
Abstract
Preservation of sperm at ambient storage temperature (25°C) provides a simplified alternative for short-term storage applications. The goals of the study were (i) to improve the storage ability of bovine sperm in liquid suspension by scavenging the reactive oxygen species (ROS) with the help of antioxidants and chelators, (ii) to develop a predictive tool to determine sperm motility as a function of storage temperature. Two types of antioxidants (catalase and superoxide dismutase, SOD) and 2 chelators (Desferal and ethylenediaminetetraacetic acid, EDTA) were used. Different concentrations of the supplements were added in egg yolk Tris (EYT) extender and stored at 4°C, room temperature (25°C), and 37°C, respectively. Motility and membrane integrity (MI) were assessed on days 1, 4, 7, 10, and 14 for samples stored at 4°C and 25°C. Results did not suggest the beneficial effect of SOD and EDTA. Catalase caused a slight improvement in motility at 4°C storage and its effectiveness was found to increase at higher storage temperature. The presence of Desferal resulted in a much higher increase in motility and membrane integrity at both 4°C and 25°C. Similarly, the fitting of Arrhenius damage model to the motility data provided a useful tool to predict motility degradation time as a function of storage temperature.
Introduction
S
A major factor contributing to the disruption of cellular function in sperm during storage at near ambient conditions is the formation of reactive oxygen species (ROS), which are mainly composed of peroxides (H2O2), superoxide anions (O2·−) and hydroxyl free radicals (OH·).4–8 Studies have shown that the reactions producing these free radicals are more active at higher storage temperatures than in the cryopreserved state.5,9 ROS molecules combine with cellu-lar components and transform into oxygen-based damage products. In addition to ROS, a cellular environment contains several endogenous redox active metal ions.10 These metal ions, especially iron and copper, react with the peroxides to form the hydroxyl radicals through the Fenton reaction.11 Sperm plasma membranes are highly sensitive to peroxidation caused by free radicals because of high content of unsaturated fatty acids in their membranes.12 Lipid peroxidation causes the membrane to be leaky and leads to complete loss of motility and membrane integrity.13 Since sperm do not possess any internal repair mechanism against cellular damage,5 an environment where damage is minimized is essential to protect it from ROS-induced damage. For similar reasons, a reduced oxidative stress environment is equally desirable in cryopreservation and the partial desiccation storage of mammalian cells (storage of cells at near ambient temperature in the partially desiccated state).14–18
The goal of the study was to increase the short-term storage ability of bovine sperm in liquid suspension by reducing the oxidative stresses through the use of antioxidants and chelators. The hypothesis is that the addition of an optimal amount of the supplements helps reduce the oxidative stresses, thus minimizing motility loss and membrane and/or mitochondrial damage. Antioxidants counteract the formation of superoxide anions and peroxides while chelators sequester the free metal ions, thereby minimizing the production of hydroxyl radical by the transition metal (mainly iron).19,20 Two different antioxidants namely catalase and SOD, and 2 metal chelators namely deferoxamine (Desferal) and EDTA were used. SOD scavenges superoxides into oxygen and hydrogen peroxide and catalase decomposes peroxides to water and oxygen.21,22 Desferal is an iron chelator while EDTA forms strong complexes with many divalent and trivalent metal ions including Ca2+, Mg2+, Zn2+, Mn2+, Fe2+, Fe3+, Cu2+, Pb2+, and Co3+. The supplements were selected owing to their scavenging properties and their widespread application in sperm against ROS damage and oxidative stresses.5,23−25 Two different modes of sperm injury were assessed, namely the motility and the plasma membrane integrity. In addition to assessing the effect of the supplements on cellular damage, the study also investigated the sperm degradation kinetics at 3 different storage temperatures (4°C, 25°C, and 37°C). Motility data for these temperatures were fitted into an Arrhenius model.26 The resulting Arrhenius plot would make it possible to predict the motility degradation time (τ) for any storage temperature within the temperature range (4°C–37°C).
Materials and Methods
Semen shipments
Fresh bovine semen samples from Holstein bulls were shipped every week from ABS Global, WI, via United Parcel Service (UPS) next day service. The sperm were diluted in a 1:2 ratio with EYT extender and placed in a 15-mL conical tube. EYT was composed of 20% egg yolk (by volume), 2.42 weight % Tris (hydroxymethyl aminomethane), 1.38 weight % citric acid monohydrate, and 1.0 weight % fructose. During shipment, the tube was packed in a foam brick refrigerant that maintained the temperature at 4°C. The sample was stored in a refrigerator at 4°C upon arrival. Progressive sperm motility was typically in the range of 80%–85% and the membrane integrity was over 90% immediately on arrival.
