
Abstract
The objectives of this study were to determine: 1) the optimal concentration (1.0 or 1.5 M) and duration of exposure (5, 10, or 20 min) of ovarian tissue to 1,2-propanediol (PROH) on morphology and viability of caprine preantral follicles; and 2) the effect of supplementing cryopreservation medium supplementation with Trolox® (0.1, 0.5, or 1.0 mM) or catalase (5, 10, or 20 IU/mL) on follicular morphology, viability, and lipid peroxidation. Cryopreservation decreased (p<0.05) percentages of normal follicles relative to the control (84%). Although supplementation of the cryopreservation medium (1.0 M PROH) with catalase (10 or 20 IU/mL) or Trolox® (0.1 mM) resulted in follicular morphology and viability similar to that in the controls (P>0.05), lipid peroxidation was reduced only when 20 IU/mL catalase was added to the cryopreservation medium.
Introduction
It is well known that exposing cells to cryoprotectants causes oxidative stress.13–15 Additionally, exposure to cold or freezing stress damages cell membranes through lipid peroxidation (LPO), which can be evaluated by measuring malondialdehyde (MDA). 16 Therefore, freezing medium supplementation with antioxidants have improved survival rates of hematopoietic stem cells 13 and hepatocytes.14,15 Recently, it was reported that supplementation of the freezing medium with antioxidants such as Trolox®, with or without catalase, reduced LPO in ram semen. 17 Even though ovarian tissue cryopreservation has been widely studied, there is a paucity of information regarding supplementing the medium with antioxidants. We recently reported that ascorbic acid preserved follicle morphology, but not viability of vitrified ovine ovarian tissue. 18 However, there are apparently no reports regarding the use of other antioxidants, for example, Trolox® and catalase (in association with PROH) in a slow-freezing protocol. In the present study, morphology and viability of preantral follicles were assessed after freezing using 1.0 or 1.5 M PROH for varying durations (5, 10, or 20 min), with or without the addition of catalase or Trolox®.
Materials and Methods
Chemicals
All chemicals and media used in this study were purchased from the Sigma Chemical Company (St. Louis, MO). For all of the media, the pH was adjusted to 7.4.
Source of ovaries
Ovaries (n=28) from adult mixed breed goats (n=14) were obtained at a local abattoir. These ovaries were washed once in 70% alcohol, twice in Minimum Essential Medium Eagle (MEM; Sigma), and transported to the laboratory (within 1 h after recovery) in thermoflasks at 20°C. From each ovarian pair, cortical fragments (∼3 mm3) were prepared as follows.
Histological analysis
Five pairs of ovaries were dissected, with each pair yielding 13 fragments. One fragment (control) was subjected to routine histological examination, whereas the remaining 12 fragments were used for toxicity testing (i.e., exposure to PROH, n=6), or cryopreservation (n=6). For toxicity testing, MEM+ [MEM supplemented with 10% heat-inactivated fetal calf serum (FCS; Laborclin, Pinhais, PR, Brazil)] was used as the holding medium, supplemented with 1.0 or 1.5 M PROH. Ovarian tissue exposure was performed in 2 mL macrotubes (MINITUB do Brasil Ltda., Porto Alegre, RS, Brazil) at 20°C for 5, 10, or 20 min. Immediately after exposure, ovarian fragments were submitted to cryoprotectant removal using a three-step wash (5 min each) in: (i) MEM++0.5 M sucrose, (ii) MEM++0.25 M sucrose, and (iii) MEM+. Subsequently, ovarian fragments were subjected to histological analysis. For freezing, six ovarian fragments were exposed to PROH and subjected to a cryopreservation protocol, as described. 19 The cryoprotectant was removed as described above, and ovarian fragments were fixed for routine histological analysis. 6 Preantral follicles were classified as: (i) morphologically normal when they contained an intact oocyte and intact granulosa cells; or (ii) degenerated when their oocyte nucleus had become pyknotic, the oocyte was shrunken, and granulosa cells were detached from the basement membrane.
