
Abstract
Tissue banks represent essential resources and platforms for biomedical research serving basic, translational, and clinical research projects. In this article, we describe 2 models of biobanking and tissue preservation with different approaches and aims. Archive tissue biobanking is described here as a resource of residual pathology tissues for translational research, which represents the huge clinical heterogeneity. In this context, managing of tissues and RNA quality in archive tissue are discussed. The other model of tissue biobanking is referred to as xenograft tissue banking, which represents an alternative method for obtaining large amounts of tissue, over an indefinite period, in so far as the tumor can be transferred in vivo over generations, maintaining the histological and genetic particularities. A description of the method and examples of the application are given with particular emphasis on sarcomas (Ewing's sarcoma/primitive neuroectodermal sarcoma, synovial sarcomas, and rhabdomyosarcomas) and early stages of tumor angiogenesis in sarcomas.
Introduction
The Archive as a Pathology Biobank
Tissue banks represent essential resources and platforms for biomedical research serving basic, translational, and clinical research projects by providing human tissues and their respective derivatives. 2 A requisite for research tissue banks is to provide high-quality materials, which are the consequence of optimized procedures for acquisition, storage, documentation of specimens, expert evaluation, and standardization in the material transfer procedures. All these aspects in fresh-frozen tissue biobanks are costly because of the technological requirements to obtain and process frozen specimens. Moreover, there is a need for tissue documentation and identification, which has to be fulfilled by professionals. Pathologists have professional experience and expertise in sampling, storage, diagnostic evaluation, and distribution of human tissue samples. With such requisites, the archives of the pathology departments could represent an affordable and highly characterized type of human tissue biobank. In hospitals worldwide, histopathological analyses are performed on formalin-fixed paraffin-embedded (FFPE) tissues, namely the AT. These, as a consequence, represent all the routinely treated tissues for diagnostic or surgical reasons in any hospital. After few sections are cut for the histopathology examination, all these tissue blocks are stored in archives for years or even many decades, depending on the local legislation. They are related to the clinical records, and for this reason, they have a high level of clinical information available. Moreover, they represent the actual clinical material and the real clinical and epidemiological situation at the population-based level. AT specimens surely have some limitations in comparison to fresh-frozen samples because of the chemical modifications caused by formaldehyde on nucleic acids and proteins. 3 However, in comparison to biobanks of fresh-frozen tissues, pathology biobanks have 3 main advantages: (i) low costs, as they are already present in every department of pathology; (ii) tissues are morphologically well defined and referred to a specific clinical diagnosis, performed by experienced pathologists; (iii) the huge number of available samples with even rare lesions and long follow-up.
A pathology archive could change from biorepository to biobank just by anonymizing the samples (Fig. 1), before those samples and the related clinical information are exported for research purposes. The most suitable way is to modify with pseudo-anonymization (double coding), wherein only the case study will be used in a specific project.

The concept of tissue archives as a biobank. The pathology archive can change from biorepository to biobank function simply by anonymizing the samples. Anonymized samples could be used in large-scale validation studies using both molecular and morphological approaches.
In this context, the expertise of pathologists is not limited to the provision and choice of tissues, but they have also been the first to perform molecular analysis in ATs.4–10 For this reason, a group of experienced molecular pathologists worked together on a European project “Archive Tissues: Improving Molecular Medicine Research and Clinical Practice–IMPACTS” (www.impactsnetwork.eu) to validate and standardize these methods, which are going to be published in a comprehensive handbook. 11 The aforementioned group of pathologists is now developing a Pan-European AT biobanking network (PAT-B Network). This will allow European pathologists to participate in multicenter projects on a voluntary and collaborative basis. The involvement of pathologists in these projects will help spread the experience and diffusion of molecular methods in pathology diagnostics, in the same way as it happened in the past with the direct experience of pathologists using immunohistochemistry. PAT-B Network is based on free and collaborative participation on specific projects. The participants will be considered as full collaborators of the project.
The participants in the PAT-B Network can be also involved in other localized or countrywide biobanking networks. This will not affect the activity performed within PAT-B. The PAT-B Network was developed only to perform very large international projects on clinical biomarker validation.