Method of analysis
Sperm were suspended and stored in 1-mL Eppendorf tubes at a concentration of 10 million sperm/mL in EYT extender on the day the samples were received (day 1). Before introducing the sperm to the extender, supplements were added at different concentrations. Catalase from bovine lever (Catalog # C40), Desferal (Catalog # D9533), EDTA (Catalog # 03960), and SOD (Catalog # S5395) were purchased from Sigma Aldrich. The concentrations included (i) catalase: 0.1, 0.05, and 0.01 mg/mL; (ii) SOD: 500, 100, and 10 U; (iii) Desferal: 5, 1, and 0.1 mM; and (iv) EDTA: 5, 1, and 0.1 mM. Sperm suspended in EYT was used as control for all the experiments. The samples were stored at 3 different temperatures: 4°C, 25°C, and 37°C. For 4°C and 25°C storage samples, motility and membrane integrity were assessed on days 1, 4, 7, 10, and 14. Values were normalized with respect to the motility/membrane integrity values of the control on day 1. For the 37°C storage experiments, samples with catalase and Desferal were selected. Motility was assessed for these samples at 0, 6, 8, 16, 24, and 48 h.
Evaluation of motility
Motility was evaluated visually under a bright field microscope (Nikon Eclipse TS 100) using a glass slide (Fisher Scientific, Catalog # 22-339-408) covered with a coverslip. A sample size of 10 µL and an objective lens of ×20 magnification were used to observe motility. Motility is expressed as the percentage of progressively motile sperm as a fraction of the total number of sperm (motile + immotile). The reported values are cumulative numbers of motile and immotile sperm observed at several fields in the sample slide with a field area of 0.025 mm2. This was done to address the possible spatial disparity that can arise at different fields within the same sample slide.
Evaluation of membrane integrity
Membrane integrity assessment was performed using the Hoechst 33342 (Sigma Aldrich, St. Louis, MO; Catalog # B2261) and Ethidium Homodimer (Sigma Aldrich; Catalog # E1903). The staining media was prepared by adding 10 µL of stock solution of Hoechst and 5 µL of Ethidium Homodimer to 1 mL of EYT. All evaluations were performed on samples mounted on glass slides with coverslip on top. They were viewed under a fluorescent microscope (Nikon Eclipse E600) using the red (G-2E/C TRITC) and blue (UV-2A) excitation/emission filters at a magnification of ×20. Membrane integrity was expressed as the ratio of the number of sperm stained blue minus the number stained red over the number of sperm stained blue.27
Arrhenius damage model
The Arrhenius damage model was used to evaluate the degradation pattern based on the motility data for different storage temperatures. The equation is expressed in the following forms:


where τ is the characteristic time for a given temperature and was selected as the time (in hours) at which the normalized motility value reached 1/e (36.7%). Ea is the activation energy (kcal/mol) and A is the frequency factor (s−1), T is the temperature (K), and R is the universal gas constant (kcal/mol-K).
Equation 2 was used to generate the Arrhenius plot for the control and samples with Desferal, catalase, and Desferal + catalase. The τ values were calculated from the experimental data of motility at different storage temperatures. A plot of the natural logarithm of τ vs. 1000/T was generated to determine the values of the Arrhenius parameters, Ea and A. Ea is given by the slope of the linear curve that fits the experiment data and A is given by the value of the y-axis intercept when 1000/T = 0.26
Statistical analysis
The experiment values represent the average of 4 repeats. The error bars represent the standard deviations of the repeats. The statistical significance of the motility and membrane integrity data were analyzed using a standard single factor ANOVA test in Microsoft Excel 2003. Significance was assessed as P ≤ 0.05.
Results
As mentioned in the materials and methods section, motility was evaluated for 4°C storage conditions for a range of antioxidants and chelators concentrations. The motility values for these different concentrations are shown in Table 1. The results suggest a lower motility when the additives were present at the highest concentrations. Based on the results, a single concentration was selected for membrane integrity tests and other motility tests at higher temperature. The concentrations that were selected for further analysis include 1 mM Desferal, 0.05 mg/mL catalase, 100 U SOD, and 1 mM EDTA.
