
Abstract
Background:
The Human Milk Banking Association of North America (HMBANA) sets the purity and quality standards for donor human milk (DHM) in the United States, which includes zero bacterial presence after Holder pasteurization. This study aimed to determine if nutrient and bacterial composition of DHM with limited bacterial presence after pasteurization change over 4 days of refrigerated storage.
Methods:
Twenty-five unique samples of DHM with limited bacterial growth postpasteurization were collected from two HMBANA milk banks. Infant formula was used as a comparison. Samples were stored in the refrigerator and a portion of milk was removed at 24-hour intervals beginning at hour 0 to 96 for analysis. Aerobic bacteria, protein, lactose, and immunoglobulin A (IgA) content were measured. Longitudinal changes between 0 and 96 hours were analyzed using repeated measures analysis of variance and mixed models test. p < 0.05 was deemed significant.
Results:
There was no significant difference in lactose, protein, bacteria, or IgA content over storage duration (p = 0.649, p = 0.690, and p = 0.385, p = 0.805, respectively). Total aerobic bacteria were less than 102 colony-forming units (CFUs) in 81% of the time points tested for DHM samples. Total aerobic bacteria were too many to count (>300 CFUs) in the infant formula sample at all time points.
Conclusion:
In periods of high demand for DHM, DHM with low bacteria growth postpasteurization may be an option as a supplemental food for the growing number of healthy infants who receive DHM. Future studies should investigate the bacterial strains in this milk.
Introduction
Human milk has been well characterized and is considered the optimal nutrition for newborn infants by the American Academy of Pediatrics, The World Health Organization, and the United States Surgeon General. 1 Current policies recommend infants be exclusively breastfed for the first 6 months of life, and then introduction of complementary foods with continued breastfeeding for at least 1 year. 2 When mother's own milk is unavailable, donor human milk (DHM) is the recommended feeding strategy, especially for the preterm infant who is at risk for necrotizing enterocolitis.2–4
Guidelines for the production and distribution of DHM in the United States are primarily determined by the Human Milk Banking Association of North America (HMBANA). 5 In the United States, DHM undergoes Holder pasteurization where the milk is warmed to 62.5 degrees Celsius for 30 minutes, which is effective in inactivating microbial agents that may be present, while maintaining many of the beneficial components of DHM.6,7 Safety standards of pasteurized DHM include testing each batch for presence of bacteria after pasteurization. If one bacterial colony-forming unit (CFU) is present, the milk is discarded. 8 These stringent standards are in place to protect the vulnerable hospitalized preterm infant because milk banks have historically primarily served hospital Neonatal Intensive Care Units (NICU), with 88% of level 3 and 4 NICUs across the United States utilizing DHM.9,10 However, other, less vulnerable infant populations have been increasingly using DHM, which may not require such stringent standards. 11
Two recent studies investigated the percentage of DHM rejected due to postpasteurization bacterial growth and found that up to 12.6% of DHM is discarded because of bacterial presence.12,13 Studies have reported no relationship between presence of bacteria in HM and infection in premature infants, which is in contrast to HMBANA's current guidelines for zero bacterial presence.14,15
As a comparison, the dairy industry allows up to 20,000 CFUs per milliliter of dairy milk as part of the Grade A pasteurized milk ordinance. 16 CFUs are not addressed in the FDA regulations for powdered infant formula (PIF), but PIFs must be assessed for the presence of Cronobacter and Salmonella species. 17 All storage studies to evaluate shelf-life of DHM have been conducted in DHM with no bacterial presence after pasteurization; no study has looked at DHM that contains minimal bacterial growth postpasteurization. This DHM has the potential to be repurposed to less vulnerable populations instead of being discarded.
The purpose of this study was to determine if nutrient (protein, lactose, and immunoglobulin A [IgA]) and bacterial composition of DHM with limited bacterial presence after pasteurization change over 4 days of refrigerated storage. We hypothesized that protein, lactose, IgA, and bacterial content would not significantly change over 4 days of storage at 4°C.
Methods
Twenty-five unique samples of deidentified DHM that had been rejected due to postpasteurization bacterial growth were collected from two HMBANA milk banks, Mother's Milk Bank Northeast (Newton, MA) and King's Daughters Milk Bank at CHKD (Norfolk, VA). Powdered infant formula (Gerber Good Start GentlePro Powder Infant Formula, Stage 1) was used as a comparison because formula is the recommended food when human milk is not available. DHM samples were shipped overnight on dry ice and stored at −20°C until sample preparation. This study was classified as non-human subjects research by the University of North Carolina Greensboro Institutional Review Board (protocol 17-0523).
