationary phase and viable counts were approaching 10 8 CFU/mL. The concentration of SEA increased with increasing time and a significant increase in the rate of production was observed after 11.2 hours (p < 0.001), (Fig. 4). Only one sample of HP DHM inoculated with S. aureus and incubated at 37°C showed enterotoxin B at the 18th hour of incubation.
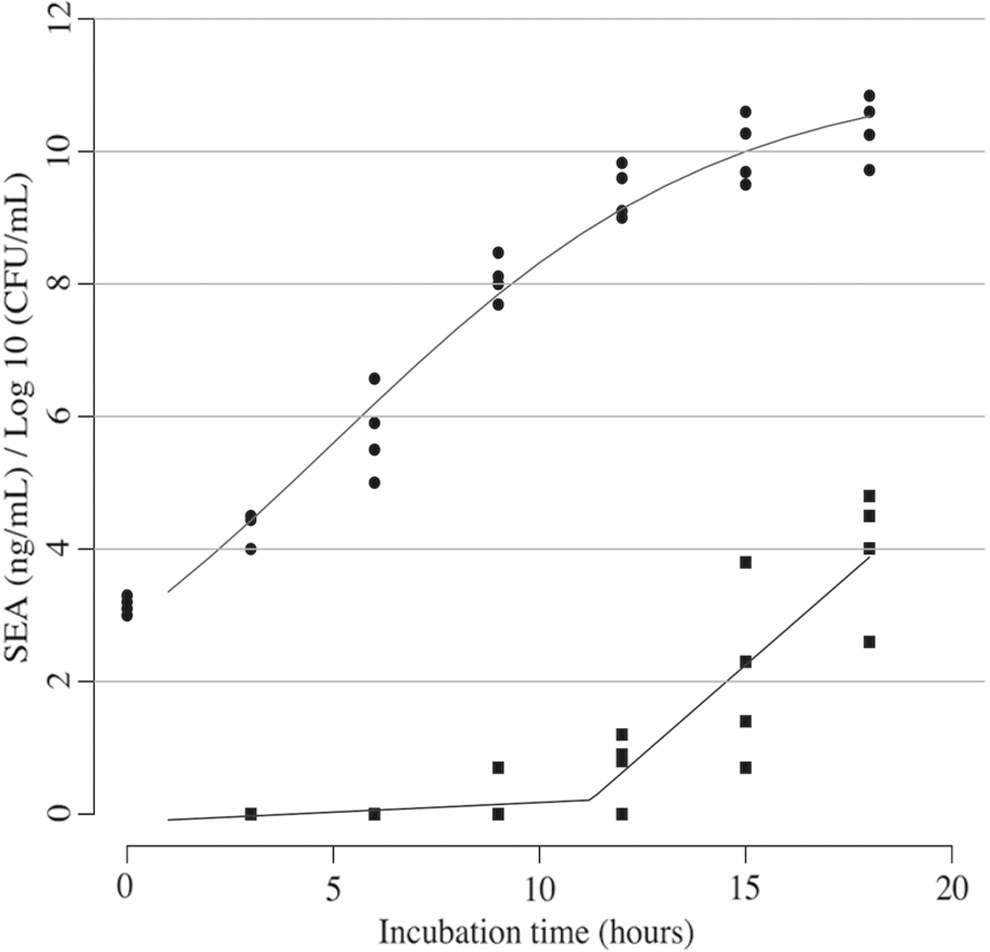
S. aureus growth in milk incubated at 37° C (circles) with the logistic growth curve. SEA production (squares) with segmented linear regression fit. SEA, Staphylococcal enterotoxin A.
Discussion
S. aureus is the most common potential pathogen detected in DHM and is capable of producing enterotoxins that may be harmful to the preterm infant. This study aimed to determine if enterotoxin was produced in HP DHM, UV-C-treated DHM, or inoculated then UV-C-treated DHM. We detected S. aureus enterotoxin in HP DHM samples inoculated with S. aureus after 9 hours of incubation at 37°C, and this was likely due to the reduction in immunological milk proteins after Holder pasteurization.
Proliferation of S. aureus in HP DHM samples was responsible for the production of enterotoxin. Our study showed that SEA production was positively related to bacterial density in all HP samples incubated at 37°C (Fig. 4). This is consistent with a previous study that showed a correlation between the production of enterotoxin in pasteurized cow milk and staphylococcal colony counts. 14 The bacterial colony counts increased in HP HM incubated at 37°C over time until they reached a stationary phase at 15 hours of incubation (Fig. 1a). However, production of enterotoxin B demonstrated different kinetics, being detected at the 12th hour in LB broth incubated at 37°C and only at the 18th hour in a single HP DHM sample. The limited detection of SEB can be attributed to its sensitivity to culture conditions, whereas SEA production is more closely related to the growth kinetics of S. aureus. 30 Furthermore, SEA has been shown to be produced throughout the log phase of S. aureus growth, whereas SEB is produced during the transition from the log phase to the stationary phase. 31 Our findings are consistent with the literature as we detected SEB at the 12th hour between log and stationary phase (Fig. 1a).
