
Abstract
Recently, there has been considerable interest in developing processing methods that enable storage of cells in a dry state. Most of these studies describe cell viability following processing as a function of the final moisture content reached or the duration of drying. Recently, a cumulative osmotic stress model has been proposed, which takes both final moisture content and duration of drying into consideration in an effort to account for the effects of cumulative processing stresses. The present study demonstrates the applicability of this approach and elucidates a simple mechanistic technique to reduce cumulative osmotic stress during processing. Mouse macrophage cells (J774) were exposed to increasing concentrations of trehalose-containing 0.33× phosphate-buffered saline (PBS) by step-changing the extracellular solution in 2 increments of 0.7 Osm, using only trehalose as the additive solute. Three minutes was provided for equilibration prior to drying in a traditional low-humidity chamber. The data were compared with that of cells dried directly in isotonic 0.2 M trehalose in 0.33× PBS. Following dehydration, cells were rehydrated and viability was assessed 45 min postrehydration using a combination of trypan blue staining for membrane integrity of detached cells and calcein AM–propidium iodide fluorescence assay for live–dead staining of the attached cells. Cells that were preprocessed to higher trehalose concentrations in step changes prior to drying had higher viability scores at comparable final moisture levels when compared with cells dried in iso-osmotic solution, up to a limit of ∼8 Osm, at which point cells processed by both methods approached zero viability.
Introduction
A careful look at the dry preservation literature indicates that a variety of preservation approaches have been used to improve the outcome of dry processing. Most of these preservation efforts center around the objective of creating a glassy environment inside the cell at low moisture contents, by introducing glass-forming sugars, such as trehalose, into the cells. Techniques such as genetic transfection, 6 genetically engineered pore formation, 7 native pore activation, 8 thermal poration, 9 and fluid phase endocytosis 10 have been used to introduce trehalose into the intracellular space. These studies have indicated that although the presence of intracellular trehalose increases the extent of survival of the mammalian cells during drying, the protection offered by these techniques was not enough to preserve cells for a long term in a desiccated state. Cells generally lose viability during processing while still at relatively high moisture contents, well before the cell enters the viscosity regime that can lead to stabilization. Although efforts are underway to find potential strategies to create a glassy environment at higher moisture levels 11 to enable preservation with less dehydration, there remains a need to manage the osmotic stresses that occur during processing for dry preservation.
A variety of different drying techniques have been used to dehydrate cells for storage in the dry state.12–17 Many studies have used environmental drying chambers to desiccate cell samples suspended in droplets18–20 or in thin films.19,21 Spray drying, 22 specialized chamber drying, 16 microwave drying, 17 and vacuum drying 23 have also been used to dry cells. A more complex freeze-drying technique has been used by Wolkers et al. 9 to successfully preserve nonnucleated platelets. Similar approaches with nucleated cells met with limited success. 24 With the exception of freeze-drying approaches, among the different drying techniques used to study dry preservation of mammalian cells, a common theme is that cells are dried indirectly by dehydration of the complete cell suspension. Thus, as the matrix dries, one expects the cells to also become osmotically dehydrated, at least during the initial stages of drying. As drying proceeds, and the matrix becomes more viscous, movement of water from the cell and throughout the matrix becomes diffusion limited and it becomes more difficult to predict the true hydration level of the cell as a function of time. In drying studies of mammalian cells, cell survival or viability is generally reported as a function of a gravimetrically based measure of the entire sample mass (ie, cells, water, and all solutes). The drying time or the rate of drying is rarely considered.
Recently, Chakraborty 25 and Elliott et al. 26 presented a thermodynamics-based cumulative osmotic stress model that attributes cell viability losses in the high moisture content regime of a drying process to a combination of both the time and level of exposure to hyperosmotic solutions. The osmolality of the suspending media was predicted from gravimetric data acquired during drying by assuming that the Van't Hoff factor or degree of dissociation for the dissolved solutes did not change appreciably during the initial phases of drying. As the cumulative osmotic stress model depends on both the extracellular osmolality of the cells and the duration of exposure of the cells to hyperosmotic conditions, reducing the time it takes to reach a given moisture level should increase the proportion of cells that remain viable at that moisture level, as long as the rate of drying does not appreciably exceed the time taken by the cell to volumetrically respond to the change. It is well known that at a given temperature, mammalian cells can withstand only moderate changes in the extracellular osmolality,27–33 a process constrained by the hydraulic conductivity of the cell membrane.31–33 A large change in extracellular osmolality, beyond the tolerable limit of the cell, can cause severe cell injury and generally leads to cell death. 34 Muldrew and McGann 35 have suggested that a large difference in the chemical potential of water between the intra and extracellular environment can irreversibly alter the membrane and ultrastructure of the cells.
