
Abstract
An identified impediment to the advancement of science in the field of proteomics is the deterioration of proteins in tissue upon removal of the tissue from its natural state. To reduce this degradation, human tissues are frozen and stored in either liquid nitrogen or −80°C environments. It is believed that frozen tissue in ultralow temperatures preserves proteins against enzyme degradation. Various molecular, biophysical, and biochemical analytical studies require that frozen tissues be thawed before being used for analyses. Depending on downstream analyses, tissues are thawed at different temperatures (37°C, room temperature or 4°C). However, there is very little literature that describes the effects of different thaw temperatures on enzymatic inactivation in tissue lysates. We investigated the effects of preprocessing variable thaw temperature on postprocessed lysates using tyrosine phosphatase and phosphatase and tensin homolog activity assays. In our study we examined the thawing of frozen human thyroid tissues at the traditional temperatures of 4°C (on ice), 37°C (in an oven), and the novel temperature of 95°C (using Stabilizor T1™). The tissue lysates were processed without the addition of enzymatic inhibitors. Our results showed that in benign, malignant, and diseased tissues, high temperature thawing is effective in reducing enzymatic activity. In normal tissue, the reduction is dependent on individual enzymes. This suggests that if tissue lysates are to be obtained from frozen tissues without the addition of inhibitors, high temperature thawing might have marked improvement in downstream non-enzymatic analyses of diseased and neoplastic tissues.
Introduction
The use of cryopreserved tissue in research experiments entails a combination of 2 distinct processes known as the freeze–thaw process. In the first part of this process, tissues are initially frozen and then stored at ultralow temperatures to preserve protein. However, either rapid or slow cooling may cause damage to the cell. Rapid cooling causes the formation of ice crystals within the cell, damaging the cell membrane by direct mechanical action. In contrast, during a slow cooling process, ice crystals are formed outside the cell resulting in a hyperosmotic environment within the cell. Damage in this case is caused by secondary effects due to changes in composition in the liquid phase within the cell.4,5
The second component of the freeze–thaw process (ie, thawing) begins when tissues are removed from ultralow temperatures for analytical studies. Cryopreserved tissues can be thawed at different temperatures depending on various molecular, biophysical, and biochemical downstream analyses. For example, rat liver slices used for electrical impedance studies were rapidly thawed in a saline solution at 37°C, 6 whereas dynamic viscoelastic properties of cartilage were investigated by thawing at 37.5°C, 7 both suggesting that a temperature of 37°C is generally used for thawing tissue slices. However, frozen slide sections that are used for histological and immunohistological staining are generally thawed and processed at room temperature. 8 For tissues that require mincing, thawing at 4°C seems to be the temperature of choice.9,10
Any freeze–thaw cycle may result in damage to cells or their components, cell membranes, or the cytoskeleton. 11 To limit this damage, a single or combination of cryoprotecting agents (CPA) such as sucrose, dimethyl sulfoxide (DMSO), and polyethylene glycol can be used. 5 For example, DMSO has been used for several decades as the most efficient CPA for many types of cells and tissues. 12 However, the use of CPAs still does not protect the tissues from experimental manipulations that are necessary for biochemical analysis. In addition, these manipulations may introduce proteolytic artifacts that were not present in the intact tissue. 2
The freeze–thaw process can modify the activities of phosphorylated proteins 13 possibly due to reactivation of enzymatic activity during thawing (4°C–37°C). However, at predominantly high temperatures, thermodynamic effects can cause changes in protein conformation, resulting in protein denaturation and subsequent loss of protein function. 14 Devices such as the microwave and heat Stabilizor T1™ from Denator AB are capable of rapidly producing high temperatures of 80°C–90°C and 95.1°C respectively. In tissues, this high temperature leads to a permanent reduction in enzymatic activity, thereby minimizing enzyme-dependent metabolic changes. The use of the microwave for animal tissue preparation by irradiation of the animal has been in use since 1970s while Stabilizor T1 is a recent heat stabilization technology. Studies using heat Stabilizor T1 have shown that the high temperature of 95°C can inactivate enzymes.15–18 Hence, we propose that the heat Stabilizor T1 will be effective in reducing enzymatic activity during the thawing process.