M
Abbreviations: EDTA, ethylenediaminetetraacetic acid; SOD, superoxide dismutase.
Effect of antioxidants
The plots in Figure 1 show the motility of sperm supplemented with (i) catalase, (ii) SOD, and (iii) catalase + SOD. Figure 1A is a normalized motility plot of samples stored at 4°C. The presence of catalase caused a slight improvement in motility compared to the control. Motility on days 7, 10, and 14 were statistically significant in the presence of catalase (P = 0.012, 0.025, and 0.015, respectively). The values were 57.4%, 33.9%, and 15.4% compared to the corresponding control values of 45.2%, 27.1%, and 8%. On the other hand, there was no significant improvement in motility either in the presence of SOD or a combination of catalase and SOD (P > 0.05).
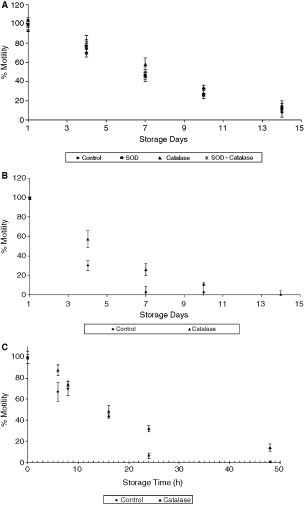
Effect of antioxidants on sperm motility at different temperatures. (
During the study of room temperature storage, samples containing SOD degraded after a week of storage and produced a pungent odor. A check of motility did not detect any motile sperm. Hence, SOD was not included in the higher temperature studies. The normalized motility of sperm at room temperature storage in the presence of catalase is shown in Figure 1B. There was a rapid decrease in motility for the control samples with an almost complete motility loss by day 7. The addition of catalase yielded significantly higher motility (P < 0.05) on days 4 and 7. There was, however, large motility drops during longer storage period. The values were 11% on day 10 and 0% on day 14.
Unlike with storage at 4°C and 25°C, motility was reduced to the order of hours at 37°C (Fig. 1C). Compared to the controls, samples with catalase showed significantly higher motility at longer storage periods. The corresponding motility values were 6.8% and 31.8% at the end of 24 h and were 0.5% and 14% at the end of 48 h. In summary, there was an improvement in sperm motility in the presence of catalase but no significant effect in the presence of SOD. Moreover, there was a progressive improvement in the effect of catalase at higher temperatures than at 4°C storage.
On the other hand, membrane integrity values were not significantly affected by catalase or SOD. The normalized membrane integrity values of the samples with catalase and SOD at 4°C storage are plotted in Figure 2A. While the motility values in the presence of catalase were statistically significant on days 7, 10, and 14, the corresponding membrane integrity values were not statistically different (P ≥ 0.05). A comparison of the membrane integrity curve with the corresponding motility curve shows a distinct degradation pattern. The rate of decrease in sperm motility was much more rapid than membrane integrity. Even for the controls, there was a big difference in the motility and membrane integrity values over longer storage period. The values were 8% and 47.3% on day 14. Figure 2B shows the normalized membrane integrity of the sperm suspensions stored at room temperature. Similar to 4°C storage results, the membrane integrity for samples with catalase was not statistically different from the control (P = 0.16).
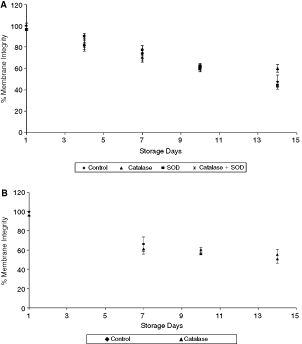
Effect of antioxidants on sperm membrane integrity at different temperatures (
Effect of chelators
Figure 3 shows the motility of sperm in the presence of chelators. EDTA and Desferal had opposite effects on sperm motility at 4°C storage (Fig. 3A). Motility of the sperm in the presence of Desferal was much higher and statistically significant compared to the control on days 7, 10, and 14. The addition of EDTA caused a significant decrease in sperm motility on days 7, 10, and 14 (P > 0.05). On day 14, the motility values for the (i) control and the samples supplemented with (ii) EDTA and (iii) Desferal were 8%, 5.4%, and 32.8%, respectively. Figure 3B represents the normalized motility plot for the sperm suspended at room temperature in the presence of Desferal. Results showed a rapid decrease in motility for the control samples, with an almost complete loss of motility by day 7. On the other hand, the presence of Desferal helped maintain motility, which were at 46% and 26% on days 10 and 14.
