
Abstract
Biochemical analyses of many brain diseases have highlighted that oxidative damage of proteins and astrogliosis are important events associated with pathology. However, human studies on the status of protein oxidation/nitration and astrogliosis [indicated by expression of glial fibrillary acidic protein (GFAP)] heavily depend on postmortem tissues that might be altered by pre and postmortem factors. To evaluate the effect of these variables, we tested whether the status of GFAP expression, oxidized proteins, and nitrated proteins (by protein 3-nitrotyrosine or 3-NT) were affected in postmortem human brains (n=48) by increased storage time (11.8–104.1 months), postmortem interval (PMI) (2.5–26 h), gender difference, and agonal state (based on Glasgow coma scale: range: 3–15) in different anatomical regions—frontal cortex (FC), cerebellum (CB) and medulla oblongata (MD). We observed that increasing storage time significantly decreased the stability of all 3 markers in MD (oxyblot: P=0.003; 3-NT: P=0.01; GFAP: P=0.03) and that of oxidized proteins in CB (P=0.04), whereas the status of all markers was not significantly altered in FC. On the other hand, PMI and agonal state did not influence the status of all the markers tested in any of the regions. Similarly, except for the decreased protein 3-NT among women in CB compared with men (P=0.04), there was no effect due to gender differences in other brain regions for other markers. These data highlight the influence of storage time on preservation of markers of protein damage and astrogliosis and the inherent differences in brain regions, with implications for studies on brain pathology employing stored human samples.
Introduction
However, the data regarding these processes obtained from experimental studies are based on animal models and require validation in human tissue. For this, researchers rely on the postmortem samples for biochemical studies due to nonavailability of fresh brain tissue. However, the data from postmortem samples are dependent on tissue quality, which, in turn, is influenced by age, gender, medication, and agonal state (termed as premortem factors), and postmortem interval (PMI), tissue preservation, and duration and the conditions of storage (termed as postmortem parameters). Although the role of pre and postmortem factors in RNA stability and gene expression has been investigated,10,11 studies on brain proteins are limited. 12 There are some previous reports including ours 13 and others14,15 that have demonstrated the effect of agonal state and PMI on specific proteins. This has serious implications for studies that are aimed at correlating the presence of different bio-markers with disease processes. If the status of GFAP and oxidatively damaged proteins in the brain tissue is altered independent of disease-related changes, then this will significantly affect the interpretation of the experimental results. Hence, it is pertinent to investigate the effect of pre and postmortem factors on oxidative protein damage and GFAP expression in human brains.
In the current study, we have analyzed the postmortem brains (n=48) stored in the Human Brain Tissue Repository (HBTR) to determine whether increasing PMI (2.5–26 h), agonal state [based on Glasgow coma scale (GCS): range: 3–15], prolonged storage time at −80°C (11.8–104.1 months), and gender of the subjects have any effect on the status of GFAP expression and oxidized and nitrated (protein 3-nitrotyrosine or 3-NT) proteins in postmortem tissues.
Materials and Methods
All the chemicals used were of analytical grade. Bulk chemicals were obtained from Merck and Sisco Research Laboratories Pvt. Ltd. Fine chemicals, anti-3-NT and anti-dinitrophenyl polyclonal antibodies, and protease inhibitor cocktail were from Sigma. Anti-β-actin polyclonal antibody from Imgenex Corp. and anti-GFAP monoclonal antibody from Dako were procured. Nitrocellulose membrane was obtained from Millipore. Horseradish peroxidase-conjugated secondary antibodies were obtained from Bangalore Genei.