Currently, there is very little literature that describes the effects of different thaw temperatures on enzymatic inactivation in tissue lysates. To understand the effects of the preprocessing variable thaw temperature on postprocessed lysates, we examined 2 phosphatase enzyme activities in human thyroid tissues thawed at different temperatures. The phosphatase enzymes that we examined were tyrosine phosphatase and phosphatase and tensin homolog (PTEN). Svensson et al. examined tyrosine phosphatase activity on heat-stabilized mouse tissues. 16 We replicated part of this study in human samples. The second enzyme PTEN was investigated due to its importance as a tumor suppressor that constitutes the main node of the PI3K-Akt pathway 19 and its differential expression in normal and neoplastic thyroid tissues. 20 Our study compares the presence of phosphatase activities in lysates of diseased, benign, malignant, and normal thyroid tissues thawed at traditional temperatures of 4°C, 37°C, to the novel temperature of 95°C.
Materials and Methods
Tissue procurement
Samples were collected, coded, and anonymized according to standard operating procedures for tissue procurement by the Cooperative Human Tissue Network (CHTN). Remnant thyroid tissues from surgical procedures were provided by the CHTN, which is funded by the National Cancer Institute. All procedures were conducted in accordance with Institutional Review Board of the University of Pennsylvania. The categories of thyroid tissue used in this study were benign, diseased, malignant, and normal (Table 1).
Thyroid was selected since it represents an organ that is affected by multiple diseases as reflected by the diagnosis within each category (n=3–4 cases/category).
Thyroid samples were dissected and placed in Mini Poly-Cons® biospecimen containers (volume capacity of 0.5 oz) obtained from Poly-Cons. The container was then closed and submerged in liquid nitrogen (LN2) for freezing. The frozen samples were then stored in a −80°C freezer until thawed. The average time from surgical resection to cryopreservation was 60±20 min. Before freezing, an aliquot was obtained from each sample, and processed to a formalin-fixed and paraffin-embedded (FFPE) block. A hemotoxylin and eosin slide was prepared from each FFPE block and reviewed by a board certified pathologist to confirm the diagnosis.
Tissue thaw methods
Frozen thyroid tissues of comparable weights and sizes were thawed 1 of 3 ways: 4°C (on ice), 37°C (in the oven), or at 95°C (using Stabilizor T1). Tissues were thawed at 4°C by placing the sample container on wet ice for 4–6 min, whereas tissues thawed at 37°C were placed in the oven for 10 min in open sample containers. Stabilizor T1 instrumentation (Denator AB) was used to thaw samples at 95°C as described in Svensson et al. Briefly, the samples were placed in inert Teflon-fluorinated ethylene propylene Maintainor Cards (Denator AB), and the air was removed by automated vacuum to minimize potential protein oxidation and maximize efficient heat transfer. Specimens were then subjected to a combination of pressure and heat, with the LASER determining the amount of time required for each tissue to be heated 16 (Table 2). Frozen unthawed tissues processed immediately upon removal from the −80°C freezer were used as controls.
Tissues were taken from −80°C storage and thawed 4°C (on ice), 37°C (in the oven), or at 95°C (using Stabilizor T1).
Tissue lysate preparation
After thawing, the tissues were homogenized using TissueLyser LT® (Qiagen, Inc.). The homogenization was performed in lysis buffer from the SensoLyte pNPP Protein Phosphatase Assay Kit® (AnaSpec) without using protease or phosphatase inhibitors.
Protein isolation and quantitation
The homogenate was centrifuged at 16,000 g for 20 min at 4°C to pellet cell debris. The supernatant was collected and stored at −80°C until protein concentration was measured by BCA assay (Pierce BCA Assay™; Thermo Fisher Scientific, Inc.).
Phosphatase activities determination
Tyrosine phosphatase activity was calorimetrically assayed by measuring absorbance at 405 nm using the SensoLyte pNPP Protein Phosphatase assay kit (AnaSpec). The phosphatase assay buffer in the phosphatase assay kit has been formulated to measure protein tyrosine phosphatase 1b (PTP-1B).