Sample preparation
The 100–120 mL bottles of DHM were defrosted in the refrigerator at 4°C. Once samples were defrosted, bottles were gently mixed by inverting 10–12 times by hand. Baseline samples, which represent time 0, were removed (∼10 mL), aliquoted into appropriate amounts, and stored in the freezer (at approximately −20°C) until analysis. The remaining DHM was returned to the refrigerator at 4°C and stored for 96 hours to simulate home storage.
Four days of storage were chosen for this study because it is twice as long as the current HMBANA guidelines for defrosted DHM storage which is 48 hours. 5 To simulate a “home environment,” the primary investigator washed hands, but did not wear gloves, a laboratory coat, or other PPE, or disinfect the countertop before each removal. Once in the refrigerator, at hours 24, 48, 72, and 96, the primary investigator gently mixed each bottle by inverting 10–12 times by hand and removed ∼10 mL of sample that was aliquoted into necessary amounts, labeled with the time of removal, and stored in a freezer (approximately −20°C) until analysis. Temperature of the refrigerator was recorded at each removal (0, 24, 48, 72, and 96 hours).
Sample analysis
Lactose was measured using the Megazyme enzymatic assay (K-LAGCAR, Megazyme International, Wicklow, Ireland), based on the AOAC method 2006.06 which utilizes three reactions to quantify lactose in human milk. 18 Lactose concentrations were calculated from equations generated from a standard curve of known lactose monohydrate (64044-51-5; Fisher Scientific, Fair Lawn, NJ). Standards and samples were measured in triplicate using an Epoch Bio-tek Microplate Spectrophotometer Plate Reader (7091000; BIO-TEK, Winooski, VT).
Total protein was measured using Pierce™ Bicinchoninic Acid (BCA) Protein Assay Kit (23225; Thermo-Scientific, Waltham, MA). 19 This assay uses the peptide bonds in proteins to reduce Cu2+ to Cu1+. Cu1+ is then chelated by two molecules of BCA, which forms a purple complex that absorbs light at 562 nm. To determine the concentration of protein in the sample, it is compared to the absorbance of a standard bovine serum albumin with known protein concentrations. Standards and samples were measured in triplicate using an Epoch Bio-tek Microplate Spectrophotometer Plate Reader.
Total aerobic bacteria were measured by a plate enumeration method with a range of detection of 0–300 CFUs (3M Petrifilm® Aerobic Plate Counts, 70200572124; 3M, St. Paul, MN). Sterile peptone was used as a control. Samples were undiluted and measured on a single plate. Plating was performed under a biosafety bench with sterile pipette tips. Plate counting was conducted under a biosafety hood with a hand counter and magnifier. If CFUs were too numerous to count (>300), they were designated a CFU count of 300 for analysis.
IgA activity was measured using an enzyme-linked immunoassay (ELISA) developed by Chen.20,21 The IgA first binds to an Escherichia coli antigen, developed using eight E. coli strains acquired from the STEC Center (Michigan State University, East Lansing, MI), which is then bound to an anti-human-IgA antibody. The antibody is labeled with Horseradish Peroxidase enzyme (HRP, A0295; Sigma Aldrich, St. Louis, MO), which, in the presence of hydrogen peroxide, catalyzes the conversion of the E. coli-IgA complex in the presence of 2,2′-azino-bis-3-ethylbenzothiazoline-6-sulfonic acid (ABTS, A-1888-5G; Sigma Aldrich) into a colored product. A spectrophotometer measures the color intensity at 405 nm, which is then compared to known concentrations of human IgA standards (I2636; Sigma Aldrich). Standards and samples were measured in triplicate using an Epoch Bio-tek Microplate Spectrophotometer Plate Reader (Synergy HT; BioTek Instruments, Winooski, VT).
Statistical analysis
Sample size was calculated based on the number of samples necessary to detect a 20% change in lactose concentration with an 80% power and an alpha value of 0.05 was found to be 14. We chose a sample size of 25, which aligned better with similar previous studies. Lactose was chosen for power calculations because if bacteria are growing in the DHM, they would likely be fermenting the lactose, thus decreasing its content in the samples.
Statistical analysis was conducted using SPSS software (version 28) and Microsoft Excel (Version 16.42). Longitudinal changes in nutrients, bioactive factors, and bacteria over 96 hours of storage were evaluated with repeated measures analysis of variance test and linear mixed model. p < 0.05 was deemed significant. Sample ID was treated as a random effect.
