
Abstract
The Siberian tiger ear marginal tissue fibroblast cell line (STF34) from 34 samples was successfully established using primary explants technique and cell cryoconservation technology. STF34 cells were adherent, with a population doubling time of 24 h. Chromosome analysis showed that 90.2%–91.6% of cells were diploid (2n = 38). Isoenzyme analyses of lactate dehydrogenase and malate dehydrogenase showed that STF34 cells had no cross-contamination with other species. Tests for cell line contamination with bacteria, fungi, viruses, and mycoplasmas were all negative. Every index of the STF34 cell line meets all the standard quality controls of American Type Culture Collection. Not only has the germline of this important Siberian tiger species been preserved at the cell level, but also valuable material had been provided for genome, postgenome, and somacloning research.
Introduction
To preserve the wealth of biodiversity in China, there is a very urgent need to commence rigorous conservation of endangered species. 4 Establishing fibroblast lines from endangered species has been proposed as a practical approach for this purpose; not only does it preserve precious genetic material, but also it provides an excellent resource for biological research. 5
In our report, we used the combined method of microorganism detection, chromosome analysis, isoenzyme analysis, and fluorescent protein genes transfection to detect the established Siberian tiger cell line (named STF34 line) overall. Our project is to cryopreserve this international protected genetic resource for the purposes of reviving endangered breed by cloning and supplying a convenient and effective resource for genetic research.
Materials and Methods
Cell culture
Ear tissue samples were sampled from 34 Siberian tigers (16 male and 18 female), then rinsed, chopped into 1-mm3 pieces, and seeded on the surface of a tissue culture flask containing Dulbecco's Modified Eagle’s Medium (DMEM) + 10% fetal bovine serum in a 37°C incubator with 5% CO2. 4 Cells at the logarithmic phase were harvested. The cell sheet was rinsed twice with phosphate-buffered saline to remove all traces of trypsin inhibitors in the serum and 0.25% trypsin solution was added. The flasks were inverted and incubated for 3 min at 37°C before being turned over. Subsequently, the flasks were shaken gently to detach cells from their walls and the medium was added to the cell suspension to stop the trypsinization. The cells were split into new flasks under the ratio 1:2 or 1:3 and incubated at 37°C with 5% CO2.
Cryogenic preservation and recovery
The cells were supplemented with fresh medium at 24 h prior to freezing, to make sure the nutrition was sufficiently absorbed by the cells. The cell suspension was acquired by digesting cells in 0.25% trypsin. The suspension was centrifuged at 1000 rpm for 8 min and supernatant was abandoned. The harvested cells were resuspended in freezing medium [10% dimethyl sulfoxide (DMSO) + 50% fetal bovine serum + 40% DMEM] to a final density of (2–3) × 106 viable cells/mL. Subsequently, 1 mL of the cell suspension was allocated into 1 sterile plastic cryogenic vial labeled with the animal name, gender, cell line, passage number, and date. The vials were sealed and kept at 4°C for 20–30 min to allow DMSO to equilibrate, and then after programmed freezing for 12 h, they were transferred to a liquid nitrogen storage system quickly and efficiently.4,6 To recover and reseed the cells, the frozen tubes were removed from liquid nitrogen and quickly thawed in 42°C water bath, and then the cells were transferred into a flask with complete medium and cultured at 37°C with 5% CO2.
Growth curve and estimation of cell viability by trypan blue staining
The cells were plated on 24-well microplates, with a density of 1.5 × 104 cells/well, and were cultured for 7 days. Cells were counted every 24 h until the plateau phase. Mean values were used to plot a growth curve and population doubling time (PDT) was calculated based on the growth curve. 7 Assays of cell viability before freezing or after recovery were carried out by using the trypan blue vital stain method, and 1000 cells were counted for viability.