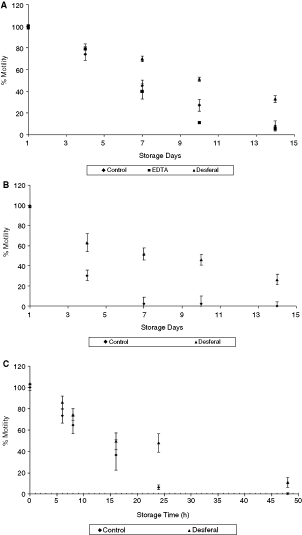
Effect of chelators on sperm motility at different temperatures (
The motility curve for 37°C storage (Fig. 3C) showed significantly higher values in the presence of Desferal. The control samples suffered a complete motility loss in about 24 h of storage. The addition of Desferal provided a longer motility survival with complete motility loss occurring beyond 48 h. Motility at 24 h of storage was 6% and 45.1%, respectively, for the control and samples with Desferal. In summary, Desferal caused a significant improvement in sperm motility while EDTA resulted in a further deterioration.
Membrane integrity results showed opposing effects of EDTA and Desferal at 4°C, similar to the corresponding motility results (Fig. 4A). The presence of EDTA caused a significant decrease in membrane integrity on days 10 and 14 (P = 0.033 and 0.0143). Desferal, on the other hand, caused an increase in the membrane integrity values. Statistical analyses showed the membrane integrity of the Desferal containing samples to be statistically higher than the controls on days 7, 10, and 14 (P = 0.0381, 0.0047, and 0.0013). Figure 4B shows the effect of Desferal on membrane integrity at room temperature storage. Samples with Desferal yielded a significantly higher membrane integrity values on all storage days. The values for the controls and samples with Desferal were 50.3% and 60.6% on day 14.
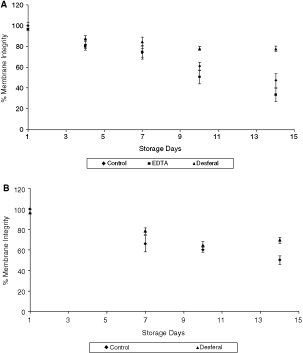
Effect of chelators on sperm membrane integrity at temperatures (
Effect of antioxidants + chelators
In addition to investigating the individual role of antioxidant and metal chelators, the synergistic effect of the antioxidants plus Desferal was also examined. Figure 5A is the motility plot for 4°C storage. The combinations included catalase + Desferal, SOD + Desferal, and SOD + catalase + Desferal. Compared to the control, motility was significantly higher on days 7, 10, and 14 in the presence of these additives (P ≤ 0.05). The presence of Desferal + catalase at room temperature storage also showed a significantly higher motility in comparison to the control (Fig. 5B). However, when these results were compared to the effect of Desferal only samples, the values were not statistically different. This indicates that the higher values of the samples with the antioxidants and Desferal mixture were contributed mainly by Desferal. The effect of Desferal + catalase at 37°C storage was a significant increase in sperm motility as compared to the controls (Fig. 5C). Motility was 49.9% and 15% after 24 and 48 h of storage. However, these values were similar to those of catalase only or Desferal only samples (38.6% and 11.5% for catalase and 48% and 11% for Desferal). The results, therefore, do not suggest any significant improvement in motility due to the combination of antioxidants and chelator.
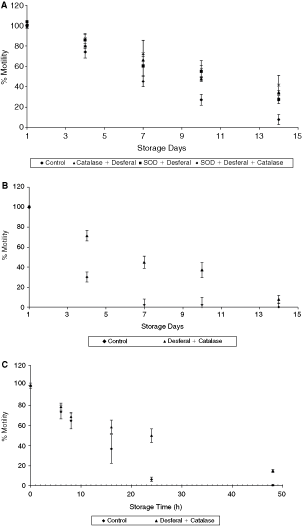
Effect of antioxidants + chelators on sperm motility at different temperatures. (
The membrane integrity values of the combined effect of antioxidants and Desferal at 4°C storage and 25°C storage are shown in Figure 6A and 6B. Though the values were higher than the control on all storage days, the samples exhibited neither a beneficial effect nor statistical significance when compared to the Desferal only samples. The higher membrane integrity of the sperm can therefore be attributed to Desferal in the mixture.