Even if formalin is banned, ATs will be present for histological examination. We could expect the use of alternative alcohol-based fixatives for better preservation of both nucleic acids and proteins.7,9,12–16 In this case the banning of formalin will improve AT biobanking, both in terms of toxicity and biomolecular preservation.
RNA Quality in AT
Nucleic acids, and specifically RNA, are modified by formalin during the fixation process. Nevertheless, differences in tissue preservation could also affect the quality of RNA. Before being analyzed, human tissues undergo essential treatments, which fall under the designation of “preanalytical treatment.” These are represented in detail by the surgical removal of tissue, grossing (in the pathology laboratory), fixation, and paraffin embedding.
Although grossing, paraffin embedding, and tissue banking are well defined in literature, there is no standardized definition of the time interval between surgical removal to tissue grossing and fixation. 17 Preanalytical treatment of tissues is one of the most variable and debated issues in molecular analysis. Information about the time between ablation of tissues and tissue fixation or the time and the temperature of fixation procedures is hardly ever available. The major problem derived from the preanalytical time interval is connected to the hypoxia conditions of the tissues, which could occur in surgical specimens, especially of larger size. Thirty minutes is conventionally considered the limit for a conservative treatment of the tissues, before fixation or freezing. 17 This time interval could be related to gene expression changes.18–20 So far, preanalytical time interval, which is comprehensive of warm ischemia in the surgical theater and the time required for grossing and fixation in the surgical pathology laboratories, is overdue to be standardized and represents the major variable affecting the quality of biomolecules, especially RNA. To obtain the highest level of standardization in tissue processing, the policy experienced by Bussolati et al. was to transport specimens under vacuum 21 and to use a well-defined time of fixation (from 5 h, for relatively small specimens, up to 24 h, for large specimens). It is well known that the longer the fixation the lower will be the quality of RNA. 3 Tissues fixed for 12–24 h have presented higher preservation of biomolecules in comparison to longer fixation.22–24 The reaction between formalin and nucleotide monomers has been shown to progress in 2 steps: the first is represented by the addition of a formaldehyde group to a base on a form of N-methylol group (N-CH2-OH), and the second is the slower electrophilic attack of N-methylol group on an amino base to form the methylene bridge between 2 amino groups. 3 However, nucleic acids with mono-adducts (N-CH2-OH) could be demodified and analyzed. 3 In our experience, despite the aforementioned problems, RNA is amenable to real-time polymerase chain reaction (RT-PCR) analyses. Different protocols, from the use of homemade solutions to commercial kits, have been developed for the extraction of this nucleic acid from ATs, and in our experience, they are all amenable to conventional RT-PCR. 5 In our experience, the use of commercial kits, based upon silica column purification of extracts, provides the best RNA quality assessed by RT-PCR in comparison to homemade protocols and commercial monophasic solutions, and these methods are faster. 5 Nevertheless, the use of colon-based commercial kits is far from being a way to standardize molecular methods in AT. Small adjustments to the manufacturer's instructions (eg, proteinase K digestion) may cause interlaboratory variability in both RNA quantity and quality, demonstrating that different results could be obtained by different groups using the same commercial kits. 5 An important parameter requested to standardize methods is surely the assessment of RNA quality in ATs. The use of capillary electrophoresis (eg, Bioanalyzer or Experion) is not predictive of the utility of RNA samples isolated from ATs in molecular analyses, because the chemical modification of RNA by formaldehyde is not resolved by electrophoretic assays.5,7 The best and most affordable methods to evaluate RNA quality in AT is the use of RT-PCR, both endpoint or real-time, by amplifying different fragments of increasing length of housekeeping genes.7,25
Translational Research Using AT
The findings in molecular biology portend a new era in the treatment and prevention of human diseases. A great amount of information has been collected on cancers, but the translation of these discoveries into clinical practice represents a bottleneck. The availability of tissues is one of the major necessities in molecular research in many fields, especially in oncology. This translational research is performed to make therapy more specific and to avoid deleterious side effects. Pathologists are already the tissue providers to the biobanks of fresh-frozen tissues, but this is not the only way in which they can contribute, because they have 2 other basic features. The first is the ability to recognize thousands of different lesions at the morphological level, which only with the help of pathology itself can be correctly defined and microdissected to obtain reproducible research results. The second is the possibility for pathologists to have huge archives of fixed and paraffin-embedded tissues, clinically annotated and with any lesion, even the rarest, which now can be analyzed at the molecular level. 11 These are the only tissues available for most of the patients for any further molecular diagnosis.