Results
Lactose, IgA, and protein analytical methods were conducted in triplicate and the average coefficients of variation were as follows: 2.7% for lactose, 5.7% for IgA, and 2.5% for protein. R2 values were >0.997 for IgA, >0.999 for lactose, and >0.989 for protein. There was zero aerobic bacterial growth for the sterile peptone control. Descriptive statistics for DHM samples are reported in Table 1.
Mean (Range) Values for Nutrient and Bacterial Content of Donor Human Milk (n = 25) Over 96 Hours of Refrigerated Storage
Longitudinal changes in nutrients, bioactive factors, and bacteria over 96 hours of storage were evaluated with repeated measures analysis of variance test and linear mixed model. There was no significant change in nutrient or bacterial composition throughout storage duration.
CFU, colony-forming unit; IgA, immunoglobulin A.
Changes over refrigerated storage duration
Linear changes in content of lactose, IgA, protein, and bacteria are shown in Figure 1. There was no significant difference in lactose, IgA, protein, or bacteria content in DHM over 96 hours of refrigerated storage (p = 0.649, p = 0.805, p = 0.690, and p = 0.385, respectively).
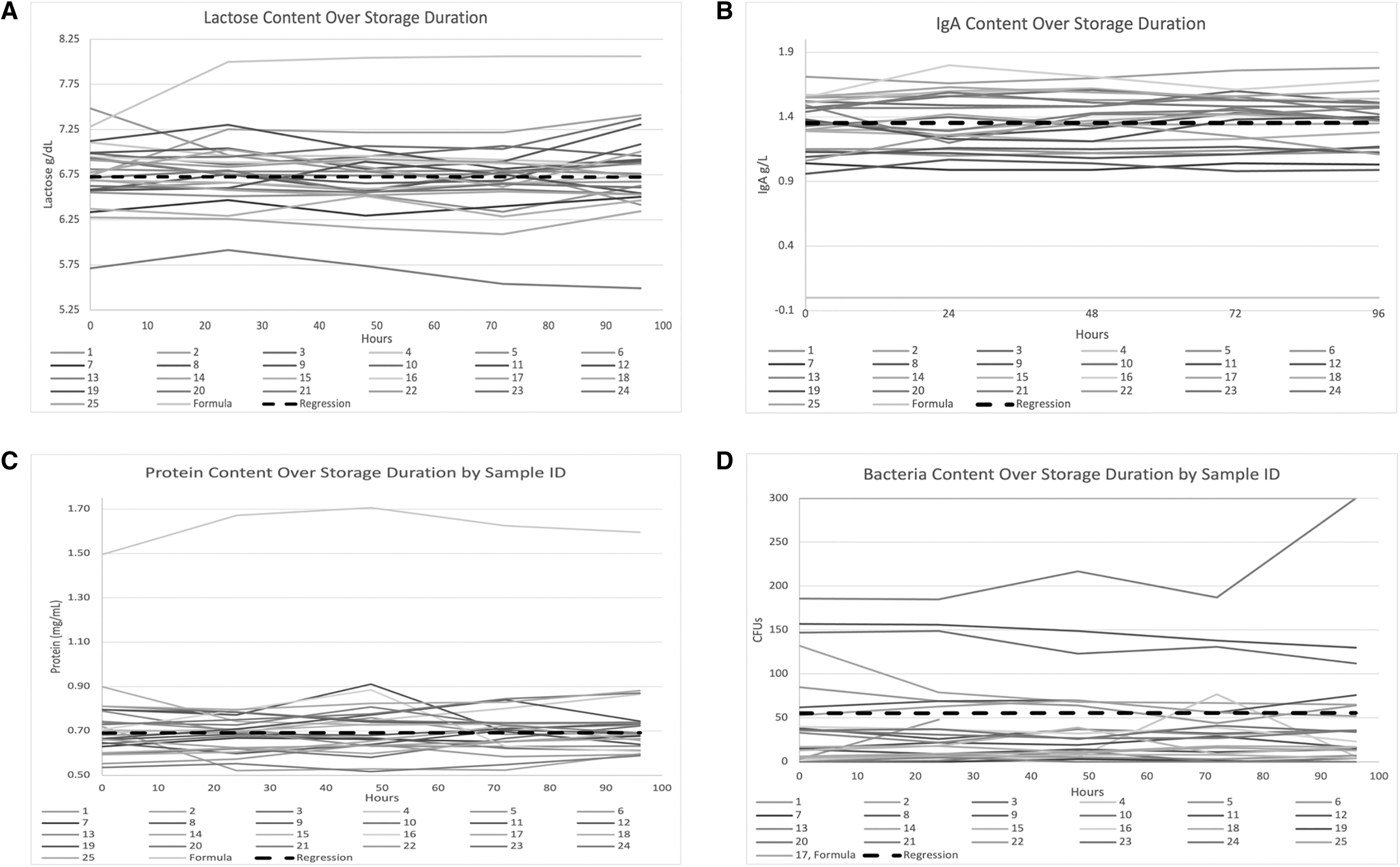
Nutrient or bacterial content by sample ID over storage duration (96 hours).
