
Abstract
The DNA barcoding technique is often used as a tool for validating species identity in biobanks. In the case of amphibians, the mitochondrial DNA (mtDNA) 16S ribosomal RNA (rRNA) gene is reported to fulfill the requirements of a universal DNA barcoding marker. The 16S primers are designed to specifically bind to the 16S rRNA gene, which is a very well-conserved mtDNA gene sequence in amphibians. DNA was extracted from thirteen known but different species of amphibians within the Zoological Society of London/Amphibian Ark's cryobank. After this, the DNA was amplified and analyzed by 1 the traditional DNA barcoding procedure that involves conventional polymerase chain reaction (PCR) and DNA sequencing and 2 a novel procedure, involving real-time PCR and melting temperatures. Both procedures used the same 16S primers. Successful DNA amplification and validation to the species or genus level was achieved in 10 out the 13 cases using the traditional approach. Nevertheless, after real-time PCR and melting temperature analysis, some variability was found between Common Frog samples but more concerning, the same melting temperature was recorded in unrelated species (Common Toad, Common Frog and Amazon Milk Frog), despite their 16S sequences exhibiting a high degree of variability. We conclude that traditional DNA barcoding using 16S rRNA sequences is suitable for validating the specific identity of amphibian samples within biobanks and that modification of the current 16S real-time PCR and melting temperature analysis is required before it can be employed as a cheaper and faster alternative.
Introduction
DNA barcoding
The main goals of DNA barcoding are to (i) identify all eukaryotic species by rapid acquisition of molecular data,9,10,12 also in microscopic organisms with complex or inaccessible morphology, 12 in damaged organisms or fragments,9,16 (ii) discover potentially new species,4,17–19 (iii) match adults with immature specimens especially when species have polymorphic life cycles and/or exhibit pronounced phenotypic plasticity9,17,20–22 and (iv) reveal cryptic species.17,23,24 However, the main difficulty of DNA barcoding is finding a suitable gene sequence that (i) allows robust and repeatable amplification and sequencing, (ii) has sufficient variability to discriminate a broad spectrum of species,9,25 (iii) has sufficient conservation to be less variable within than between species, and (iv) has sufficient phylogenetic information to assign species to major taxa. 25 To be effective, interspecific differences should be clearly and preferably significantly greater than intraspecific differences.3,4,10
Mitochondrial DNA (mtDNA) offers several advantages over nuclear DNA as a barcoding marker. Nuclear DNA undergoes relatively slow mutation compared with mtDNA and would require a much longer nucleotide sequence than is necessary with mtDNA to differentiate species. 3 The mitochondrial genome contains 13 protein-coding genes: cytochrome c oxidase subunits 1, 2, 3; cytochrome b subunit, NADH dehydrogenase subunits 1, 2, 3, 4, 4L, 5, 6, and ATPase subunits 6 and 8, and also 2 ribosomal RNA (rRNA) genes (16S and 12S) and 22 transfer RNA genes. 26 Hebert et al. (2003a) proposed to use a 648 bp region from the 5′-end of the cytochrome c oxidase subunit 1 (COI) gene as a standard DNA barcode aiming at creating a universal system to identify species.9,10,12,27 They established that the 5′ section of this mitochondrial gene was sufficient to place species into taxonomic categories, at least from phyla to orders. 12 Moreover, the COI gene has proved to be suitable for the identification of a large range of animal taxa with a 95%–97% resolution.4,10,12,17,21,28–30
The use of 16S region as the universal barcode for amphibians
The number of new amphibian species discovered annually is increasing. 31 Paradoxically, however, amphibian populations are undergoing a drastic global decline due to anthropogenic influences such as habitat destruction and pollution but also due to diseases such as the fungus Batrachochytrium dendrobatidis.32,33 Many amphibian species are morphologically similar to others or have complex life cycles, with a larval phase radically different in morphology from the adult phase, which can only be differentiated by genetic analysis. This complicates the identification and description based on their morphology alone. 20 DNA barcoding is clearly one tool that could be used for validating species identity and has been applied in several studies to successfully distinguish between different species of tadpoles that look morphologically similar.20,34,35
Skepticism about the COI-gene as a standard DNA barcode has been expressed. 36 In relation to amphibians, COI has not been used in a large number of phylogenetic or phylogeographic studies,37–39 and rarely as the only marker. The main problems with using COI as a DNA barcode in amphibians have been due to (i) the high variability of priming sites (limiting the robustness and repeatability of amplifying DNA sequences) and (ii) the overlap in divergence values observed within and between species, making it difficult to define threshold values and, therefore, identify species. 40 Since the COI barcode seems to not perform as efficiently for amphibians as for other groups of animals, alternative genes have been sought. 41
Vences et al. (2005) and other researchers42,43 suggested the use of the 16S rRNA gene as DNA barcoding marker in amphibians. Although many amphibian species are relatively old entities 44 variable regions within the 16S gene, corresponding to loops in the rRNA structure, ensure a sufficient amount of mutation is observed among species to allow their identification. Previous results in amphibians, fishes, reptiles, and mammals indicate that 16S is sufficiently variable to unambiguously identify most species. 25 Specifically in amphibians, an amplification success of 100% was recorded for 16S in a subset of fresh and well-preserved samples of mantellid frogs from Madagascar, while various combinations of COI primers had lower success rates (50%–70%).25,40 Interspecific pairwise 16S divergences were at a level suitable for assignment of larval stages to species (1%–17%), with low degrees of pairwise haplotype divergence within populations (0%–1%).25,40 Since amplification of 16S has also been successful for amphibians in other studies,35,43,45 it seems likely that 16S rRNA could be more effective as a standard DNA barcoding marker than COI for amphibians.