The present study investigates a simple mechanistic way to reduce the water content of cellular suspensions during processing for dry preservation while minimizing processing losses. By step-changing the extracellular solution in a manner that does not exceed the critical limits of cell volume change, and/or chemical potential differential, a reduction in the cumulative osmotic stress can be achieved as cells are dehydrated through the high-to-intermediate moisture range of a drying process. The same approach is routinely used to load cryoprotective agents prior to cryopreservation processing for mammalian cells. 36 Although ultimately the solution viscosity limits the extent of dehydration that can be achieved by this approach, in this work we have shown that the utilization of step changes in the early stage of drying can improve the overall survival of cells at a given dehydration level.
Materials and Methods
Cell culture
Mouse macrophage cells (J774.A1) were obtained from American Type Culture Collection. Cultures were maintained at 37°C, 10% CO2–90% air in cell culture media consisting of Dulbecco's modified Eagle's medium (DMEM) with 4.5 g/L glucose (Mediatech), 10% fetal bovine serum (Mediatech), and 1% of 5000 international units (I.U.) penicillin and 5000 μg/mL streptomycin solution (Mediatech). Cells were first cultured in 25-cm2 cell culture T-flasks (Corning) for 3 days. As cells neared confluence they were detached from flasks by scraping and then transferred into 50-mL spinner culture flasks (Wheaton Science Products). All cells used for experiments were taken from established suspension cultures that were maintained at a density of <1 × 106 cells/mL.
Trehalose loading
Macrophage cells have the capacity to take up solutes from the extracellular milieu by fluid phase endocytosis. 37 This native mechanism was used to achieve intracellular loading of trehalose.10,17 Cells were suspended in regular culture media that was supplemented with 50 mM trehalose and then incubated for 18 h in 50-mL spinner culture flasks. Following incubation in trehalose the cells were centrifuged and resuspended in the desired drying solution.
Optimization of step changes
The maximum allowable increase in osmolality that can be tolerated in a single step was determined by exposing the cells to a series of hypertonic trehalose solutions in phosphate-buffered saline (PBS). A cell pellet with 1 × 104 cells was created by centrifugation and then exposed to aliquots of hypertonic trehalose solutions of different osmolalities (0.5, 0.6, 0.8, and 1.17 Osm) in a single step from iso-osmotic conditions. All solutions were prepared from a base of 0.33× PBS. The goal was to determine the maximum allowable step change in extracellular osmolality that did not compromise the membrane integrity. Cell pellets were exposed to different hypertonic conditions for 3 min to allow sufficient time for water transport from the cells. The membrane integrity was then evaluated by resuspending the cell pellet in 0.3 Osm PBS–trehalose solution containing trypan blue. These results were used as the basis for optimizing the next osmolality step change. Cell pellets that had been previously exposed to 0.6 Osm trehalose–PBS solution in 1 step from isotonic condition were again exposed to hypertonic trehalose–PBS solutions having osmolalities of 1.17, 1.8, and 3.14 Osm. After allowing the cells to reequilibrate to the new high osmolality conditions for 3 min, the cell pellet was diluted back to isotonic conditions in 2 steps (suspending the pellet for 3 min in 0.6 Osm trehalose-PBS solution, followed by resuspension in isotonic trehalose-PBS solution containing trypan blue).
Dehydration studies
Cell samples were prepared by plating a droplet of 20 μL volume with a cell density of 5 × 105 cells/mL onto 22-mm2 plastic coverslips (Fisher Scientific). The coverslips were then placed in 35 × 10 mm culture dishes (Corning) and placed in an incubator for 45 min to allow cell attachment. Following the incubation period, the media were gently aspirated and an equal volume of 0.6 Osm drying solution was placed on top of the plated cells. After 3 min the drying solution was carefully removed and 7 μL of 1.17 Osm drying solution was placed on top of the plated cells. This volume is equivalent to the amount of suspending solution that would be present if the suspension was dried to the corresponding solution osmolality by traditional methods. These coverslips were then weighed and placed in a Secador auto-desiccator (Fisher Scientific) for various time periods ranging from 10 to 120 min for drying. Control cells did not undergo any step change in the concentration of the extracellular solution and were placed directly into the auto-desiccator for drying. Samples were weighed again at the end of the drying period. At the end of the drying period, the samples were rehydrated with 100 μL of full-complement DMEM at 37°C and placed in the incubator for a recovery period of 45 min.