Measurement of microorganisms in STF34 cell line
Detection of contamination with bacteria, fungi, and yeasts
The detailed procedure used for bacteria, fungi, and yeasts contamination testing was referred from the work by Doyle et al. 8
Testing for viruses
Routine examination for cytopathogenic effects using phase-contrast microscopy was performed following Hay's hemadsorption protocol. 9
Mycoplasma detection
The cells were stained with Hoechst 33258, and enzyme-linked immunosorbent assay Mycoplasma Detection kit (Roche) was used. 10
Chromosome analysis and isoenzyme analysis
Chromosomes were prepared, fixed, and stained following standard methods. 11 After Giesma staining, the chromosome numbers per spread were counted for 100 spreads under an oil-immersion objective. Relative length-to-arm ratio and centromeric index and type were counted according to the protocol of Sun et al. 12
Isoenzyme patterns of lactic dehydrogenase (LDH) and malic dehydrogenase (MDH) were detected using the vertical slab noncontinuous polyacrylamide gel electrophoresis assay according to the protocol of Wu et al. 5 Mobility was measured as the ratio of distance migrated by the band to the distance migrated by the indicator dye.
Expression of fluorescent protein genes in Siberian tiger fibroblastic cell
To obtain the highest transfection efficiency and low cytotoxicity, transfection conditions were optimized by varying cell density and concentrations of fluorescent protein vectors pEGFP-N3, pEGFP-C1, pECFP-N1, pECFP-mito, pDsRed1-N1, and pEYFP-N1 plasmid DNA (BD Biosciences Clontech) and using Lipofectamine™ 2000 (Invitrogen), according to Lipofectamine media methods. To estimate the transfection efficiency, the cells were observed under excitation wavelengths of 405, 488, and 543 nm separately, using confocal microscopy (Nikon TE-2000-E).
Results and Analysis
Morphology of fibroblasts from Siberian tiger
We used a primary explants technique and cryogenic preservation technology to establish the STF34 cell line and proceeded to biology and genetics detection. Fibroblast-like or epithelial-like cells could be seen migrating from the tissue pieces at 5–12 days after explanting (Fig. 1A). The cells continued to proliferate and were subcultured when they reached 90% confluence (Fig. 2B). After 2–3 passages, we obtained the purified fibroblasts (Fig. 1C). In addition, they were viable and grew well after they were thawed (Fig. 1D–F). The motilities of fibroblasts before freezing and recovery measured by trypan blue staining were 98.5% and 96.7%, respectively.

Culture and morphology of Siberian tiger fibroblast cell (×100). (

Growth curve, detection of microbial contamination, and karyotype of STF34 cells. (
Growth curve (cell dynamic state observation) and microbial analysis
The growth curve of the STF cell line before cryopreservation and after recovery displayed a typical “S” shape (Fig. 2A) and the PDT was 24 h. There was a lag time or latency phase of about 24 h after seeding, corresponding to the adaptation and recovery of the cells from protease damage, and then the cells proliferated rapidly and entered exponential phase. Tests for contamination with bacteria, fungi, and yeasts were negative. No viruses were indicated by cytopathogenic evidence or by the hemadsorption test. Under a fluorescent microscope, fibroblast nuclei stained with Hoechst 33258 appeared as blue ellipses, showing that the established cell line was mycoplasma negative (Fig. 2B).
Karyogram and the chromosome number of Siberian tiger
The chromosome number of Siberian tiger was 2n = 38, consisting of 36 autosomes and 2 sex chromosomes, XY or XX (Fig. 2C). According to the opposite length, the centromere position, and trabant, we divided the autosomes into 5 groups: group A: 2, 4, 7, 11, and 13 are metacentric chromosomes (M); group B: 1, 5, 8, 9, 10, 12, 16, and 17 are submetacentric chromosomes (SM); group C: 3 and 6 are subtelocentric chromosomes (ST); group D: 14 and 15 are telocentric chromosomes (T); group E: 18 is a metacentric chromosome, trabant. The heterosomes (X, X) were the metacentric chromosomes (Table 1). The karyotype composition of the Siberian tiger is 12 (M) + 16 (SM) + 4 (ST) + 4 (T), XY (M, M). The chromosome numbers per spread were counted for 100 spreads of the first, second, and fourth passages, and the frequencies of cells with 2n = 38 were 91.6%, 91.2%, and 90.2%, respectively.