Human tissue samples
Human brain samples were sourced from the HBTR, Department of Neuropathology, National Institute of Mental Health and Neurosciences (NIMHANS), Bangalore, India. The brain tissues were collected after approval from the institutional ethics committee and written informed consent from close relatives of the subjects. The brains were from normal subjects who had succumbed to road traffic accidents (nonalcoholics, nondiabetics, not on any medication, and with no known neurological or psychiatric disorders). Demographic and clinical details of all the subjects and the agonal state scored by the GCS were collected from medical records. The GCS score was based on (1) best eye response (E; range: 1–4), (2) best verbal response (V; range: 1–5), and (3) best motor response (M; range: 1–6) with a total range of 3 to 15 (noted at the time of hospital admission) with score=3 corresponding to the most severe agonal state. The hospital stay after admission varied among the patients. During the hospital stay, assisted ventilation and parentral administration of mannitol, hydrocortisone, and ionotropic drugs were administered to maintain the vital parameters whenever necessary.
After the death of the patients, the PMI (the time interval between the time of death and placing the dissected brain slices into the freezer maintained at −80°C) was recorded. Within 1 h of death, routinely, the body was transferred to a refrigerator maintained at 2°C–4°C with a recorder and uninterrupted power supply. After autopsy, the brains were recovered, coronally sliced, kept flat on a salt-ice mixture (−15°C to −18°C) during dissection, and then transferred in plastic zip lock bags into a box to be stored at −80°C in the HBTR. The procedure of dissection took ∼30–45 min, and the brain slices that had been transported on the salt-ice mixture were immediately transferred into the deep freezer. The brain areas chosen for the study were anatomically farthest from the site of injury and without distinct edema or grossly apparent pathology. Although the major portion of the tissue was frozen for biochemical studies, a portion corresponding to the mirror image of the stored tissue bits was fixed in buffered formalin. These tissues were subjected to histopathological assessment, and the samples that maintained tissue integrity were utilized for the study (data not shown). The protocol of autopsy, tissue handling, and other procedures were uniform for all the samples. Such tissues have been earlier utilized as control samples in PD research and other studies.16,17
In the current study, 3 anatomical areas, frontal cortex (FC), cerebellum (CB), and medulla oblongata (MD), were analyzed from 48 postmortem brains (Table 1). The gender distribution (men: n=31; women: n=17), age (range: 2 days to 80 years), PMI (range: 2.5–26 h), storage time (11–104.1 months), and agonal state (GCS score: 3 to 15) were noted in all the cases.
M, male; F, female; PMI, postmortem interval; GCS, Glasgow coma scale; NA, not available.
Histology
The formalin-fixed tissues were embedded in paraffin, sectioned, and stained with hematoxylin-eosin, Nissl, and Luxol fast blue for myelin. 13 The sections were examined (blinded to storage time, PMI, agonal state, and gender) for myelin pallor, neuronal staining character, and anoxic changes by 2 neuropathologists (A.M. and S.K.S.).
Preparation of protein extracts
Brain tissue (∼100 mg) was manually homogenized in cold 1× phosphate buffered saline containing protease inhibitor cocktail on ice and sonicated in a Sonics-vibra cell sonicator (Sonics and Materials Inc.). 13 The extract was centrifuged (15,000 g, 10 min) to separate the insoluble debris. The supernatant corresponding to the soluble fraction was subjected to protein estimation followed by biochemical assays.
Estimation of pH
pH of the frozen brain tissue was determined immediately after thawing, based on the method previously described. 18 The tissue samples (∼150 mg) were manually homogenized in 10 volumes of distilled water adjusted to pH 7.0 (1:10 w/v). pH was recorded in the homogenates at room temperature by using a pH meter (Control Dynamics) previously calibrated with 2 standards (pH 4.0 and pH 7.0).