Zoological Society of london/Amphibian Ark/Frozen Ark Project
Some 20.64%, 12.37%, 22.10%, and 30.26% of mammal, bird, reptile, and amphibian species are facing a high risk of extinction in the wild over the next 10 years (www.iucnredlist.org/). For this reason, the Zoological Society of London (ZSL), the Natural History Museum, and the Institute of Genetics at the University of Nottingham established The Frozen Ark Project. The Frozen Ark project is a biobank that collects, preserves, and stores genetic resources such as tissue, gametes, viable cells, and DNA of the world's endangered animal species. It is a global reference collection for research and conservation. Furthermore, frozen viable cells may enable conservation biologists to help abate the dangerous loss of genetic variation that can cause infertility and early death in populations of endangered species. If cells are preserved, invaluable information about the species is saved (www.frozenark.org).
Amphibians are an important component of the global ecosystem, as indicators of environmental health and contributors to human health. The Amphibian Ark (AArk) was formed as a part of the Amphibian Conservation Action Plan (www.amphibianark.org). It is a joint effort of three principal partners: the World Association of Zoos and Aquariums, the IUCN/SSC Conservation Breeding Specialist Group, and the IUCN/SSC Amphibian Specialist Group and seeks to select species to be maintained in captivity that would otherwise go extinct. Both the AArk and the Frozen Ark project recognize biobanking amphibian samples as a vital back-up strategy to these ex situ conservation programs that are only likely to save a small proportion of the species threatened with extinction. This need prompted the development of the ZSL/AArk cryobank that contains 21 amphibian species presently. 46
It is necessary to verify the identity of the species stored within biobanks. This is especially important with regard to amphibians due to the difficulty in distinguishing between species phenotypically, particularly during the larval stages (as mentioned earlier). Verifying samples within a biobank belong to a particular species is also vital if they are to be used for research and/or breeding purposes. In verifying the specific identity of samples within biobanks, a large amount of genetic information on species can also be collected, potentially useful for establishing the genetic variability and phylogeny of populations and/or species.
Traditional barcoding procedure
The DNA barcoding procedure permits a rapid acquisition of molecular data compared with the morphological approach, which is time consuming, complicated, and sometimes totally confusing or almost impossible. 9 The traditional DNA barcoding technique includes DNA extraction from the sample specimen, amplification of the DNA by using a PCR, and, finally, DNA sequencing. The resulting sequence data are then used in (i) a simple Basic Level Alignment Search Tool (BLAST) approach where a raw similarity score is used to determine the nearest neighbor to the query sequence, 47 and (ii) to construct a phylogenetic tree using a clustering method, such as distance-based neighbor-joining (NJ).4,10,12,17,48
The use of melting temperature analysis
Real-time PCR using 16S primers and an intercalating dye such as Sybr Green I coupled with melting temperature analysis could also be suitable for species identification. When Sybr Green I binds to double-stranded (ds)-DNA, it emits a fluorescent signal, which is detected and recorded by the real-time PCR instrument. Sybr Green I binding to ds-DNA is reversible. Consequently, melting PCR products that are double stranded at the end of the real-time PCR run result in a rapid loss of Sybr Green I binding and fluorescence as single-stranded-DNA is formed; this again is detected and recorded by the real-time PCR machine. 49 Melting temperatures of PCR products are determined not only by their length but also by their base-pair composition. Although the length of each amphibian 16S fragment should be the same, the base-pair composition between species should differ sufficiently to make distinguishing between them using melt temperatures possible. This method has been successfully used for genotyping and mutation scanning in humans.50,51
The DNA barcoding procedure is less time consuming than morphological taxonomy, but the combination of real-time PCR and melt temperature analysis could prove a cheaper and less time-consuming option than the traditional DNA barcoding, mainly due to the omission of the amplicon purification and sequencing steps. However, a relationship between the 16S fragment melting temperatures and the DNA sequence needs to be found to validate the approach. Consequently, the main objectives of this study were (i) to determine whether the 16S rRNA gene is a suitable DNA barcode for verifying the specific identity of amphibian samples (both DNA and tissue) stored within the ZSL/AArk cryobank and (ii) to determine whether a relationship between 16S fragment melting temperature and sequence exists, such that the traditional barcoding procedure could be replaced by a cheaper and less time-consuming alternative in the future.