Viability assessment following drying
Following the recovery period, samples were removed from the incubator, the extracellular medium was removed, and the number of detached live cells was determined by trypan blue staining. An aliquot of calcein-AM and propidium iodide (PI) solution was added to the cells that remained attached to the coverslip and the samples were again incubated for 5 min at 37°C. Micrographs were collected using an Olympus IX81 motorized inverted research microscope (Olympus America) with fluorescein isothiocyanate and PI fluorocubes. The attached cell viability was determined by counting the live and dead cells on 3 representative images on the same slide. The total viability was estimated by adding the viability scores of both attached and detached cells as in Chakraborty et al. (2008).
17
The detached cell viability (DV) was calculated as follows:
The attached cell viability score (AV) was calculated according to the following formula:
Data analysis
Survival data for cells dried with and without a preprocessing step were plotted as a function of water content (g H2O/g dw) and effective osmolality (Osmol/kg). Effective osmolality of the dried samples was derived from the gravimetric data by using the following equation
26
:
Here, k represents the effective number of osmoles per kg of dry weight. The value of K was experimentally determined from the osmolality measurements of the drying solution (Osm0) and the initial solution moisture content [H2O (kg)/dry weight (kg)]. For further details, please see Elliott et al. (2008).
23
The equation assumes that the Van't Hoff factor determined for initial suspending solutions did not change appreciably as the solutions were dried. The equation also assumes that the moisture content on a dried sample analyzed in dry weight basis is proportional to the inverse of the osmolality of the sample.
26
The limitations of this approximation are presented in more detail in the discussion. The duration of drying was then incorporated as a vector by plotting the survival data as a function of the cumulative osmotic stress, defined previously
26
as a function of both time and osmotic exposure (Fig. 5) according to the following equation:
where ΔOsm′ is the deviation of effective osmolality from isotonic condition at time t.
Results
Optimization of step change
The fraction of viable cells following rehydration from the first step change is shown in Fig. 1. As expected, the number of viable cells decreased as the extent of hypertonicity of the step change increased. The viability following rehydration from the 0.6 Osm/kg trehalose–PBS solution remained very high and it was chosen as first step change for preprocessing of cells prior to traditional drying (Fig. 1).

Fraction of viable cells following rehydration after cell samples were exposed to different hypertonic conditions in a single step. The viability following rehydration with 0.6 Osm/kg trehalose–PBS solution remained higher than the cell samples that were exposed to 0.8 or 1.17 Osm/kg trehalose–PBS solution (n = 5). PBS, phosphate-buffered saline.