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis and western blot
Brain total protein extracts (∼50 μg/lane) were run on 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS PAGE) followed by western blot, with antibodies against GFAP and β-actin as previously described. 13
Estimation of protein carbonyls, protein nitration, and GFAP
Oxyblots were carried out as described earlier.13,19 Briefly, brain protein extract (4 mg/mL) was derivatized by dinitrophenyl hydrazine (DNPH) in the presence of 12% SDS for 20 min at room temperature. The reaction was stopped with 2 M Tris in 30% glycerol, and 5 μL of the sample was spotted in triplicate onto a nitrocellulose membrane under vacuum pressure by using a vacuum manifold (Scie-Plas Ltd.). The membrane was subjected to western analysis with anti-DNP antibody with appropriate controls. For quantitation of protein 3-NT, GFAP, and β-actin, soluble protein (10 μg/sample) from different samples was spotted in triplicate and subjected to slot blot as previously described. 13
All the blots were dried and scanned in a gel documentation system (BioRad Laboratories), densitometrically quantified, and normalized against the respective anti-β-actin signal by using dedicated software (Quantity One; BioRad laboratories).
Statistical analysis
Quantitative data were accumulated from at least 3 independent experiments and expressed as mean±standard deviation followed by analysis of variance and Pearson's correlation of linear regression (r value). In all the experiments, data with P<0.05 were considered statistically significant.
Results
We first assessed the quality of the brain samples by measuring the tissue pH in the 3 anatomical areas FC, CB, and MD in all the samples and compared them with regard to storage time, PMI, gender, and agonal state (GCS score). Although the pH value varied in FC from 5.45 to 7.44, there was no significant difference with increasing storage time (r=0.102, P=0.249) (Fig. 1A). Similarly, CB (pH range=5.56–6.92) and MD (range=5.64–7.18) also did not vary in