Materials and Methods
All reagents and equipment were obtained from Applied Biosystems unless stated otherwise.
DNA extraction
Total DNA was isolated from the of 20 amphibian samples (individuals) present within the ZSL/AArk cryobank consisting of 13 species (Table 1) using the Dneasy Blood and Tissue kit (Qiagen Ltd.) according to the manufacturer's instructions.
PCR, polymerase chain reaction.
Conventional PCR analysis
Each PCR was performed in 50-μL reactions containing 200 ng of total DNA, 1×PCR Buffer (Bioline), 1.5 mm MgCl2 (Bioline), 0.8 mm dNTPs (Bioline), 0.5 μm each of primers 16S-H (5′-CCGGTCTGAACTCAGATCACG-3′), and 16S-L (5′-CGCCTGTTTACCAAAAACAT′3′ 25 ) and 2.5 U BioTaq DNA polymerase (Bioline). Reactions were run on the GeneAmp PCR system 9700 at 95°C for 5 min, followed by 35 cycles of 94°C for 30 s, annealing temperature 53°C for 30 s and 72°C for 30 s and a final 7 min at 72°C. Primers 16S-H and 16S-L were designed to amplify a 580-base pair region of the mtDNA 16S rRNA gene. 25 PCR products were resolved on 2% agarose gels at 100 V for 1 h, and product size was checked against a 100-base pair DNA ladder (New England Biolabs). Images of gels were acquired using a CCD camera (SynGene Gene Genius system; Synoptics Ltd.) and SynGene GeneSnap software.
DNA sequencing
Briefly, the 16S PCR products were purified using the QIAquick® PCR purification kit (Qiagen Ltd), according to the manufacturer's instructions, diluted to 20 ng/ul, and sequenced using the 16S-H and 16S-L primers (each diluted to 10 mm), the ABI BigDye® Terminator cycle sequencing kit v3.1, and the ABI PRISM 3730xl. 16S sequences were then compared with nucleotide sequences in the Blast database using the Blast search (www.ncbi.nlm.nih.gov/entrez/BLAST/) to find the nearest neighbor sequence. Nucleotide variability between 16S sequences was scored using the default settings of ClustalW2.52,53 A phylogram of the 16S sequences was generated using the default NJ method settings of ClustalW2. 48
Real-time PCR and melt temperature analysis
Each PCR was performed in 14-μL reactions containing 7 ng of total DNA, 7 μL SYBR® Green PCR Master Mix, 0.9 μM of each 16S-primer, and 5.1 μL ddH2O. Reactions were performed in a StepOnePlus™ Real-Time PCR System (software version 2.1), and the conditions were one hold step at 95°C for 10 min followed by 40 cycles of 95°C for 15 s, 65°C for 30 s, and 72°C for 30 s. At the end of the PCR, products were melted by increasing the temperature from 60 to 95°C at 0.5 °C increments. All total DNA samples were analyzed in duplicate.
Results and Discussion
Previously, the DNA barcoding technique had been used as a tool for validating bird species identity in biobanks. 54 This procedure uses a single-gene sequence from a part of the genome that has a species-specific identity and can be sequenced across many species using a single primer pair. 10 Many amphibian species are morphologically similar to each other or have complex life cycles with radically different morphology at different stages that can only be differentiated by genetic analysis. 20 For this reason, several different DNA samples from individuals of known amphibian species were amplified with 16S primers, and the amplicons generated were analyzed by both DNA sequencing and melting temperatures. This allowed the performance of each method in validating the specific identity of amphibian samples in biobanks to be compared. DNA from twenty individuals consisting of thirteen different amphibian species were analyzed using the two methods outlined.