There are methods available for molecular research at the DNA and RNA level in paraffin-embedded tissues8,26–30 and also at the protein level, not only by immunohistochemistry but even by western blot and protein arrays.31,32 By the use of AT, it is possible to analyze a huge amount of human tissues with molecular techniques and integrate the experimental data with the clinical records. In our opinion, this is the fastest way to translate the findings of basic research into patient care. This is already a reality and the demonstration is that more than 9 studies have been published from 2006 to date with the keywords “molecular translational research in FFPE” (retrieved from PubMed).
We hope that in the future pathologists will play a major role in the process of biobanking and molecular translational research, accelerating in this way also the clinical application.
Nude Mice Xenografts of Human Tumors: A Model for the Analysis of Tumoral Biobanking
Human malignant tumors can be implanted in immunodeficient mice, where they may grow as xenografts, supported by murine stroma33,34 (Fig. 2). Once established, xenograft lines may be grown indefinitely as heterogeneous tissue better resembling the original tumors than in vitro grown cell lines. Such lines are very useful as cancer models for preclinical experiments35–37 ; however, some properties may not correctly represent the original tumors, as in the case of interactions with the host stroma as well as with the new immune system of the mouse. In addition, a xenograft is established from a small piece of tissue and may not always be well representative of the whole tumor, in which heterogeneity may occur. 38 Further, the selection processes may differ in a mouse host, and new gene aberrations may appear upon passage. Even so, this remains a technique that provides significant added value to the morphological and biological study of human tumors.
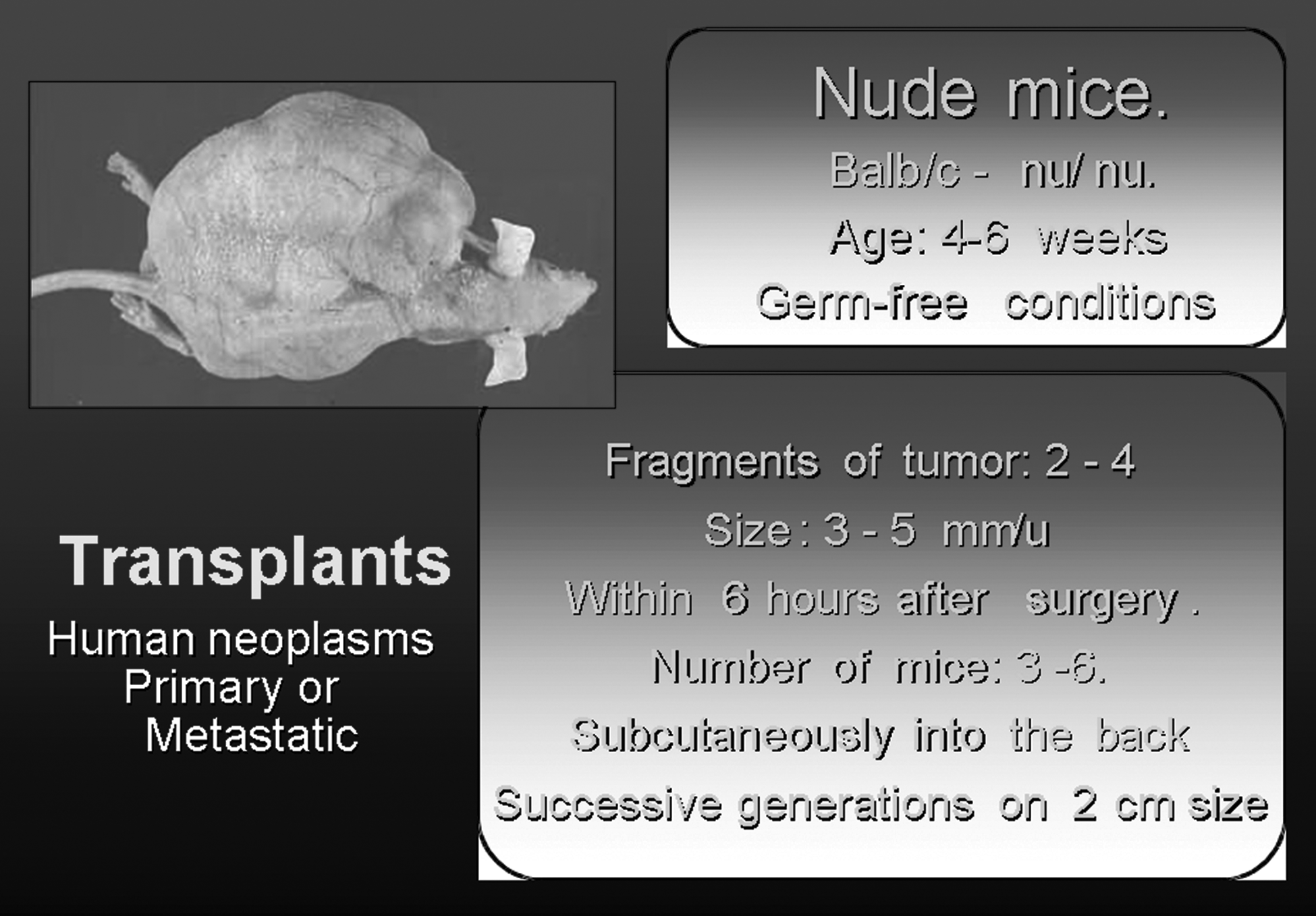
Model for xenografting tumors.
Xenografts may also be indirectly established by injection of in vitro cell lines into mice. However, such lines are less useful as preclinical models 39 as specific mutations may be induced, 40 their programming may be irreversibly changed during in vitro culture, 41 and their response to therapy frequently differs from the patients' one. 39 Therefore, it is important to know how similar xenografts are to their tumor of origin and how faithfully their genomes are maintained upon passage in a xenobiotic host. Even when a cancer cell is well representative of its tumor of origin, patterns of gene expression and signaling are quite dynamic and would be heavily influenced by the growth pattern, vascularization, interactions with and signaling from host stroma cells, and the reduced or absent interactions with the immune system. However, it has been shown that expression profiles of nonmatched sarcoma xenografts cluster together with patient samples of the same tumor type, suggesting good representations of the relevant phenotypes, although minor but critical features may be altered.