pH with increasing storage time (CB: r=−0.125, P=0.208; MD: r=0.023, P=0.44). Tissue pH did not vary significantly with increasing PMI (FC: r=0.183, P=0.115; CB: r=−0.184, P=0.115; MD: r=0.105, P=0.248) (Fig. 1B) and GCS score in all the 3 regions tested (FC: r=−0.05, P=0.38; CB: r=0.17, P=0.13; MD: r=0.01, P=0.46) (Fig. 1C). Gender-based comparison of pH values showed that although the pH varied in men and women in FC (men: mean=6.23±0.37; women=6.14±0.36), CB (men: mean=6.2±0.31; women: mean=6.08±0.23), and MD (men: mean=6.12±0.33; women: 6.14±0.19), there was no statistically significant difference in the anatomical areas (FC: P=0.445; CB: P=0.117; MD: P=0.872) (Fig. 1D).
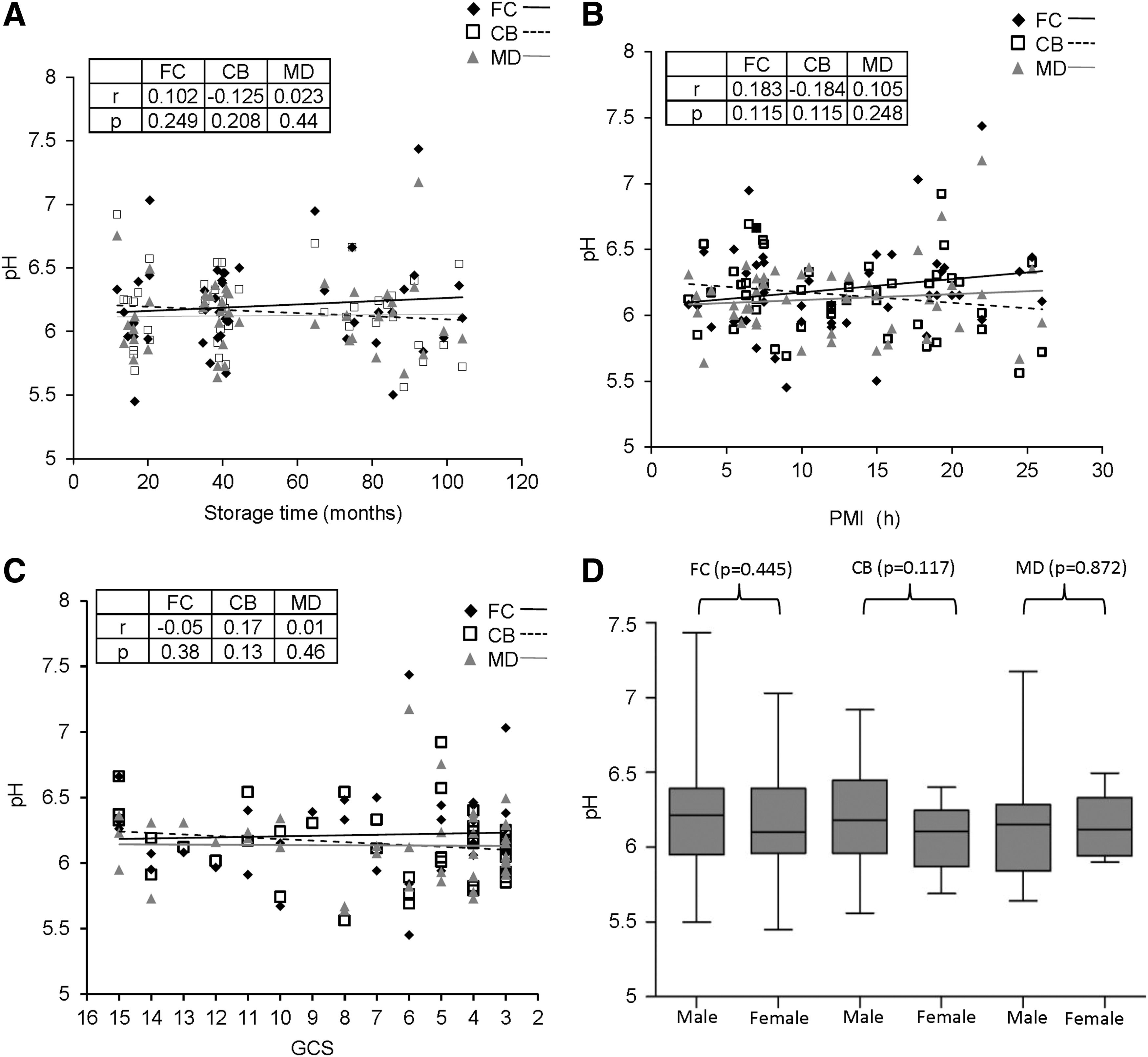
Analysis of the effects of pre and postmortem factors on tissue pH in postmortem brains. Tissue pH in frontal cortex (FC), cerebellum (CB) and medulla oblongata (MD) of post mortem human brain samples (n=48) was estimated in duplicate, and the average for each point was plotted as mean±standard deviation (SD) followed by regression analysis as denoted by r and P values. Scatter plot graph with linear regression line for distribution of tissue pH in the 3 regions, with increasing storage time
Next, we tested whether the status of oxidatively damaged brain proteins were altered by pre and postmortem factors. This was carried out by western blot based quantitation of protein carbonylation (and corresponding β-actin signal) in different samples by using the slot-blot method. The protein carbonyls/β-actin ratio with increasing storage time showed a significant decrease in CB and MD, whereas it was relatively unaltered in FC (FC: r=−0.14, P=0.19; CB: r=−0.27, P=0.04; MD: r=−0.42, P=0.003) (Fig. 2B). However, the protein oxidation status was unchanged in any of the regions with increasing PMI (FC: r=−0.06, P=0.35; CB: r=−0.01, P=0.47; MD: r=0.02, P=0.45) (Fig. 2C), agonal state (FC: r=0.16, P=0.15; CB: r=0.20, P=0.10; MD: r=0.07, P=0.32) (Fig. 2D), and gender difference (FC: P=0.134; CB: P=0.28; MD: P=0.054) (Fig. 2E).