Viability and lipid peroxidation analyses
To select the best exposure time, four pairs of ovaries were divided in four fragments. One fragment (control) was immediately submitted to follicular isolation and viability analysis, while the other 3 fragments were exposed to 1.0 M PROH for 5, 10, or 20 min, and cryopreserved. After thawing and cryoprotectant removal, ovarian fragments were submitted to follicular isolation and viability analysis. Based on the results obtained, the other five pairs of ovaries were dissected, with each pair yielding 15 fragments. One fragment (control) was immediately submitted to follicular isolation and viability analyses. The remaining 14 fragments (two fragments per treatment) were exposed for 5 min to 1.0 M PROH, with or without catalase (5, 10, or 20 IU/mL) or Trolox® (0.1, 0.5, or 1 mM). After exposure, ovarian fragments were cryopreserved as described above. For viability analysis, enlarged preantral follicles were mechanically isolated from ovarian tissue as described. 20 To each 100 μL of follicular suspension, 5 μL of 0.4% trypan blue was added and incubated for 5 min. 21 Follicles were scored as viable when unstained, and nonviable when stained with trypan blue, and the percentages of viable follicles calculated for each experimental condition. Treatments resulting in the best preserved follicular viability were assessed for lipid peroxidation. For lipid peroxidation analysis, tissue samples (∼140 mg) were individually homogenized in 50 mM Tris-HCl, pH 7.4 (1/5; w/v) and centrifuged for 15 min at 2400 g. Subsequently, an aliquot (300 μL) of the supernatant was incubated for 3 h at 95°C with a solution of 1.5 mL of 20% acetic acid containing 0.27 M HCl and 1.5 L of 0.8% aqueous solution of thiobarbituric acid (TBA), as described. 22 Absorbance was recorded using a plate reader at 532 nm to determine MDA concentrations.
Statistical analysis
All data were initially submitted to Kolmogorov–Smirnov and Bartlett's tests to confirm normal distribution and homogeneity of variance, respectively. Analysis of variance was then carried out using PROC GLM procedure of SAS (SAS Inst. Inc., Cary, NC) as follows: Regarding to histological analysis, a 2×2 factorial arrangements of treatments with PROH concentrations (1.0 M and 1.5 M) and procedure (only exposure and exposure followed by cryopreservation) as the main effects was applied. Ovarian fragments were considered as the experimental unit and the following model was used:
where Yijk=dependent variable (percentage of morphologically normal and viable preantral follicles), Ci=PROH concentration, Pj=procedure, Ci×Pj=interaction between PROH concentration and procedure, and eij=residual error. When any main effect or interactions were significant, means were separated by least significant difference. Concerning to viability and lipid peroxidation analyses, a randomized design of different concentrations of two antioxidants (catalase and Trolox®) was applied, being ovarian fragments also considered as the experimental unit. In all cases, differences were considered to be significant when p<0.05 and results were expressed as mean±standard deviation (SD).
Results
Histological analysis
Exposure to PROH, at both concentrations (1.0 or 1.5 M) and all exposure times (5, 10, or 20 min), resulted in similar percentages of morphologically normal follicles when compared to the control (84%; Fig. 1). However, after freezing, all treatments had decreased (p<0.05) percentage of normal follicles relative to the control. Although there was no difference between exposure time or PROH concentration on the percentages of morphologically normal follicles after cryopreservation, frozen-thawed ovarian tissue previously exposed to 1.0 or 1.5 M PROH for 20 min had a lower (p<0.05) percentage of normal follicles compared to tissue only exposed to PROH for 20 min. Figure 2 illustrates normal and degenerated preantral follicles before and after cryopreservation.