Figure 2 shows the second step of the stepwise hyperosmotic exposure experiment. A hyperosmotic exposure from 0.6 to 1.17 Osm yielded maximum viability upon rehydration to isotonic conditions. No further step changes were evaluated because of the practical limits imposed by the increasing viscosity of the solutions. Using the free-volume approach of linearized William–Landel–Ferry equation,38,39 viscosity of 1.17 Osm trehalose–PBS solution at 22°C was estimated to be ∼3.7 cP. This concentration of trehalose–PBS drying solution was used to dry cells to lower moisture contents in a low-humidity desiccation chamber.
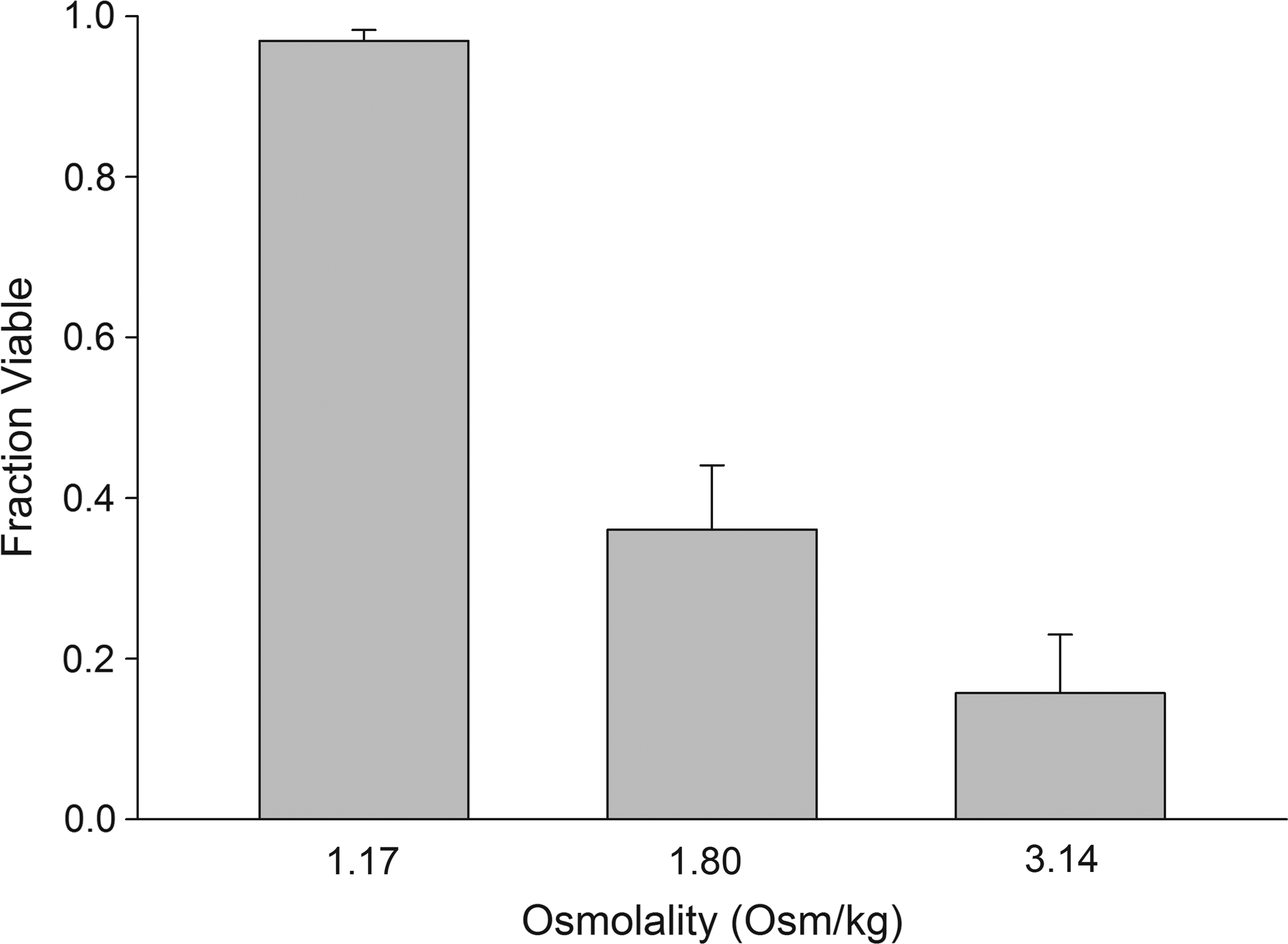
Fraction of viable cells following rehydration after cells were exposed to hyperosmotic conditions in 2 steps. The samples were exposed to 0.6 Osm/kg trehalose–phosphate-buffered saline solution in first step for 3 min, followed by exposure to 1.17, 1.8, and 3.14 Osm/kg for 3 min before returning to iso-osmotic condition. A hypertonic exposure from 0.6 to 1.17 Osm yielded maximum viability upon rehydration to isotonic conditions (n = 5).
Viability of cell samples
Immediately after drying for the desired time, samples were rehydrated with 100 μL of full-complement DMEM at 37°C and placed in the incubator for a recovery period of 45 min. The total viability measured following the 45-min recovery period was plotted in terms of grams of water per gram of dry weight (Fig. 3). To illustrate the differences in viability trends between the preprocessed cells dried from 1.17 Osm trehalose solution and cells dried from isotonic 0.308 trehalose solution, standard nonlinear curve fitting analysis was performed. Based on the R 2 values, logistic dose–response curves were found to be the best fit for both the datasets (Fig. 3).

Comparison of viability responses of J774 mouse macrophage cell samples that were dried after step-changing the extracellular osmolality to 1.17 Osm/kg and samples that were dried from isotonic condition (0.308 Osm/kg). The fractional viability was plotted against grams of water per gram of dry weight of the sample (g H2O/g dw). A nonlinear regression analysis of the 2 datasets were performed and logistic dose–response curves (
In this model the value of osmotic exposure that yields 50% viability is defined as the critical level (CL50). The CL50 value was statistically different for the 2 datasets, indicating a fundamental difference in the trends of the 2 datasets. Although 50% of the pretreated cell population remained viable at the moisture level of 0.65 ± 0.014 g of water per gram of dry weight, for cell populations dried in isotonic trehalose solution this cell viability level reached 1.19 ± 0.008 g of water per gram of dry weight. Processing losses are thus significantly higher when cells are dried from isotonic conditions.