M: 1.0–1.6 metacentric chromosome; SM: 1.7–2.9 submetacentric chromosome; ST: 3.0–6.0 subtelocentric chromosome; T: ≥7.0 telocentric chromosome.
Isoenzyme analysis of Siberian tiger cell line
The LDH bands obtained from Siberian tiger were compared with those from other species or breeds, and five isoenzyme bands (LDH-1, 2, 3, 4, 5) were observed (Fig. 3A). Enzymatic activities were in order LDH-3, LDH-2, LDH-4, LDH-5, LDH-1; LDH-2, LDH-3, and LDH-4 were dominant, whereas LDH-1 and LDH-5 were scarcely observable.
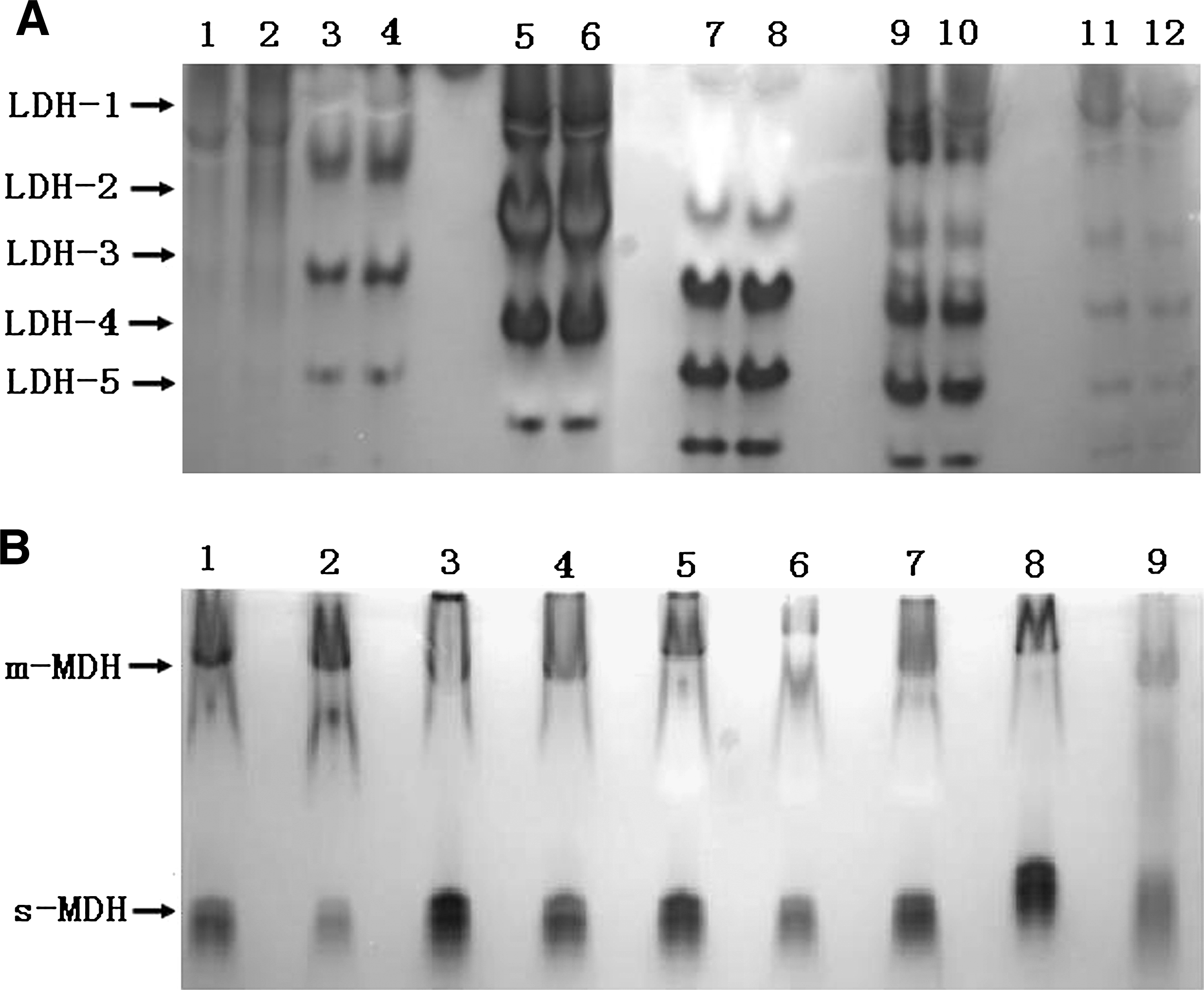
LDH and MDH zymotypes of STF34 cell line. The animals had their own unique bands with a different relative mobility. These results showed that there was no cross-contamination between different species. (
Two MDH isoenzyme bands [cytosolic MDH (s-MDH) and mitochondrial MDH (m-MDH)] were observed in the Siberian tiger (Fig. 3A), and there was a single m-MDH band near the cathode and a single s-MDH band near the anode (Fig. 3B). Similar activity was seen from both m-MDH and s-MDH. There were significant differences in the isoenzyme patterns of LDH and MDH between the Siberian tiger and other cell lines in our laboratory.
The expression results of six fluorescent protein genes in Siberian tiger fibroblastic cell
Positively expressing cells were observed at 12 h after transfection and the numbers and intensity increased markedly and reached a maximum at 48 or 72 h. The transfer efficiencies of the cyan fluorescent proteins (pECFP-C1 and pECFP-mito; Fig. 4D) were significantly lower than those of the green, yellow, and red fluorescent proteins (P < 0.01), and the green fluorescent proteins (pEGFP-C1 and pEGFP-N3; Fig. 4A, B) were maximal. The expression efficiencies of the 6 fluorescent proteins were between 5.4 and 32.8. The numbers of fluorescent cells decreased at 1 week, but a few dispersed positive cells remained after 2 weeks and even after 1 and 2 months. By screening G418 (Geneticin) resistance selection and monoclonic culture for 1 month, we obtained 3 positive cell strains that expressed EGFP, EYFP, and DsRed stably.