Tumor progression is associated with a higher clinical malignancy and, in many cases, with patient's death. Causes of this tumor progression are numerous and not well known at present. In vivo and in vitro models for the study of progression are available (cell cultures, animal models, and DNA microchips). Nude mice xenografts of human tumors provide an interesting tool for the study of tumor evolution and progression in human sarcomas, because they mimic, in an experimental model, what might occur in humans.33,35,36,38,42
For the last 30 years, our group has been building up a systematic transfer of sarcomas into nude mice, mainly oriented toward the analysis of tumor progression and dedifferentiation.42–58 The actual feasibility of archival samples in formalin fixed and paraffin embedded tissues (FFPET) including tissue microarrays in tumor types different from soft tissue sarcoma is even more extensive, because the incidence of breast, colon, lung, and prostate cancers is higher. However, regarding xenograft models in these tumors, it seems that carcinomas, lymphomas, and melanomas did not grow very fast in nude mice when compared with sarcomas. The main study focus uses well-established carcinoma/lymphomas or melanoma cell lines to obtain fresh material after nude mice inoculation.
At present in our Institution, 502 tumors have been transferred and followed for different periods of time (up to 3 years in some cases). This has given us the opportunity to pursue a number of histological, electron microscopic, genetic, and molecular biological reorganizations of a given tumor that could be related with the clinical tumor progression of the patient. Herewith, we present several cases in which nude mice xenografts of the original neoplasm or of their metastasis have been used, allowing us to follow fundamental changes in their structure and their genetic configuration and opening up new insights into their biology. In addition, we have produced an in vivo model for the study of angiogenesis and vasculogenesis, which display a direct correlation between growth rate and intratumoral neovascularization. 47
Models of Tumor Xenografting
Ewing family tumors (EFT) are a subset of pediatric bone tumors, which include Askin tumors, Ewing's sarcoma (ES), and peripheral primitive neuroectodermal sarcoma (pPNET).59–63 We have xenografted 57 cases of small round cell tumors with 23 positive takes (40.3%). Transplantation of the ES/pPNET neoplasms is followed by loss of common structural patterns mainly pseudorossettes and cells adopting a more blastomatous immature picture. Nevertheless, in some cases, the neuroectodermal character with neurosecretory granules and cell processes (dendritic like) persists. Neocapillarization occurs with detection of intratumoral vasculogenesis as well as vascular co-option and capillary mimicry made by tumor cells.