The gravimetric data were then converted to osmolality using the relations described by Chakraborty 25 and Elliott et al., 26 and the viability response was plotted against this variable. The preprocessed cell samples indicated higher levels of viability at the same osmolality level (Fig. 4). Both datasets were fitted with logarithmic dose–response curves and the analysis shows that there is significant difference between the 2 processing conditions. For pretreated cells the CL50 value was 5.75 ± 0.54 Osm, whereas for cell samples dried in isotonic trehalose solution the CL50 value was 3.62 ± 0.94 Osm. The viability of both the cell samples decreased rapidly beyond the extracellular osmolality of 8 Osm/kg, suggesting the existence of a barrier to further drying with current protectants and drying approaches.
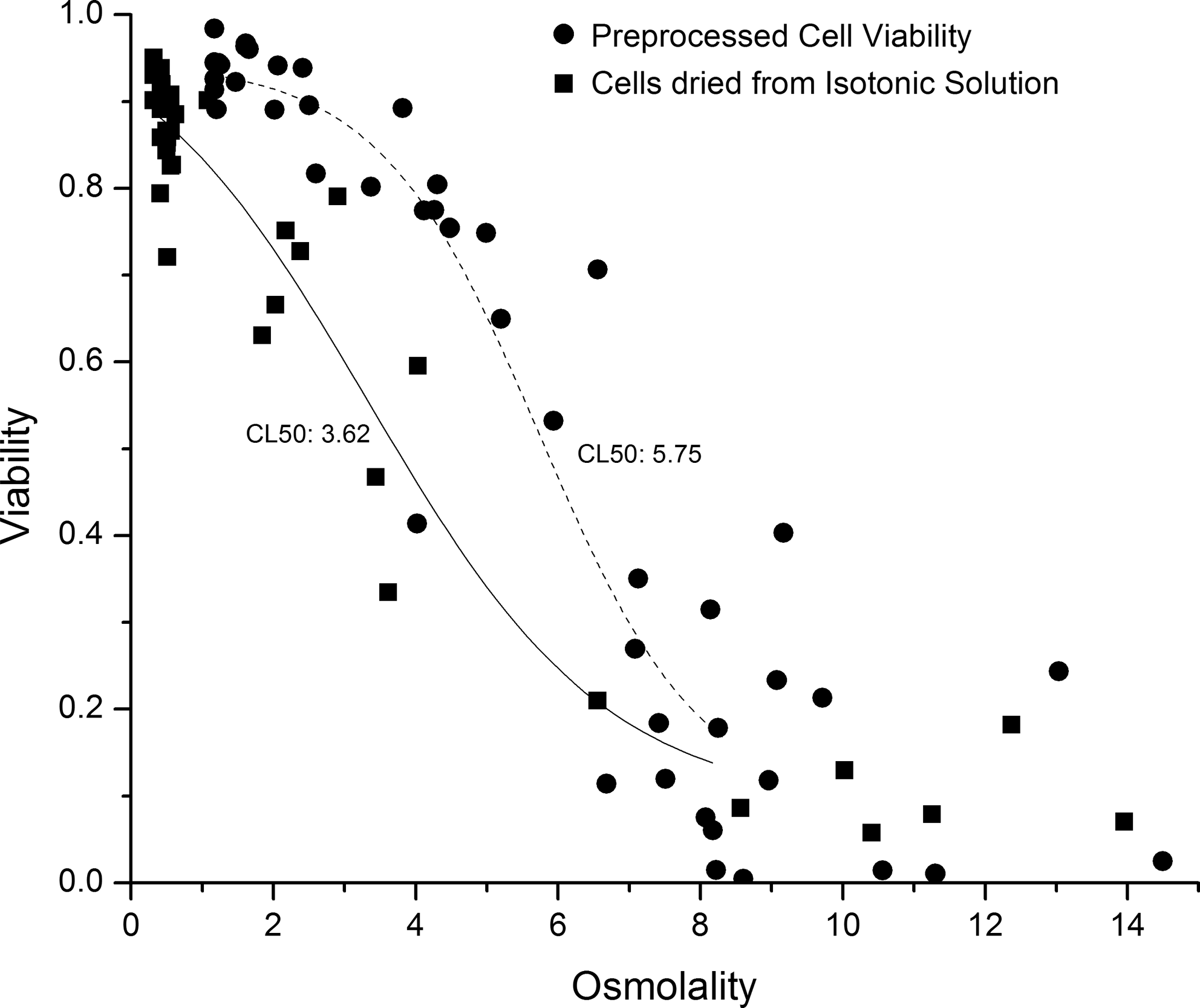
Comparison of viability responses of J774 mouse macrophage cell samples that were dried after step-changing the extracellular osmolality to 1.17 Osm/kg and samples that were dried from isotonic condition (0.308 Osm/kg). The fractional viability was plotted against estimated osmolality. Both the datasets were fitted with logarithmic dose–response curves (
Survival data were also plotted as a function of cumulative osmotic stress, which include both the level of chemical perturbation from equilibrium (extracellular osmolality) and the time component of drying. The viability trend (Fig. 5) was sigmoidal in nature for both control and preprocessed samples. As predicted by Elliott et al., 26 there is no statistical difference between the viability response of the 2 different cell samples, when plotted as a function of cumulative osmotic stress. These data suggest that the injury to mouse macrophage cells is physicochemical in nature in the high-moisture regime and is independent of the means used to remove moisture from the suspension. It is important to note that representing the viability response as a function of cumulative osmotic stress also resulted in decreased scatter in the data compared with using just the final moisture content as the independent variable, suggesting that drying time is an important contributor to the overall injury.