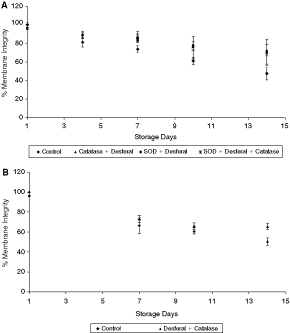
Effect of antioxidants + chelators on sperm membrane integrity at temperatures. (
Arrhenius plot
Based on the motility data of samples stored at different temperatures (4°C, 25°C, and 37°C), an Arrhenius plot was generated to study the temperature dependence of degradation kinetics. The τ value, calculated as the time for the normalized motility to drop to 1/e value (36.7%), is shown in Table 2. Figure 7 shows the Arrhenius plot for the control and samples containing Desferal, catalase, and Desferal + catalase. Result shows that the control samples exhibit a linear degradation behavior in the temperature range tested. The value of activation energy was 12.1 kcal for the controls (Table 3). On the other hand, samples with catalase, Desferal, and Desferal + catalase do not exhibit a similar trend. While the catalase samples still show a significant decrease in τ value at room temperature storage (compared to 4°C storage), samples with Desferal and Desferal + catalase show an almost stable survival up to room temperature after which they undergo a rapid decrease when further exposed to 37°C storage. As shown in Table 3, the values of the activation energy were much lower in the 4°C–25°C range (<5 kcal/mol) compared to the 25°C–37°C range (>20 kcal/mole). On a different perspective, the room temperature (25°C) served as a break point, at which there was a transition from a gradual motility loss at lower temperatures to a much steeper decline in τ values beyond that temperature. While the values were 60, 115, 278, and 206 h (in the order: control, catalase, Desferal, and catalase + Desferal) at 25°C, they were 16, 33, 24, and 38 h at 37°C.

Arrhenius plot showing the effect of catalase and Desferal. Filled diamonds (♦) represent the control samples. Filled squares (▪) represent the samples with catalase. Filled triangles (▴) represent samples with Desferal. Asterisks (∗) represent the samples with catalase + Desferal. Dashed and dotted ( __ - __ - ) line represents the linear fit to the control samples. Dashed ( __ __ ) line represents the linear fit to the samples with catalase. Dotted line ( - - - - ) represents the linear fits to the samples with Desferal. Continuous line ( ______ ) represents the linear fit to the samples with catalase + Desferal.
V
Note: τ represents the time at which the normalized motility fraction drops to a value of 1/e (36.7%).
V
Discussion
The current study was distinct from the existing oxidative stress study of various mammalian cells types in that it was motivated from the storage perspective. The study was centered on arresting the 3 major ROS (superoxide anions, hydrogen peroxide, and the hydroxyl free radicals). Superoxide anions have a short half-life (1 ms) and break down into peroxides spontaneously or enzymatically.24 H2O2 is relatively more stable compared to the super oxide anions or the hydroxyl radical. It has a long half-life and accumulates in the media after being generated.28 On the other hand, hydroxyl radical can be formed by reaction between superoxide anion and hydrogen peroxide (by Haber-Weiss reaction), or by the reduction of peroxide by ferrous ion (the Fenton reaction).19,20,24,29 The deleterious effect of hydroxyl radical lies in its extremely high reactivity, a very short half-life (of the order of 1 ns) and hence its inability to be eliminated by enzymatic actions.25
Effect of antioxidants
We had investigated the effect of SOD and catalase in scavenging the superoxide anions and hydrogen peroxides, the first 2 compounds in the ROS chain formation oxidative reactions.12 The enzymatic action of SOD and catalase are summarized by the following reactions:
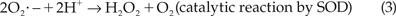

Our motility and membrane integrity results from 4°C storage do not suggest a beneficial effect of SOD. This ineffectiveness implies that superoxide anion may not be a major ROS species in the extender media. The ineffectiveness could also be attributed to their short half-life after which they spontaneously break down into peroxides.25,30,31
The presence of catalase in the storage media had a pronounced effect on sperm motility. The extent of its effectiveness was, however, a function of the storage temperature. At 4°C storage conditions, there was no big improvement in motility even though the values were higher and statistically significant from the controls (P ≤ 0.05) on days 7, 10, and 14. The results, however, show a statistically significant and much higher motility at 25°C storage, which tend to suggest that the enzymatic action of catalase is more effective at higher storage temperature as compared to 4°C storage. On the other hand, catalase did not provide a significant improvement in membrane integrity. The lower effectiveness of the scavengers in inhibiting membrane damages also applied to the other additives under study. Our control values suggest that the membranes are less affected by oxidative stresses, and this justifies the lesser role of the additives in protecting the sperm membrane integrity.