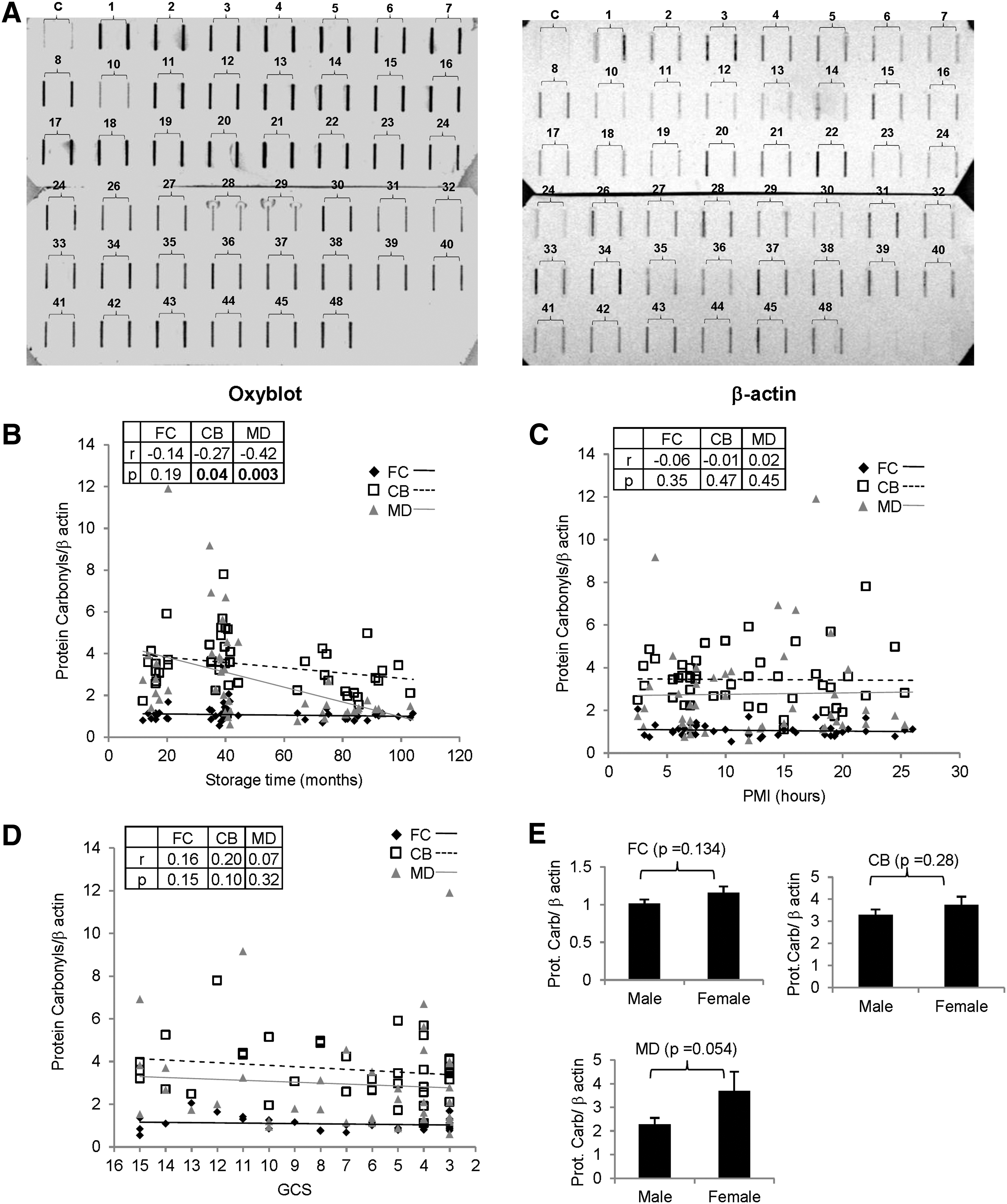
Effect of pre and postmortem factors on protein oxidation/carbonyls in postmortem brains. Oxyblot-based assessment of protein oxidation status in postmortem human brain samples (n=48) was assayed.