As previously reported,20,25,35,40 100% of frog and toad samples were successfully amplified and then validated to the species or genus level using conventional PCR and DNA sequencing with 16S primers (Table 2). The two species that were validated to the genus level (i.e., the Golden Mantella Frog and Amazon Milk Frog) appear to have high similarity with other species in the BLAST database. Sharing of similar 16S sequences among different but related species that occupy similar geographical ranges (e.g., Mantellinae are found only on Madagascar and Trachycephalus species in Table 2 occupy the northern regions of South America) could be due to incomplete lineage sorting or introgression.25,55
However, amplification of urodele DNA was not possible with the 16S primers (Table 1); high variability between the 16S primers and urodele 16S sequences could account for this observation. Indeed, pairwise comparisons between the 16S primers urodele sequences 16S sequences (i.e., Smooth Newt and Palmate Newt) available in Genbank confirm the variability is high (80% and 85% respectively). This finding is also consistent with the results recently obtained by Xia et al. 56 where COI was more suitable as a DNA barcode than 16S for salamanders.
ClustalW2 analysis using the 16S sequences produced a phylogenetic tree that contained two major Neobatrachian groups: Ranoidea and Hyloidea (Fig. 1). However, several inconsistencies between this tree and the most recent classification and phylogeny of amphibia 57 were found: the Non-Neobatrachian group was not recovered, thus Alytes muletensis (member of the Alytidae group) was placed erroneously among the Ranoidea; Phyllomedusa bicolour (member of the Hylidae group) formed its own major group when it should have been placed among the Hyloidea; Mantella aurantiaca and Theloderma corticale should have appeared more closely related to each other than to the Ranids, but our analysis suggested T. corticale and the Ranids were more closely related to each other than M. aurantiaca; and finally, Bufo bufo and the Leptodactylinae should have appeared more closely related to each other than to Trachycephalus resinifictrix, but our analysis suggested the latter species and the Leptodactylinae were more closely related to each other than B. bufo. Nonetheless, the aim of this study was to validate specific identity of samples rather than ascertain their phylogenetic relationship. To do the latter effectively clearly requires additional mtDNA (e.g., Cyt b and 12S rRNA) and several nuclear DNA markers to be employed (as described in 57 ).
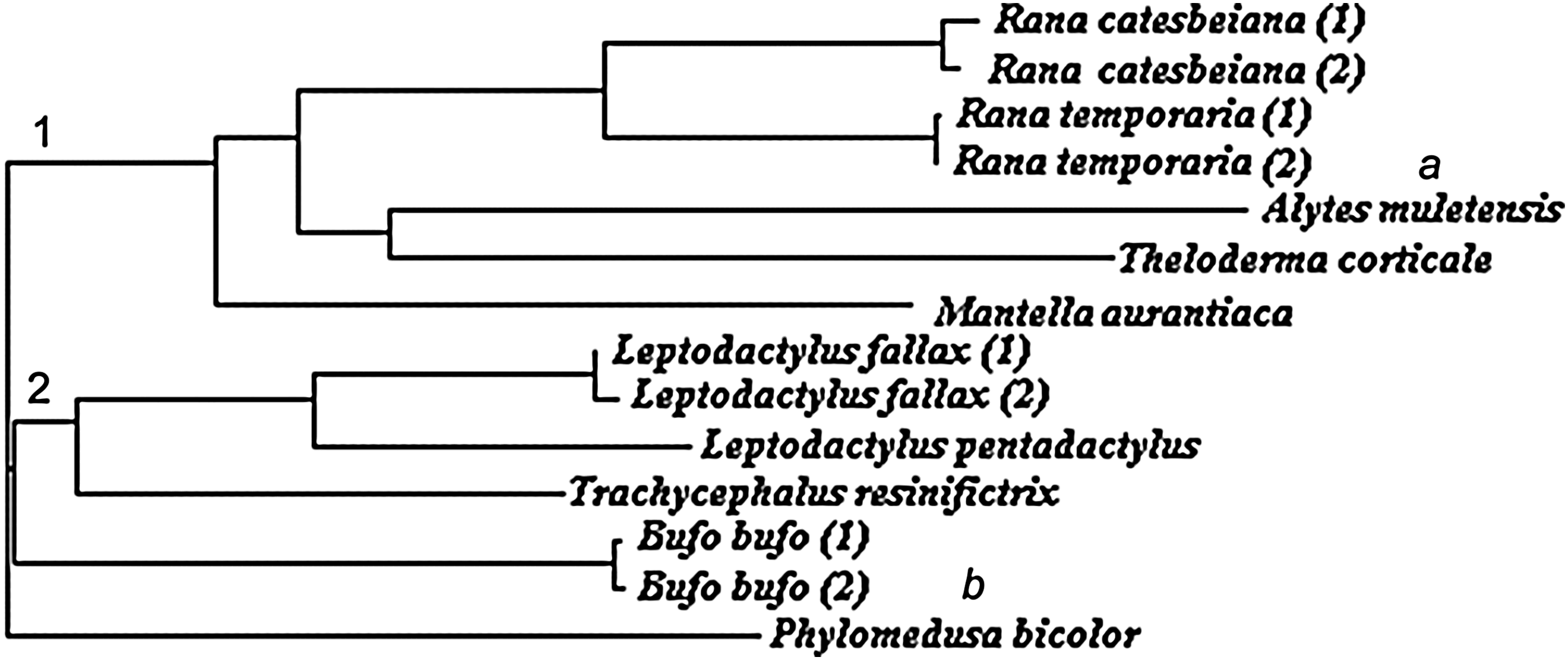
Neighbor-joining tree of the biobanked amphibian samples based on 16S sequences. Numbers indicate two major Neobatrachian groups: Ranoidea (1) and Hyloidea (2). Letters indicate inconsistencies with the most recent amphibian classification and phylogeny (57): Alytes muletensis should form its own branch, as it is a non-Neobatrachian (a), while Phyllomedusa bicolor should form a part of the Hyloidea group (b).