Various literatures show that catalase provides a good defense against peroxide-induced sperm motility loss.5,24,32 In one such study of the ROS-mediated loss of bovine sperm motility in EYT extender, Bilodeau et al. have shown that the addition of only 5 U/mL of bovine liver catalase or oviductal fluid catalase (OFC) was sufficient to overcome the loss of sperm motility caused by 100 μM hydrogen peroxide (H2O2) in EYTG and TALP extenders after 6 h of incubation.24 In yet another study of the protective effect of catalase during sperm storage, Vishwanath et al. have reported a much higher improvement in bovine sperm motility that lasted for about a month when they were stored at ambient temperature (18°C–21°C) and suspended in CAPROGEN, a citrate-based medium.5 Though our results show some improvement in sperm motility, the extent of protection exerted by catalase is much lower than reported by Vishwanath et al. This could possibly be due to the different media that they had used for their storage experiments. While their control samples (sperm suspended in CAPROGEN media) exhibited a slow decrease in motility over an extended time period (3–4 weeks), our samples stored in EYT extender showed a much rapid decrease in motility even at cold storage (4°C) condition. We also investigated the combined synergistic effect of SOD and catalase. However, there was no significant improvement in motility and membrane integrity compared to the lone effect of catalase. Since superoxides and peroxides are normal products of cellular metabolism, our results suggest that the extraneous addition of the antioxidant enzymes do not significantly impart the desired protection against the target ROS. This could be due to the impermeability of the scavenger enzymes to penetrate through the membranes and reach the site of formation of the ROS.11 On the other hand, H2O2 is membrane permeable but superoxide is not. Thus part of the H2O2 generated in the sperm cells may leak out of the cell to the media and subsequently be scavenged by the catalase supplement. This may explain the weak protection by catalase.
Effect of chelators
Compared to the antioxidants, the presence of Desferal resulted in a much higher and statistically significant (P ≤ 0.05) motility and membrane integrity values, both at 4°C and 25°C. The protective effect was more pronounced during longer storage period (on days 10 and 14) by which time the motility for the controls drop to much lower values. Desferal is an iron chelator that scavenges free Fe3+ ions, which could be present in the semen extender in the form of impurities or contributed by the dead sperm accumulated during storage.33 According to Guérin et al.,19 the iron complexes (Fe3+) present in a cellular environment catalyzes the reaction of superoxide (O2·−) with hydrogen peroxide to produce molecular oxygen (O2), OH−, and hydroxide radical (OH·), the last being the most reactive metabolite contributing to oxidative damages. The adverse catalytic effect of free ferric ion can be summarized by the following reactions, also known as the Haber-Weiss reaction:

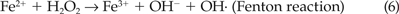

The mechanistic role of Desferal can thus be attributed to its inhibition of the formation of the hydroxyl radical that would initiate lipid peroxidation chain reaction in membrane lipids.19,20 The synergistic effect of Desferal with catalase was also investigated. Though the results showed a higher motility and membrane integrity values as compared to the controls, the values were statistically insignificant in comparison to the samples with Desferal only. These observations, coupled with the relatively insignificant role of catalase, suggest that Desferal was the major factor in imparting protection to the sperm in our experimental storage conditions.
Unlike Desferal, the addition of EDTA resulted in decreased motility for longer storage period (day 7 onward). Similarly, a significant decrease in membrane integrity was observed on days 10 and 14. This may be due to the fact that unlike Desferal, which inhibits the Fenton reaction, EDTA promotes the Fenton reaction when it chelates iron.10,34 Although study has shown that a small millimolar concentration of EDTA (0.5 and 1 mM) enhanced the straight line velocity in human sperm over a period of 4 h,35 a higher concentration of EDTA (∼17 mM) has been proven to sequester the Ca2+ ions (which is a major regulatory factor of sperm motility) thereby rendering the sperm immotile.35 According to Lee et al., EDTA decreases the human sperm motility in a dose-dependent manner and also as a function of exposure time.36 The negative effect of EDTA may therefore be attributed to promoting Fenton reaction and/or reduction of Ca2+ ion concentration. Our results suggest that EDTA should not be used as the chelating agent due to its adverse effect in sperm motility and membrane integrity.