We then quantitated protein 3-NT (and corresponding β-actin signal) in different samples by using the slot-blot approach. The protein 3-NT/β-actin ratio with increasing storage time showed a significant decrease in MD, whereas it was unchanged in FC and CB (FC: r=0.20, P=0.10; CB: r=0.13, P=0.20; MD: r=−0.36, P=0.01) (Fig. 3B). However, the protein nitration status was unchanged with increasing PMI (FC: r=−0.02, P=0.44; CB: r=0.15, P=0.17; MD: r=−0.03, P=0.44) (Fig. 3C) and agonal state (FC: r=0.01, P=0.47; CB: r=−0.11, P=0.25; MD: r=0.15, P=0.16) (Fig. 3D) in any of the regions. Although gender-based differences were insignificant in FC and MD, protein nitration was lower in women compared with men in CB (FC: P=0.35; CB: P=0.043; MD: P=0.182) (Fig. 3E).
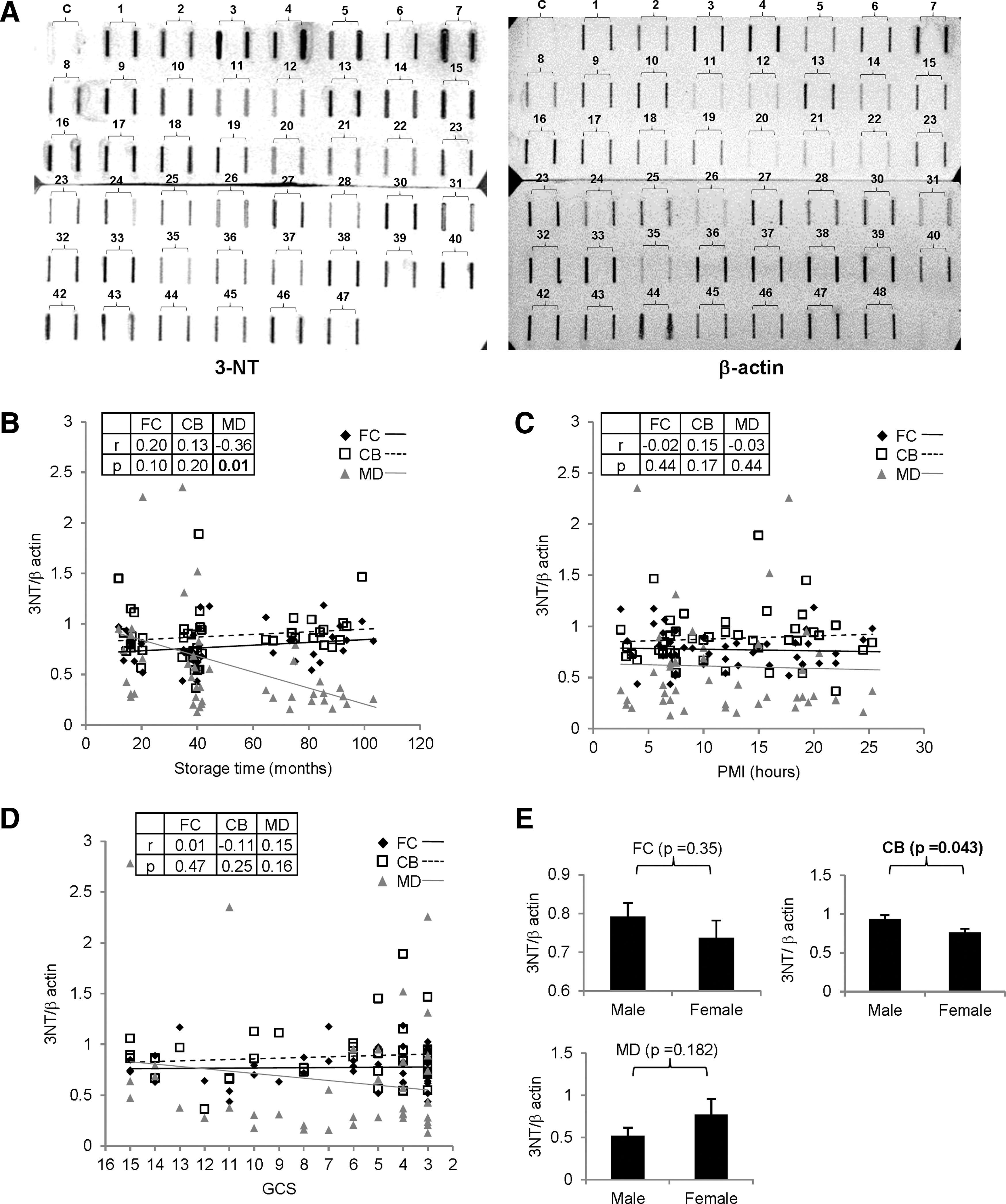
Effect of pre and postmortem factors on protein nitration (3-NT) levels in different anatomical areas of postmortem brains. Slot-blot based assessment of protein nitration status in postmortem human brain samples (n=48) was assayed.
Next, we quantitated the GFAP expression (marker of astrogliosis) and the corresponding β-actin levels by using the slot blot in different samples. The GFAP/β-actin ratio with increasing storage time showed a significant decrease in MD, whereas it was unchanged in FC and CB (FC: r=−0.08, P=0.31; CB: r=−0.05, P=0.39; MD: r=−0.29, P=0.03) (Fig. 4B). However, the GFAP status was unchanged in any of the regions with increasing PMI (FC: r=0.20, P=0.10; CB: r=0.06, P=0.34; MD: r=−0.14, P=0.18) (Fig. 4C), agonal state (FC: r=−0.04, P=0.40; CB: r=0.12, P=0.23; MD: r=0.10, P=0.26) (Fig. 4D), and gender difference (FC: P=0.67; CB: P=0.44; MD: P=0.217) (Fig. 4E).