PTEN activity was calorimetrically assayed by measuring absorbance at 450 nm using the PathScan® Total PTEN Sandwich ELISA Kit (Cell Signalling Technology).
Data analysis
Statistical analyses were performed using Students 2-tailed t-tests.
Results
The effects of thaw temperatures were investigated by following phosphatase activities in different categories of thyroid tissues: diseased, benign, malignant, and normal. The novel temperature of 95°C was compared with 2 traditional thaw temperatures (4°C and 37°C) as well as unthawed samples. The results obtained for tyrosine phosphatase were plotted as graphs across various protein concentrations (1, 0.5, and 0.1 μg/μL) as a measure of absorbance observed at 405 nm (Fig. 1). The trend in phosphatase activity levels at the highest concentration of 1 μg/μL was observed to be similar to phosphatase activity levels at lower 0.5 μg/μL and 0.1 μg/μL concentrations. Therefore, for the second enzyme, PTEN, the results were plotted at the highest protein concentration of 1 μg/μL across various categories of thyroid as a measure of absorbance observed at 450 nm (Fig. 2).
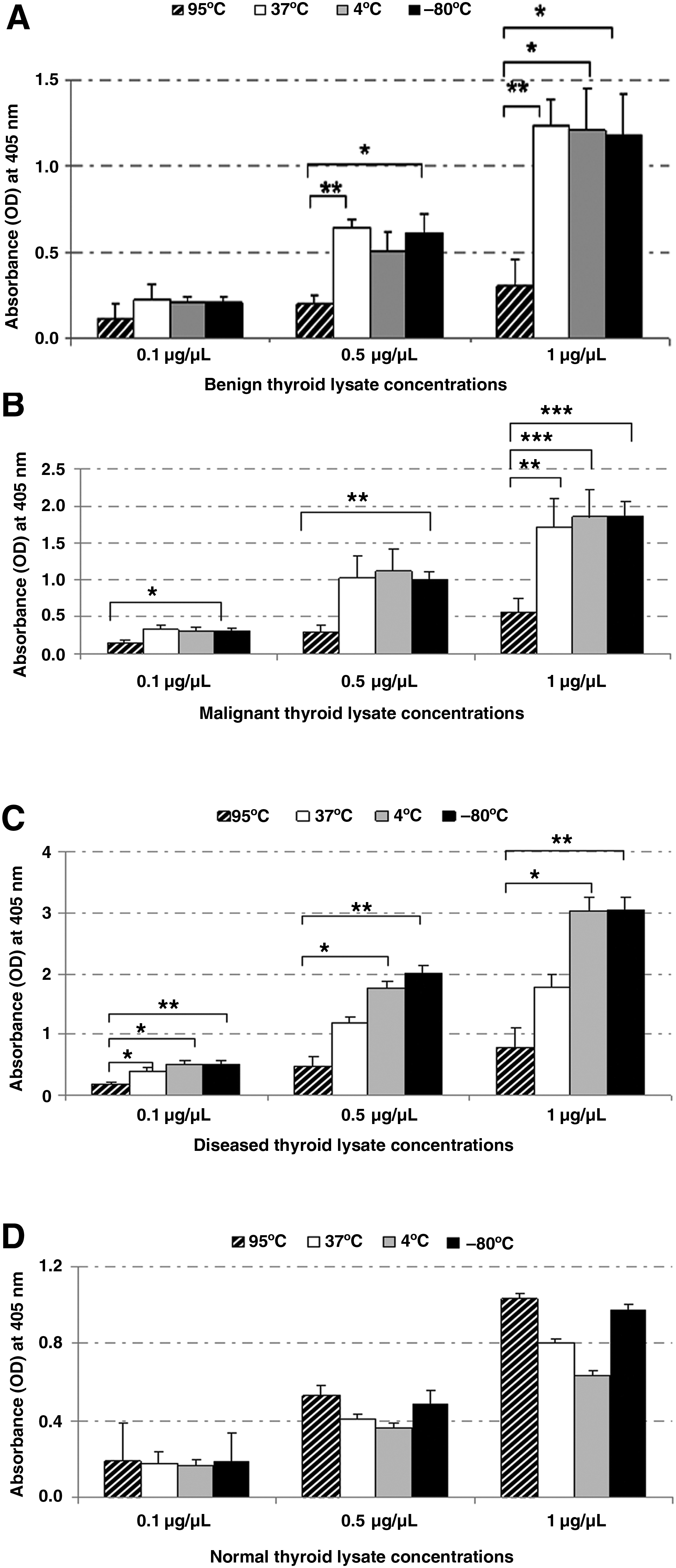
Tyrosine phosphatase activity in thyroid tissue lysates. The effects of different thawing temperatures were studied by following tyrosine phosphatase activity in different categories of thyroid tissues:

PTEN activity in thyroid tissue lysates. The effects of thaw temperatures were investigated using another phosphatase enzyme, PTEN, in different categories of thyroid tissue lysates. As earlier, the novel temperature of 95°C was compared with 2 traditional thaw temperatures (4°C and 37°C) as well as unthawed samples. The results obtained for each category of tissue lysates were plotted as graphs across 1 μg/μL protein concentrations as a measure of absorbance observed at 450 nm. Data represented as mean±SEM, *P<0.05, **P<0.01, ***P<0.001, Student's t-test, n=6–8/thaw temperature/category. PTEN, phosphatase and tensin homolog.
Enzyme activity in benign thyroid tissues