Melting temperatures were also obtained for the frog and toad (but not urodele) samples after real-time PCR with 16S primers (Table 1). Different melting temperatures between different species were expected. However, a small variability in melting temperature was found between two different Common frogs (81.09°C vs. 81.34°C). The presence of a single nucleotide polymorphism between the two 16S amplicons could have been responsible for this finding but was not evident from the sequences (Fig. 2). The small difference between the two frogs, therefore, was either due to temperature variability between wells, which can be as much as +/−0.5°C using the StepOnePlus Real-Time PCR system, even if the same DNA sample is analyzed, or due to slight DNA or salt (Mg2+) concentration variability between wells, caused by slightly differential pipetting, which can affect the Sybr Green I dye during melting. 50 Unexpectedly, we also found the same melting temperature (81.09°C) in three different species: Common Toad, Common Frog (individual 1), and Amazon Milk Frog, even though the 16S sequences of these frogs were highly variable (82%–87%). This finding highlights discordance between 16S melting temperatures and sequences, at least using the Sybr Green I dye and StepOnePlus Real-Time PCR System. However, the potential of melting temperature analysis for cheaper and faster species validation of samples in biobanks should not be discounted, just that DNA intercalating dyes (e.g., LCGreen) and melting platforms (e.g., HR-1, Idaho Technology) with much higher resolution than those employed in the current study might be required.50,51

Comparison of 16S sequences obtained from biobanked amphibian samples. Asterices indicate regions of similarity. Common Frog 1 and 2 share identical 16S sequences and a small variability (0.2°C) in melting temperature. Common Toad, Common Frog (individual 1), and Amazon Milk Frog 16S sequences are highly variable (82%–87%) but share the same (81.09°C) melting temperature, showing that 16S sequences and melting temperature are poorly correlated.
In conclusion, the 16S rRNA gene is a suitable DNA barcode for verifying the specific identify of frog and toad (but not newt) samples within biobanks. However, since there is no relationship between 16S sequence and melting temperature using the current system, further research into alternative higher resolution systems is required to validate whether melting temperature analysis with 16S primers can be employed as a cheaper and faster alternative to the traditional barcoding procedure.
Footnotes
Acknowledgments
Maya-Soriano MJ was supported by a PIF Grant from the Universitat Autonoma de Barcelona, Barcelona, Spain. The stay in ZSL was supported by PIF Grant “Ajuts per a estades de curta durada fora de Catalunya per a l'any 2010 pels becaris de les convocatòries PIF de la UAB” (ESTPIF2010-39). Lloyd RE was supported by a Leverhulme Early Career Research Fellowship grant (ECF/2009/0123).