Effect of pre and postmortem factors on levels of glial fibrillary acidic protein (GFAP) in postmortem brains. Slot-blot based quantitaion of GFAP in postmortem human brain samples (n=48) was carried out.
Discussion
Since postmortem human tissue is extremely important for neuroscience research, several approaches are adopted to evaluate the markers of tissue quality. Conventionally, a low PMI13,20 is a marker of high tissue quality. Brain tissue pH also reflects the integrity of postmortem tissue, as it indicates the divergent physiology and resultant metabolic changes ante-mortem and mode of death. 18 Since different anatomical areas of the brain have distinct physiology, we measured pH in FC (cholinergic), CB (GABAergic), and MD (serotonergic) and found that the pH values did not show significant variance with storage time, GCS rating, PMI, and between men and women (Fig. 1), which is consistent with recent reports. 18 Interestingly, we found 12 samples with tissue pH <6.0 that significantly represents acidic pH. However, further analysis revealed that the markers tested in samples with pH 5.45 to 6.0 were not significantly affected by acidic pH (data not shown).
Oxidative stress has been implicated in neuropathology, but the effects of disease-unrelated factors on cellular oxidative/antioxidant markers have not been investigated. Our recent study addressing this issue demonstrated that antioxidant function in postmortem brains, especially the glutathione dynamics, was significantly modulated by pre and postmortem factors mainly in MD (Harish et al., manuscript submitted). Interestingly, MD was significantly affected with regard to all parameters in the current study, thus indicating the importance of this brain region.