We observed consistent trends in the reduction of both tyrosine phosphatase and PTEN enzyme activities in 95°C thawed benign tissue lysates. A significant reduction of tyrosine phosphatase activity was observed in benign heat-stabilized lysates (95°C) at the highest concentration of 1 μg/μL, in comparison to lysates thawed at 37°C (P<0.01, n=9–12/thaw group), 4°C (P<0.05, n=9–12/thaw group), and −80°C (P<0.05, n=9–12/thaw group). A similar trend was observed at the lower concentration of 0.5 μg/μL, viz., a significant reduction in activity was seen in the 95°C thawed tissue lysates in comparison to the 37°C (P<0.01, n=9–12/thaw group) and −80°C (P<0.05, n=9–12/thaw group) tissue lysates (Fig. 1A).
The PTEN enzyme activities followed a trend with lowest enzyme activity observed in heat-stabilized tissues (95°C). Similar to tyrosine phosphatase activity, the activities observed for unthawed lysates and lysates thawed at 37°C and 4°C seemed to be of comparable to each other without significant differences (Fig. 2).
Enzyme activity in malignant thyroid tissues
In malignant heat-stabilized lysates, similar to benign tissues, at the highest concentration of 1 μg/μL, a significant reduction of tyrosine phosphatase activity was observed in heat-stabilized tissues (95°C) in comparison to lysates thawed at 37°C (P<0.01, n=9–12/thaw group), 4°C (P<0.001, n=9–12/thaw group), and −80°C (P<0.001, n=9–12/thaw group). Also, at lower concentrations of 0.5 μg/μL and 0.1 μg/μL tissue lysates, a significant reduction (P<0.01 and P<0.05, respectively, n=9–12/thaw group) in activity was seen in the 95°C thawed tissue in comparison to the −80°C lysates (Fig. 1B).
As noted with the benign tissues, PTEN activities in malignant lysates followed the trend with lowest enzyme activity observed in heat-stabilized tissues (95°C). Again, similar to tyrosine phosphatase activity, the PTEN activities observed for unthawed lysates and lysates thawed at 37°C and 4°C seemed to be comparable to each other (Fig. 2).
Enzyme activity in diseased thyroid tissues
In diseased tissue lysates, a significant reduction in tyrosine phosphatase activity was also observed in tissues thawed at 95°C in comparison to unthawed tissues (P<0.01, n=9–12/thaw group), and tissues thawed at 4°C (P<0.05, n=9–12/thaw group). The enzyme activity was the lowest in heat-stabilized lysates (95°C) and the highest in frozen, unthawed samples (Fig. 1C). Similar to tyrosine phosphatase activity, PTEN enzyme activity was also significantly reduced in heat-stabilized lysates in comparison to 37°C (P<0.001, n=6–8/thaw group), 4°C (P<0.05, n=6–8/thaw group), and −80°C (P<0.01, n=6–8/thaw group) tissue lysates (Fig. 2).