Expression of pEGFP-C1, pEGFP-N3, pECFP-C1, pECFP-mito, pEYFP-N1, and pDsRed1-N1 in STF34 cell. Comparative figures of 6 fluorescent proteins at 24 h after transfection using TE-2000-E inverted microscope with excitation wavelengths of 433–588 nm to determine the transfection efficiency (×100). (
Confocal fluorescence microscopy was used to observe the distribution of green, yellow, and red fluorescence in the Siberian tiger cells to determine the location of fluorescent proteins. ECFP and DsRed were mostly shown in cytoplasm, whereas EGFP and EYFP showed an intense nuclear signal (Fig. 4G–I).
Discussion
Establishment of Siberian tiger group fibroblast cell line
Somatic cells, such as diploid fibroblasts, can be used as nuclear donors, and they can be subjected to genetic manipulation in vitro. Morphological results indicated that the primary Siberian tiger cells are usually mixed with epithelial cells. Different tolerances to trypsin were evident as the fibroblasts fell off first when treated with trypsin and were adherent again quickly after passage, whereas most epithelial cells were not adherent or stably adherent and fell off only when treated with mechanical agitation. 13 For this reason, purified fibroblasts could be obtained after 2–3 passages. 14 The procedures used in this study conformed to the protocols of the American Type Culture Collection (ATCC) technical bulletin for primary culture, subculture, and freezing. 15 The average viability after thawing was >90%, and freezing had little influence on the viability of the fibroblasts. Hence, it is likely that the Siberian tiger genome can be preserved by freezing fibroblasts in liquid nitrogen for long-term storage. Because too many passages and excess trypsin digestion could adversely affect the biological characteristics, especially hereditary characteristics, all cells should undergo a minimum number of passages.