In contrast to HP DHM, no enterotoxin production was detected in UV-C-treated and raw DHM when incubated at 37°C, presumably due to the lower bacterial growth kinetics (Fig. 1b, c). The reduced bacterial colony count in the UV-C-treated and raw DHM was expected, due to the preservation of immune proteins in UV-C and raw DHM. The HM immune proteins lysozyme, sIgA, and lactoferrin are the most abundant proteins exerting the majority of antimicrobial activity in HM.32–34 Commensurate with previous studies,16,25 we measured significantly lower concentrations of lactoferrin, lysozyme, and sIgA in HP DHM compared with raw and UV-C-treated DHM. The retention of three major proteins is likely contributing to the higher antibacterial activity and suppression of the bacterial growth in UV-C-treated DHM (Table 1). This assumption is further supported by the inverse correlation between the bacterial growth rate and the retention of lactoferrin, lysozyme, and sIgA (Fig. 3). Furthermore, an optimum condition for S. aureus growth is a lack of bacterial competition. The presence of bacterial competition will result in lower bacterial proliferation and consequently minimal amounts or no enterotoxin production. This has been shown in raw or fermented foods and unpasteurized bovine milk.14,35 Such an environment (without competing microorganism) was produced in the HP DHM and UV-C-treated HM (treated preinoculation). However, UV-C HM showed a similar colony count to raw HM, whereas the colony count increased in HP DHM indicating the importance of HM immune proteins as bactericidal agents.
HP DHM samples did not produce detectable enterotoxin when incubated at 4°C. The absence of enterotoxin can be attributed to the decreasing bacterial load in the samples, which was in the order of one- to two-fold over the 14 days of incubation (Fig. 2a). Enterotoxin was not detected in UV-C-treated DHM or raw milk. The decrease in colony count by three-fold by the 10th day of incubation (Fig. 2c) in UV-C-treated DHM and the complete absence of bacterial colonies by day 9 (Fig. 2b) in raw milk explains the lack of enterotoxin production. This observation was consistent with previous studies that showed bacterial levels in raw and leftover HP DHM samples that were contaminated with commensal bacteria, reduced significantly when stored in the refrigerator (4°C) for 4 to 6 days.36,37 In addition, the rapid decline of S. aureus in CFU/mL in UV-C-treated raw DHM stored at 4°C compared with the less pronounced decline of the same inoculum in HP DHM may be due to the loss of immune proteins after heat treatment in the latter samples, 38 which has also been shown in previous studies.5,39,40
The experiment to determine if S. aureus is able to proliferate or produce enterotoxin after treatment of inoculated milk with UV-C was reassuring; the spiked bacteria were unable to proliferate after treatment (Fig. 2d). Thus, no enterotoxin production was detected during incubation at both 37 and 4°C. This somewhat allays the concern that bacteria damaged by UV-C may still be able to produce enterotoxin where treatment protocols are able to inactivate all viable bacteria. UV-C treatment of DHM is therefore an attractive alternative to heat pasteurization.
The limitations of this study are the detection limit of the enterotoxin assay (detection limit 0.5 ng/mL), which raises the slim possibility that enterotoxin might be produced below the level of detection. However, Evenson et al. showed that the dose of enterotoxin to produce gastroenteritis in school-aged children is 0.5 ng/mL. 41 The dosage required to cause complications in preterm infants remains unknown.
Conclusion
This study has shown that pasteurizing DHM with UV-C has potential as an alternative method to Holder pasteurization to provide safe, high-quality HM for preterm infants. We have shown that enterotoxin was only detected in HP DHM samples by the 9th hour of incubation at 37°C. Enterotoxin was not detected in UV-C, raw, and inoculated then UV-C-treated DHM at 37 or 4°C. Enterotoxin production was related to bacterial growth, which was restricted by the high level of immune protein bioactivity in UV-C-treated milk.
Footnotes
Acknowledgments
This research was funded by an unrestricted grant from Medela AG, Switzerland. Medela AG provides an unrestricted research grant to D.G. Taibah University, Saudi Arabia, provides a PhD scholarship for Y.A. Medela AG and Taibah University had no input into the design, analysis, interpretation, and publication of the study.
Disclosure Statement
No competing financial interests exist.