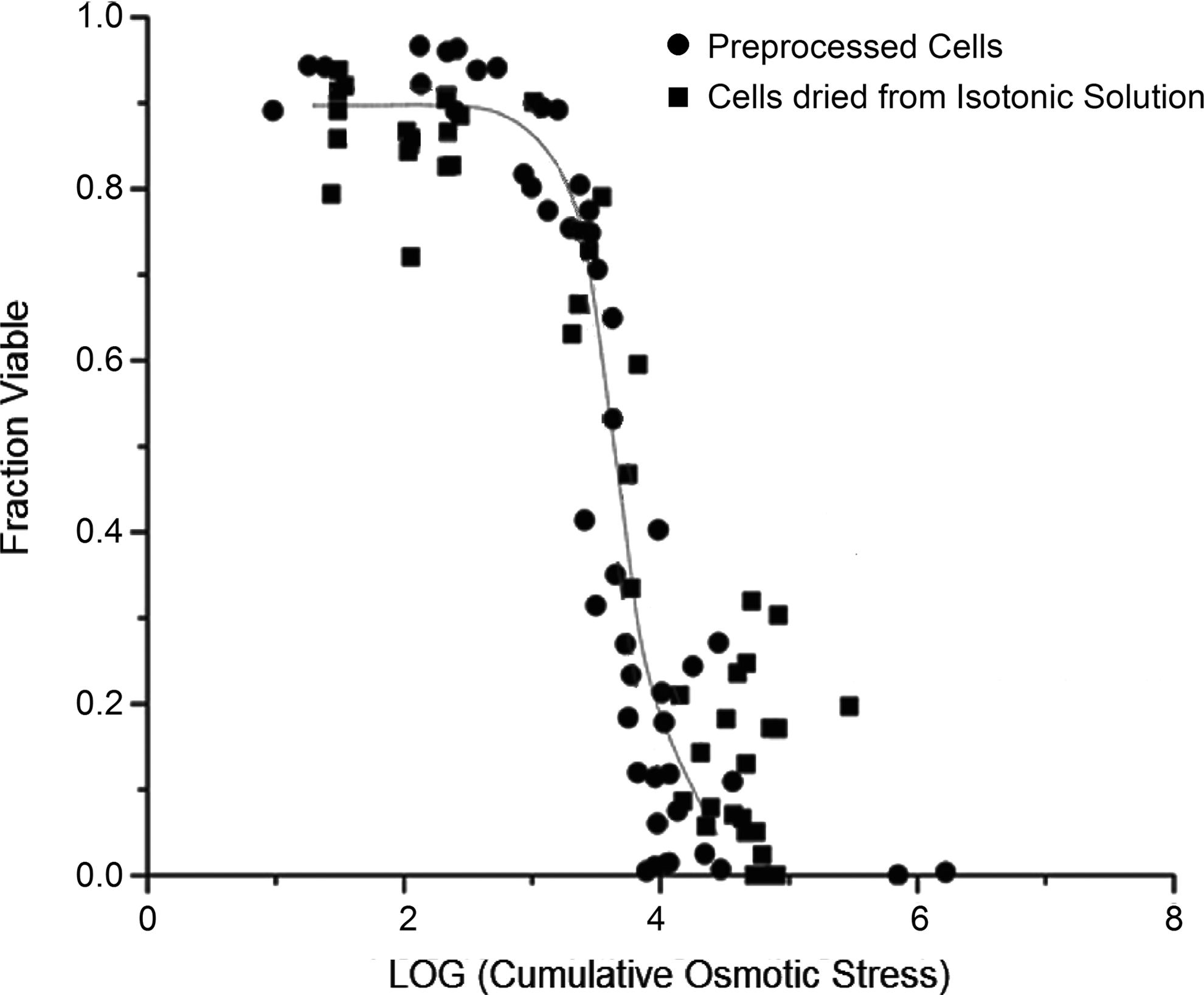
Fractional viability of J774 cells expressed as a function of the logarithm of cumulative osmotic stress during drying. Circles represent the viability of J774 cells that were dried after step-changing the extracellular osmolality to 1.17 Osm/kg. Squares represent the viability of cell samples that were dried from isotonic conditions. Both the datasets were combined and fitted to a double exponential function: 0.0759 + 0.8068 exp (–exp ((3.9214 – x)/0.4315)); R2 = 0.960.