During brain pathology, oxidative stress might also affect protein structure function by oxidative modifications. 5 Selective oxidative/nitrative modifications of specific brain proteins might contribute to neurodegenerative diseases. For example., oxidative damage of mitochondrial complex I and DJ-1 has been detected in postmortem samples of patients with PD and AD. 21 However, the effect of pre and postmortem factors on the status of oxidized proteins needs to be delineated for proper interpretation of the results in such studies. To address this, analysis of protein oxidation in human samples in the current study did not indicate any significant effect of PMI and agonal state in all the 3 regions tested consistent with our previous study 13 (Fig. 2). However, with increasing storage time, we observed a significant loss of protein oxidation signal in CB and MD (Fig. 2) that could be due to the masking of epitopes or degradation/aggregation of oxidized proteins or other unknown mechanisms. We also observed that protein oxidation was significantly lower in female brains in CB compared with male brains, thus indicating a gender-specific alteration in protein stability and oxidation (Fig. 2). Interestingly, the importance of the role of gender in oxidative processes linked to brain aging and pathology is not well characterized. 22 Consistent with our data from human brains, other studies using animal brains have recorded significant antioxidant load and lower oxidative damage in female brains 23 with implications for neurobehavioral changes and selective susceptibility to neurodegenerative disorders.24,25
Similar to protein oxidation, most of the studies on protein nitration have concentrated on its role in cellular damage during aging, 26 neurodegeneration,7,27 and mitochondrial damage. 28 For example, α-synuclein and Parkin, 2 important proteins linked to PD, are modified by nitrosative stress, as demonstrated in the postmortem brain samples and experimental models.29,30 However, based on our study, it needs to be further validated whether the protein nitration noted in human postmortem samples is indeed representative of the pathology or the extraneous pre and postmortem factors. Analysis of postmortem brains in the current study showed that although protein-3NT signal was unaltered with increasing PMI and agonal state, MD revealed loss of protein nitration signal with increasing storage time (Fig. 3). Similar to the altered protein oxidation in MD, nitrated proteins could have aggregated into insoluble complexes masking the reactive epitopes, thereby reducing the protein nitration signals (Fig. 3).
In our earlier study, 13 we observed a marginal but statistically insignificant increase in GFAP expression in SN with increasing PMI in contrast to other anatomical areas reflecting probable regional differences in astrogliosis. In the current study, we did not record any change in the signal intensity of GFAP with prolongation of terminal agonal state or increase in PMI. However, with an increase in storage time of the human brain, the GFAP signals were similarly reduced to protein oxidation and nitration in MD but not in FC and CB (Fig. 4). We surmise that with storage, the GFAP, a fibrillar protein, aggregates, thus altering the immunological signals, but this requires further validation.
In conclusion, the current study was aimed at understanding whether pre and postmortem factors had a bearing on the postmortem preservation of oxidized and nitrated brain proteins and GFAP in 3 anatomical areas of postmortem human brain tissues stored for long periods. Tissue pH was not affected by the pre and postmortem factors, thus indicating fairly well-preserved tissue quality for the study. Interestingly, storage time had a significant bearing on the stability of oxidized and nitrated proteins and GFAP, thus reflecting alteration in the tissue quality and preservation-dependent protein structure function, which needs to be kept in mind in biochemical studies using brain tissues. However, other factors such as PMI, agonal state, and gender difference did not significantly influence the markers tested in our samples. Further, MD showed relatively higher vulnerability to postmortem factors. Such studies aimed at analyzing biomarkers are required to substantiate the reliability of biochemical studies using postmortem samples and to understand the physiological and pathological events in the brain.
Footnotes
Acknowledgments
This study was financially supported by the HBTR (S.K.S.), Department of Neuropathology, NIMHANS, and Indian Council of Medical Research (ICMR IRIS ID No. 2009-07710) (M.M.S.B.). G.H. is supported by a senior research fellowship from ICMR, India. The authors would like to thank the donors and their relatives for the kind gift of human brains for neurobiological studies.
Author Disclosure Statement
The authors declare that there are no conflicts of interest.