Total aerobic bacteria were less than 102 CFUs in 81% of the time points tested for DHM samples. Total aerobic bacteria were too many to count (>300 CFUs) in the control sample (infant formula) at all time points. The infant formula contained an average 1.2 g/dL lactose and 0.9 g/dL protein higher content compared to the DHM samples and no IgA.
Discussion
There has been no study to investigate the quality and storage of DHM that has bacterial presence after Holder pasteurization. Historically, this DHM has been discarded. This study aimed to investigate bacterial and nutrient changes in DHM that had been rejected by HMBANA milk banks due to limited bacterial growth postpasteurization. Over 4 days of storage at 4°C, we found no significant change in aerobic bacteria, IgA, lactose, or protein content. These findings are in agreement with previous literature that investigated refrigerated storage of DHM that had been accepted postpasteurization (without bacterial presence). Mandru et al. and Vickers et al. found that the bacterial content of pasteurized DHM did not change over 4–9 days of refrigerated storage under clinical handling and storage settings.22,23
Moreover, a review by Schlotterer et al. found that three studies reported there was no change in bacterial content in DHM over 4–9 days of refrigerated storage, with one conducted using home settings. 24 Prior studies evaluated DHM that had passed the initial requirement of 0 CFUs postpasteurization. The results from prior studies along with the results of this study suggest that DHM with and without bacteria postpasteurization maintains some of its antimicrobial properties and prevents bacterial growth over 4 days of refrigerated storage under clinical and home settings. In addition, Schlotterer et al. investigated protein, lysozyme, and IgA in DHM that had no bacterial growth postpasteurization and found no significant change over 96 hours of refrigerated storage. 21 These align with the results from this study, suggesting that DHM with and without limited bacteria postpasteurization maintains its nutrient composition over 4 days of refrigerated storage.
The majority (81%) of DHM samples in our study contained less aerobic bacteria compared to the infant formula. Powdered infant formula is not analyzed for bacterial levels. Instead, it is analyzed for certain strains of bacteria that are known to be pathogenic, specifically Salmonella and Cronobacter species. 17 Bacteria are innate in human milk and previous research of Schanler et al. found no correlation between bacterial content in mother's milk and infection risk in extremely premature infants. 14
This suggests the limited bacteria that remain in DHM postpasteurization may be of no greater risk than bacteria found in powdered infant formula or native human milk. However, a limitation of this study is that we did not analyze the DHM for the strains of bacteria in the milk, including spore-forming bacteria such as Bacillus cereus. Future studies should analyze the bacteria strains present in DHM, which contains bacteria postpasteurization to determine if any are potentially pathogenic.
The relevance of this study is exemplified by the recent formula shortage in the United States that began in February 2022. While the American Academy of Pediatrics released a statement urging the White House to take action to address this crisis and offered potential solutions for caregivers feeding their infants formula, many caregivers still struggled to find formula to feed their infants, with some attempting to make their own infant formula. 25 By not discarding the DHM with limited bacteria after pasteurization, this milk could be utilized by term infants during periods when mother's own milk and/or formula is unavailable.
DHM is increasingly being used for term infant. 11 However, it is still unknown if DHM is nutritionally adequate for the term infant. Castro et al. found that vitamin C levels are significantly reduced in DHM compared to mother's milk, suggesting that infants receiving primarily DHM should receive a vitamin C supplement. 26 Future studies should investigate other nutrients in DHM that are necessary for optimal growth and development of infants before DHM is recommended as an alternative primary source of food for the term infant.
Conclusion
DHM with limited bacterial presence postpasteurization maintains its bacterial and nutrient content over 4 days of refrigerated storage, with 81% of the DHM samples containing less aerobic bacteria compared to infant formula. Based on these findings, this DHM with limited bacterial presence may be an option as a supplement for healthy term infants to remain on an exclusively human milk diet. Further investigation is warranted to determine the species of bacteria present in this milk.
Footnotes
Authors' Contributions
S.A.M. and M.T.P. contributed to study conception, study design, and data interpretation. S.A.M. had primary responsibility for data collection and writing the article. Both authors reviewed, edited, read, and approved the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.