Discussion
Dry storage of mammalian cells at ambient conditions remains an elusive goal. The progressive loss of viability as cells are dehydrated for storage in a dry state has prevented the attainment of a specimen suitable for long-term dry storage at temperatures approaching ambient. Further advances will be needed to minimize processing losses such that noncryogenic storage can become a reality.
The present study has demonstrated that higher viabilities can be achieved at lower moisture levels if rational osmolality changes are made in the high moisture range of drying. The total viability of mouse macrophage cells, when expressed on a g H2O/g dw basis (Fig. 3), was observed to be higher for preprocessed cells, over a range of moisture levels, compared with cells dried from isotonic conditions. The difference between groups was equally apparent when the viability data were expressed in terms of osmolality (Fig. 4). Effective osmolality was derived from the gravimetric data by assuming that the Van't Hoff factors of the solutes did not change appreciably with concentration. 26 This is a reasonable approximation in the high moisture content regime where most of the biological response is observed, but this assumption is expected to produce greater deviations from the true osmolality as the solution becomes increasingly hypertonic. In the low moisture content regime, the diminishing diffusion rate of water in high-viscosity solutions will dominate the evolution of local chemical gradients, possibly requiring the development of an entirely different stress model in this regime. Nevertheless, the cumulative osmotic stress model provided a good framework to evaluate the benefits of including preprocessing steps in the high moisture content range.
The step change approach enables a significant reduction in the time that cells are exposed to hypertonic stresses in the isotonic to 1.17 Osm range, thereby reducing the overall cumulative osmotic stress exposure as a cell is processed to a given moisture level. The expected effect of reducing the accumulation of this stress is to be able to maintain a higher surviving fracture at a progressively drier state. The data revealed that this was indeed the case. Samples dried directly from isotonic solution reached a 50% loss of cell viability at an extracellular concentration of ∼4 Osm/kg, whereas this loss was reached at ∼6 Osm/kg for samples that were preprocessed. This result is significant because low moisture levels are essential to achieve the reduction in molecular mobility necessary for long-term storage. If progressively higher fractions of surviving populations can be achieved at lower moisture contents, preservation in a dry state becomes more tractable. In both groups the viability approached zero at the same moisture content (∼8 Osm/kg), suggesting the possibility of a critical tolerable moisture loss for the given conditions, which would be a barrier to further reduce the molecular mobility using drying approaches. When the viability data were plotted in terms of cumulative osmotic stress, a metric that also incorporates the time taken to dry to a specific moisture level, the data followed a single master curve. The fact that the different treatments follow the same master curve when presented in terms of cumulative osmotic stress would suggest that the variation in the 2 processing approaches is well represented by the elements included in the model (hypertonicity, time of exposure), and that these elements are the predominant stress vectors in the high moisture content regime.
As an illustrative example, traditional passive drying of samples in a low-humidity chamber required ∼25 min to reach an estimated extracellular osmolality of 1.17 Osm. By step-changing the osmolality to 1.17 Osm over a period of ∼6 min before traditional drying the cumulative osmotic stress was reduced by 1.92 units. The model predicts that this reduction in the time to dry cells to intermediate moisture levels should produce an increase in viability of the cell samples by 24.7% at an extracellular osmolality of 2 Osm. The results (Fig. 4) indicated that the viability of cells increased by ∼25%, consistent with what the model predicted.
It is important to keep in mind that the physicochemical stresses responsible for a reduction in cell viability are probably more complex than a 2-factor metric described by the cumulative osmotic stress model. Other studies provide evidence of solute-specific dehydration effects, 15 and drying temperature will almost certainly play a role at some level. Nevertheless, this cumulative stress metric, which includes both time and level of osmotic stress, offers a tool to unravel the nature of injury during drying and is helpful in designing methodologies for dehydration processing of mammalian cells with improved outcomes.