Specific cytogenetic rearrangements are present, with the resultant fusion genes involving EWS gene and members of the E-twenty six (ETS) family of transcription factors that, from a clinical point of view, provide a valuable diagnostic and prognostic marker. Ewing sarcoma breakpoint region (EWS)-ETS rearrangements are implicated in generating malignant transformation of EFT, but the presence of additional genetic alterations must be considered in their pathogenesis.52,59–63 These genetic alterations might not necessarily be tumor specific but may display interindividual variations that could account for differences in EFT phenotype as well as in clinical behavior, differentiation capacity, invasive potential, and treatment resistance. Among these secondary alterations, deregulation of the G1 to S-phase transition key genes plays an important role in the tumor pathogenesis of the EFT.49,61
In our collection of EFT, we have some examples where alterations in 9p21 locus are clearly implicated in the tumor progression. Particularly, in a case with available samples of primary tumor (PT), xenotransplants from PT, recurrence, metastasis, and xenotransplants from metastasis, we observed a loss of expression and homozygous deletion of those genes codified by the 9p21 locus (p16INK4A, p14ARF y p15INK4B). 49 These results show that deletion of the 9p21 locus was clearly manifest during tumor progression and that it was already present in the primary sample but only affecting a small tumor cell population. We reached this conclusion because homozygous deletion of 9p21 was detected in the xenotransplants from the metastasis and PT as well as in the recurrence and metastasis stage. 49
Other cytogenetic numerical changes have also been observed in this case by means of comparative genomic hybridization (CGH) and fluorescence in situ hybridization. These changes include an increase in the copy numbers of chromosomes 8 and 12 during tumor progression.49,52
Synovial sarcomas (SS) are malignant neoplasms of mesenchymal origin displaying a variety of differentiation characteristics. The case of SS constitutes an exception within this context because of the presence of a double epithelial and mesenchymal element, both malignant and simultaneously appearing within the same tumor fields in close continuity.64–68 Based on these structures, 2 main categories have been described, not only histologically but also confirmed with electron microscopy and immunohistochemistry. The SS presents a biphasic type, with an epithelial and spindle cell component, and a monophasic type, exclusively constituted by the spindle cell component of the former.56–58 Our experience in nude mice xenotransplants is of 20 xenotransplants with 13 positive takes (65%), in which both monophasic and biphasic SS are involved.
The ultrastructure confirms the biphasic pattern of this neoplasm. One or two cell layers of cylindrical cells that coexist in continuity with the cubical or flattened ones cover to the tubular glands.56–59,63
In addition, solid clusters of epithelial nonsecretory cells are found surrounded by an incomplete lamina densa material, similar to that seen in the glandular compartment.59,63
The poorly differentiated small cell variant of SS may resemble ES. At EM level, small spindle cells look like blastemic or immature mesenchymal cells with an active functional capacity but clearly differ from fibroblasts. They possess short, elongated, sharp, nonbranching cell processes, more or less densely packed in a stroma filled with flocculent material. The cytoplasm is dense with isolated vacuoles and lipid droplets. Some spindle cells produce secretory material within the cytoplasm. No transition occurs between the large spindle cells and the small ones.59,63
In the nude xenografted model, of the 13 cases that presented positivity, tumor transfer was maintained for at least 6 passages. The histology of the tumor was followed in both types of neoplasm: monophasic and biphasic SS. Particular mention should be made regarding the biphasic SS that displayed glandular structures in the original neoplasm. After the implant into the nude mice, the glandular component progressively disappeared and was substituted by the solid epithelioid pattern, which appeared intermingled with the spindle cells. The epithelial glandular pattern seems to be labile and does not maintain its original configuration after xenotransplant. On the other hand, the epithelioid cells become dominant acquiring a sarcomatous appearance.56–58
Comparable considerations are valid for the monophasic SS that kept its original configuration without any major changes. A case of small round cell SS of undifferentiated type presented a major dedifferentiation losing the entire spindle cell component and mimicking a PNET neoplasm.56–58
Cytogenetically, a reciprocal translocation has been described, the t(X;18)(p11.2; q11.2) is present in almost 100% of SS cases, being considered a specific marker, and from the oncological point of view the primary cytogenetic abnormality for this type of sarcoma.52,56–58,64–68 This translocation results in fusion genes that might be involved in their tumorigenesis and in their clinical outcome.