Enzyme activity in normal thyroid tissues
The results of the normal tissue lysates differed between the 2 phosphatase assays. The PTEN assay detected the same trend as the other categories of tissues (benign, malignant, and diseased).
PTEN enzyme was significantly reduced in heat-stabilized lysates (95°C) in comparison to 37°C (P<0.01, n=6–8/thaw group), 4°C (P<0.05, n=6–8/thaw group), and −80°C (P<0.05, n=6–8/thaw group) tissue lysates (Fig. 2). However, the tyrosine phosphatase assay observed a reversal of trend in normal tissues. In contrast to both diseased and neoplastic tissue lysates, among unthawed controls and lysates thawed at 4°C, 37°C, and 95°C, the heat-stabilized tissue lysates (95°C) had the highest activity (Fig. 1D).
Discussion
In our current study, we investigated the effects of different thawing temperatures using thyroid tissues. We selected thyroid since it represents an organ that is affected by multiple diseases as evidenced by the categories in non-neoplastic and neoplastic (benign and malignant) diseases (Table 1). 21 The effects of thawing temperatures were investigated by following the activity level of 2 phosphatase enzymes. We focused on a phosphatase enzymes because phosphorylation can vary greatly based on the freeze/thawing of samples and therefore not accurately reflect biological alterations in vivo. During the freeze–thaw process, enzyme distributions within a cell may vary. For example, Scharf et al. showed that the freeze–thaw process of stored mouse tissue samples resulted in a dramatic increase of alpha-AMP-activated protein kinase (AMPK) phosphorylation in comparison to fresh samples. 13 Also, Bjerknes and Cheng noted that the distribution of alkaline phosphatase activity was different from the fresh tissue in duodenal mucosa preparations that were subjected to a freeze–thaw cycle before incubation. 22
Traditionally, exogenous inhibitors are used to reduce the effect of proteome degradation in tissue lysates. However, the introduction of these inhibitors can have a negative impact on the downstream analysis. 2 In a study focused on the temperature-dependent postmortem degradation of protein phosphatase 2A (PP2A) in mice, Poloz and O'Day observed that protease inhibitors and calmidazolium led to increased levels of PP2A, thus not accurately reflecting the level of the phosphatase in live tissue. 23
One way of inactivating enzymes without inhibitors is through the use of high temperatures obtained by microwave focused beam irradiation. Microwave irradiation is generally applied during the postmortem preparation of tissues from small animals. 24 However, the physical characteristics of microwave frequency and oven cavities can cause uneven heating throughout the tissue, resulting in unequal deactivation of enzymatic activity. 25 For this reason, we chose Stabilizor T1 to investigate thawing at 95°C.
The effects of different thawing temperatures were investigated by following the activity reduction of tyrosine phosphatase and PTEN. Earlier work done on heat-stabilized mouse tissues by Svensson et al., using tyrosine phosphatase had shown reduced activation of the enzyme in heat-stabilized tissues in comparison to rapidly frozen tissues. 16 Hence, we used a commercially available tyrosine phosphatase kit, and examined the activity reduction in human tissues. The phosphatase assay buffer in the phosphatase assay kit is formulated to measure PTP-1B, an enzyme that belongs to the Class I PTPases. The structural motifs within the intracellular PTPases serve to localize these enzymes to a variety of sites in the cells (membranes, nucleus, and cytoskeletal elements). Thus, the PTPases are involved in diverse signaling pathways, 26 suggesting an active role in dephosphorylation of several cellular substrates involved in cell signaling. Thus, during the thaw process, a reduction in tyrosine phosphatase activity might decrease any ex vivo post-translational modifications to cell signaling proteins and hence increase the probability of discovering the substrates in their native state.