Unfortunately, the step change approach of preprocessing cells is ultimately limited by the practicality of solution manipulation. As the tonicity of the trehalose solutions increase, so does the viscosity, and hence quick changes in extracellular concentrations become difficult. Further, these high tonicities represent supersaturated solutions that require considerable effort to prepare and maintain. Beyond this practical range of step-change dehydration, samples will probably require rapid drying in the intermediate-to-low moisture content regime. Fast active drying techniques such as the microwave drying approach developed by our group 17 may be useful. Active drying technique can overcome the limitations of dehydration processing of cells by limiting the time of exposure to hyperosmotic conditions and preventing a build-up of osmotic stress. In this regard, it is interesting to note that many anhydrobiotic organisms must be dried slowly, to provide the needed time to adapt to the desiccated state by synthesizing anhydroprotectants and rearranging intracellular molecules that are essential for desiccation tolerance.40–42 Because mammalian cells are artificially imbibed with anhydroprotectants such as trehalose prior to drying, slow drying to enable biomolecular adaptations may not be a requirement. However, the possibility of drying “too fast” does exist. By exposing the cells to higher osmolality conditions in step changes, the dehydration rate was effectively increased, which had an overall beneficial effect. However, a systematic study of drying rate was not performed in this particular study. An examination of prior art can be instructive here. Using the cumulative osmotic stress model that inspired the present study, Liang and Sun 43 observed that the rate of dehydration greatly affected desiccation tolerance of seeds that are sensitive to moisture loss (recalcitrant seeds). Their studies demonstrated that there was an intermediate “optimal” drying rate for these seeds, and it was proposed that physical stresses were dominating at the highest drying rate, whereas physicochemical stresses were dominating at the lowest drying rates. There are considerable physiological differences between mammalian cells and seeds, not the least of which is the absence of a cell wall in mammalian cells. Despite that, there is some likelihood that if water is removed too rapidly, the evolution of shear stresses in the region of kinetically trapped cells can result in direct mechanical damage to cells. Further studies are needed to understand the injury mechanisms in the low moisture range and to determine if there is indeed an optimal dehydration rate for mammalian cells, as there exists for seeds.
It is also important to note that the processing steps described in the present study are not intended to be a prescriptive methodology for dry preservation of mammalian cells. The trehalose-loading approach used in this study was based on best known practices for this cell type in our laboratory, but continued advances are needed to determine the optimal level of intracellular protectants, as well as the best approach for achieving suitable concentrations throughout the cell, with minimal stress to the cell membrane. Based on our previous experience with this cell type, we have observed that membrane integrity assays performed after a 45-min postrehydration recovery period are in good agreement with apoptosis assays and metabolic assays performed over a longer time period. 17 Further work will be needed to establish the predictive value of this model for other cell types and to confirm that long-term survival and functionality are indeed preserved for these cell populations.
Studies on other cell types suggest that achieving adequate levels of intracellular stress proteins may also be an important element in attaining an overall solution that yields long-term survival in a dry state. 19 If the strategy to achieve reversible mammalian cell desiccation requires modeling the behavior of anhydrobiotic organisms, this would suggest that using chemical agents that can downregulate cellular metabolism will be another important element of a comprehensive drying formulation and process. 44 Independent of the exact drying solution composition that ultimately emerges, osmotic stresses in the high moisture content regime will need to be carefully managed.
Conclusion
In summary, the present study illustrates a simple mechanistic way in which the processing stresses involved in dehydrating cells for anhydrous preservation can be reduced, thus improving the likelihood of achieving suitable surviving populations for storage in a dry state. This processing technique alone will not be able to solve all the bottlenecks that appear in the path toward successful dry preservation of mammalian cells, but when applied in conjunction with other protective strategies it can improve the overall outcome.
In the present study we demonstrated that the model, which includes the estimated average osmolality of the dried sample (based on gravimetric data) and the total drying time, can be used to predict the viability of cells in dehydrated samples and used to create better drying protocols for dry preservation of mammalian cells. The analysis of drying outcome as a function of cumulative osmotic stress provided additional insight into the nature of cell injury induced by dehydration, compared with an interpretation based on end moisture content alone. Using the cumulative osmotic stress model we demonstrated that cell injury can be reduced by starting the passive dehydration process from an advanced drying condition that can be achieved by step changes in the extracellular solution composition. The experimental results confirmed that in mouse macrophage cells, cumulative osmotic stress is an important injury mechanism during drying. The model was able to predict injury as well as the effect of modulating the drying rate. Changes in processing conditions that can minimize the amount of time spent by the cells at intermediate osmolality levels were shown to have a very significant effect on the outcome. For example, simple process changes such as step-changing the extracellular solution composition prior to drying in a low-humidity chamber can reduce cumulative osmotic stress and improve the outcome of dry processing of mammalian cells.
Footnotes
Acknowledgment
This project was supported by a UNC Charlotte Bonnie Cone Award to G.D.E. (NSF Institutional Award No. SBE0548401).
Disclosure Statement
No competing financial interests exist.