Effect of storage temperature
The scavenging effect of catalase and Desferal was studied at 3 different temperatures: 4°C, 25°C (room temperature), and 37°C. The purpose was to obtain the degradation kinetics as a function of storage temperature. Motility results showed that the storage period was of the order of 2 weeks at 4°C and 25°C, and of the order of a day at 37°C storage. A comparison of motility between similar samples at 4°C and 25°C storage conditions showed significantly higher values at 4°C for the controls and the samples with catalase. However, in the presence of Desferal, the room temperature motility was close to the corresponding 4°C values. Though the numbers were higher at 4°C, they were statistically insignificant on days 10 and 14 (P = 0.43 and 0.11). On the other hand, a comparison of the motility values between the controls and the catalase samples at 25°C showed that catalase was more effective at room temperature compared to 4°C storage. Similarly, the comparison of the 25°C motility values between the catalase and the Desferal samples showed close values on days 1–3. These observations suggest a more effective enzymatic action of catalase at room temperature compared to 4°C storage. On the other hand, the statistical insignificance of the motility values on days 10 and 14 offer a possibility of room temperature storage for short-term applications in the presence of the Desferal.
Arrhenius plot
The Arrhenius plot provided a comprehensive summary of the nature of motility degradation as a function of storage temperature. The control samples exhibited a linear Arrhenius behavior in the temperature range tested. Contrary to the controls, samples with catalase, Desferal, and Desferal + catalase did not show a linear Arrhenius behavior. The Arrhenius characteristics for these samples can be defined as a combination of 2 intersecting linear fits, one between 4°C and 25°C (with an almost stable τ value), and the other between 25°C and 37°C (with a rapid decrease in motility values). The plot also provides useful information about the effectiveness of the supplements at a particular temperature. Results show that while Desferal provided significant protection at 4°C, catalase did not. The values of the time constants for the controls and the catalase samples were essentially overlapping at this temperature. Catalase was, however, more effective at higher temperatures as shown by the higher values of ln(τ). The effect of the additives (both Desferal and catalase) was more pronounced at 25°C storage. Though there was a sharp decrease in τ values at 37°C, similar values for samples with catalase and Desferal indicate an increased effectiveness of catalase at higher temperatures.
More importantly, the usefulness of the Arrh enius plot lay in its ability to act as a predictive tool to determine the degradation time (τ) at a given storage temperature and within the temperature range under study (4°C–37°C). The Arrhenius fits can be used to match the corresponding τ value (in the y-axis) for a particular storage temperature (in the x-axis), thus simplifying the process of predetermining the degradation behavior for the control samples and samples with catalase and/or Desferal.
Conclusion
Antioxidants and chelators impart membrane protection through the inhibition of oxidative stress that causes motility and membrane integrity loss. Our results suggested that motility is more susceptible among these 2 injury modes. Of the different antioxidants and chelators that were used, results showed the beneficial effect of Desferal, and catalase to a lesser extent. However, no beneficial effect was shown for SOD, while EDTA showed adverse effect. Addition of Desferal imparted better motility and membrane integrity protection at all the storage temperatures under study. The role of Desferal can be attributed to the chelation of iron cations that would otherwise react with the peroxide and superoxide anions to form hydroxyl ions. Similarly, the results from the 25°C experiments indicate the possibility of room temperature storage for short-term storage applications. Although catalase did not provide a major protection at low temperature (4°C), its effectiveness increased at higher storage temperatures (25°C and 37°C) and was also shown to be more effective for shorter storage periods. The temperature-dependent Arrhenius plot indicated (i) linear degradation of τ in the 4°C–37°C temperature range for the controls and (ii) dissimilar degradation patterns in the 4°C–25°C and 25°C–37°C temperature range in the presence of Desferal or catalase. The usefulness of the Arrhenius plot laid on its ability to predict motility degradation at different storage temperatures.
Footnotes
Acknowledgment
We are thankful to ABS global, DeForest, WI, for funding this research.
Author Disclosure Statement
The authors have no conflicts of interest to disclose.