The second phosphatase enzyme we examined was PTEN, a tumor suppressor that is found to be downregulated in cancer, even in the absence of genetic loss or mutation. PTEN modulates 2 seemingly distinct cellular functions of cell growth/survival and cell migration/adhesion through its lipid and protein phosphatase activities, respectively.19,27–29 Thus, similar to tyrosine phosphatase enzyme, a reduction in PTEN enzyme activity might decrease any ex vivo post-translational modifications that PTEN can cause to its substrates during thaw process.
Our results obtained without the addition of inhibitors in non-normal (benign, malignant, and diseased) thyroid lysates showed that the 95°C heat-stabilized tissue had the least tyrosine phosphatase activity across all concentrations compared with traditional thaw methods. For neoplastic tissues, the observed activity range (high to low) was as follows: unthawed lysates, lysates thawed at 4°C, lysates thawed at 37°C, and finally lysates thawed at 95°C. The observed trend in activity in diseased tissues was unthawed lysates ≥ lysates thawed at 4°C > lysates thawed at 37°C > lysates thawed at 95°C (Fig. 1A–C). The observed results are consistent with the mouse brain study that showed greater phosphatase inactivation in the heat-stabilized tissues when compared with the rapidly frozen samples even when phosphatase and protease inhibitors were added to the extraction buffer. 16 Similar results were obtained with randomized human diseased and neoplastic cases as in controlled mouse tissue.
In comparison to neoplastic and diseased tissues, our results with normal tissue lysates indicated that the phosphatase assays detected lower enzymatic activity levels for both tyrosine phosphatase and PTEN. Despite this overall reduced level of activity, our PTEN assay continued to show the trend of reduced enzyme activity in the heat-stabilized lysates (95°C) compared with unthawed controls and lysates thawed at 4°C and 37°C (Fig. 2). However higher tyrosine phosphatase activity levels were observed in both the heat-stabilized (95°C) and control samples in comparison to 4°C and 37°C thawed samples. The trend can be represented as follows: lysates thawed at 95°C ≥ unthawed lysates > lysates thawed at 37°C > lysates thawed at 4°C (Fig. 1D). The difference in the trends observed for tyrosine phosphatase and PTEN heat-stabilized lysates (95°C) in comparison to thawed (4°C, 37°C) and unthawed samples lead us to infer that enzymatic reduction by thawing at 95°C heat stabilization is dependent on the enzyme and tissue type.
Thawing at 95°C denatures protein and thus presumably stops the activity by the loss of native conformation. Since enzymes are proteins, loss of their conformation results in loss or reduced enzyme function implying the substrates that would have been modified to be present in their native state. Thus proteins obtained from tissues thawed at 95°C can be used in proteomic analyses both at the protein and peptide levels as shown in studies using 2 dimensional electrophoresis (protein level) and mass spectrometry (peptide level).17,30,31
Due to the decrease in enzyme activity in heat-stabilized (95°C) lysates, presumably because of loss of conformation at high temperature, 95°C thawing of tissue cannot be used for any activity assay that requires native conformation, including phosphatase activity assays. In addition, as seen in the normal thyroid tissue, the loss of activity is dependent on individual enzyme. However, translational modification analyses such as acetylation or glycosylation may not be affected by 95°C thawing of tissues as shown by Rountree et al., 32 suggesting that the tissues thawed at 95°C can be utilized for acetylation or glycosylation assays.
This initial study has shown the effectiveness of the 95°C thaw temperature using Stabilizor T1 system in reducing the enzyme activity in human thyroid tissue. Our investigation suggests that if tissue lysates are to be obtained from frozen tissues without the addition of inhibitors, diseased and neoplastic tissues might have marked improvement in downstream non-enzymatic analyses from thawing at high temperatures. The applicability of this temperature to effectively thaw other tissue types are yet unknown. We are currently investigating the effects of this novel temperature to evaluate different enzymes, varying time preservation intervals, and additional human tissue types.
Footnotes
Acknowledgments
We would like to thank Ms. Indira Prabakar, Ms. Christina Arana, and Ms. Elicia Yoffee for helping with experiments, and Ms. Munia Islam, and Mr. Andrew Tieniber for helping edit the article. This research was supported by the National Cancer Institute.
Author Disclosure Statement
No potential conflicts of interest for any of the authors.