Karyotype analysis and mycoplasma detection of STF34
Microbial contamination is the most frequent pollution phenomenon in cell culture. The turbidity of culture media contaminated by bacteria, eumycetes, and mycetes can be seen with the naked eye. Viruses can be seen under the microscope. But it is harder to detect mycoplasma contamination. Staining with the DNA fluorochrome Hoechst 33258 is the most effective and frequently used method for detecting mycoplasma contamination. 16
Karyotypic and isoenzyme data together can effectively confirm the origin of a cell line and identify possible cross-contamination. The practice of combining them has become a classical and standard method for characterizing cell lines and is routinely used by the main biological resource centers around the world (ATCC, ECACC, DSMZ, and Riken).17,18 Because of the fact that we want to conserve the genetic character of Siberian tiger, the fibroblasts must maintain the same diploid character as the cells in vivo. The result showed that the proportion of cells with a 2n = 38 was >90%. Chromosome analysis can relate a cell line to the gender of the animal from which it was derived and also distinguish between normal and malignant cells, because the chromosome number is more stable in normal cells. Jimei and Zhang reported that the chromosomal diploid number for Siberian tigers and South China tigers was 38, which was consistent with our study.19,20 These results indicate that Siberian tiger fibroblasts maintain good growth state and normal chromosomal diploid number after culturing in vitro.
Isoenzyme analysis
LDH and MDH are species-specific and constant, but the enzyme contents and activities differ among species, providing a biochemical indicator of species classification by chromatography and electrophoresis.
LDH is a tetrametric molecule; the H and M subunits are produced by expression of the laha and lahb genes.18,21 Zhou et al. and Arai et al. analyzed the LDH isoenzyme pattern in the leukocytes and plasma of Debao horse and pony fibroblasts.14,20 Their results showed that the plasma and Debao pony fibroblast LDH isoenzyme patterns were dominant for LDH-1, LDH-2, and LDH-3; LDH-3 and LDH-4 were dominant in the leukocytes. The present study on the LDH isoenzyme patterns of Siberian tiger fibroblasts showed that LDH-2, LDH-3, and LDH-4 were dominant. MDH is a dimeric enzyme comprising s-MDH and m-MDH subunits. The mobilities of MDH bands among poultry are essentially identical, and the same is true among mammals. But MDH from mammals migrates more rapidly than that from poultry, and the enzyme content is also greater than in poultry. In the present study, the isoenzyme bands of LDH and MDH in Siberian tiger cells were clear, indicating that the genetic characteristics were stable and there was no contamination with other cells.
Expression of fluorescent protein genes
Fluorescent proteins were widely applied as probes for intracellular dynamics and as useful tools in superresolution microscopy and were proven to be excellent tools for live-cell imaging studies. 23 We considered that the expression efficiency of exogenous genes could also be used as an efficient method to identify and evaluate the quality of cell lines cultured in vitro. Fluorescent proteins with stable structures, high expression levels, and species-independent efficiency have been used as marker genes to observe the expression, contribution, and function of target proteins in live cells and organisms. Under optimal conditions, transfection had no significant effect (P > 0.05) on cell morphology, growth, apoptosis rate, or reduplication status compared with untransfected control. 24 The fluorescent protein-positive cell strains observed here will provide donor cells for transgenic animal cloning.
Overall, the newly established STF34 cell line had all characteristics required by the cell line identification criteria of ATCC. The STF34 line provides a useful approach for conserving this important international species in China and will be an effective experimental material supply for future identification of breed-specific genetic markers and for nuclear transplantation and transgenic cloning investigations.
Footnotes
Acknowledgments
This research was supported by the Ministry of Agriculture of China for transgenic research (2008ZX08009-003), the “863” National Major Research Program (2007AA10Z170), and the National Key Technology R&D Program (2006BAD13B08).
Disclosure Statement
No competing financial interests exist.