Unlike ES/pPNET, no alterations of the cell cycle were observed. However, other gene alterations have been described, such as mutations of the PDGFRa that could have important prognostic or therapeutic implications.
Rhabdomyosarcoma (RMS) is the most frequent soft tissue malignancy in children and adolescents. This tumor is derived from the primitive mesenchyme, with a tendency toward myogenesis. Two major types are known, embryonal (ERMS) and alveolar (ARMS). We have been following for long periods 32 transplants with 14 positive cases of both ERMS and solid/ARMS.69–74 Electron microscopy has been useful in complementing the diagnosis of these tumors, particularly when dealing with the undifferentiated forms: a myoblastic differentiation appears with myosin filaments and ribosome complexes, sarcomeric-like figures with focal densities, and arrays of thick and thin filaments distributed in parallel bands. Very poorly undifferentiated RMS may express only nonspecific myofilaments oriented randomly, or the so-called leptomeric fibrils as complexes of loosely organized intermediated filaments of 10 nm in diameter, perpendicularly located to dense amorphic material.59,63,69–74
A cytogenetic and molecular analysis of 7 cases confirmed that ARMS and ERMS are entities with different genetic characteristics: the t(2;13) and the chimeric transcripts originating from the t(2;13) and t(1;13) were found only in ARMS assessing the specific association of these translocations with that histological subtype. Further, MYCN amplification was found only in ARMS. Deletion of the 9p21 genes was observed only in a spindle-cell ERMS.69–74
Vascular neogenesis analysis in vivo xenografts
We also used the nude mice xenotransplant model to study the vascular neogenesis of soft tissue tumors. Implants, 0.3–0.4 cm in size, of a ES, an osteosarcoma (OS) and a fibrosarcoma were analyzed. The study was successively followed for 24, 48, and 72 h; 1, 2, 3, and 4 weeks. Hematoxylin and eosin, periodic acid-Schiff's (PAS) staining, and immunohistochemistry for angiogenic factors were complemented with electron microscopy. These models provide excellent examples of all the systems that are involved in the recapillarization of a sarcoma. 47 After 24–48 h, the implants over-expressed several angiogenic factors. At this stage, the tumors initiated an active angiogenic remodeling in the surrounding stroma where capillary congestion and microhemorrhages occurred. Endothelial sprouting of the host capillaries was seen. Vascular co-option between the endothelial cells, newly formed endothelial cells, and the tumor cells was observed. No clear basal lamina was seen. Tumor-endothelial cell mosaics and vascular mimicry were detected only in the ES and OS. Vascular expression and neogenesis varies considerably from the fast ES to the fibrosarcoma while the OS reproduces patterns that are common in all the 3 models.
Conclusion
ATs and in vivo tumor xenograft biobanking are considered a useful pair, because they represent important resources in the common study of tumor progression, facilitating some steps in translational research. AT biobanking provides a huge amount of specimens, which represents the entire clinical heterogeneity, for molecular analyses of validation studies, and xenograft models offer an easy source of fresh material and are also a resource for in vivo experiments. The availability of these biobanks allows for the study of specific genetic alterations in the original tumor and the corresponding xenografts, additionally testing several biomarkers for future applications in translational research.
Footnotes
Acknowledgment
This study was performed with support from the EuroBoNet Network (Grant No. 018814).
Author Disclosure Statement
No competing financial interests exist.