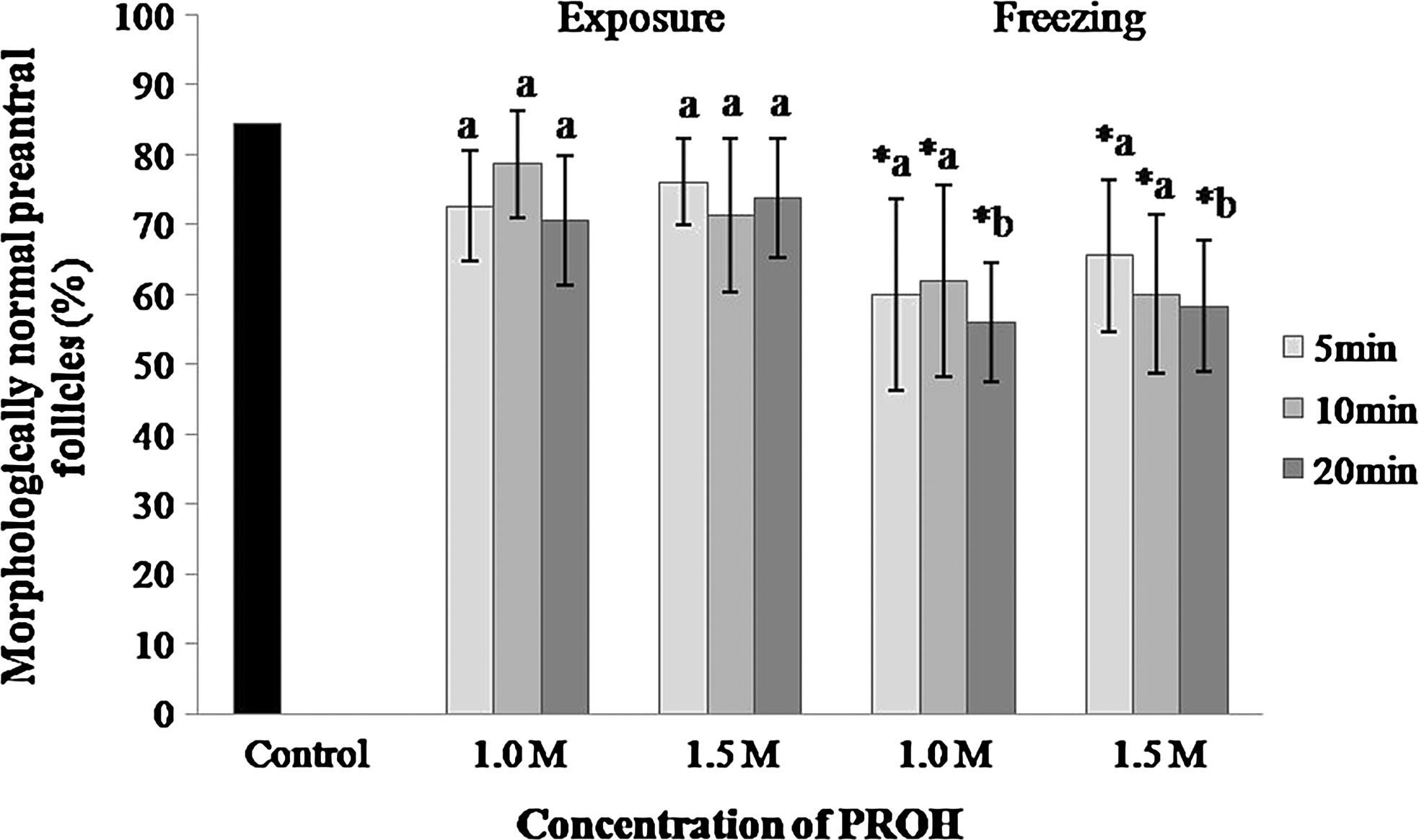
Percentage of morphologically normal preantral follicles enclosed in caprine ovarian tissue after exposure to PROH (1.0 or 1.5 M) or freezing for 5, 10, or 20 min. *Means with an asterisk differed from the control (p<0.05);

Representative images of morphologically normal
Viability and lipid peroxidation analyses
Based on histological examination, ovarian fragments were exposed to 1.0 M PROH for 5, 10, or 20 min and cryopreserved. Exposure to PROH for 5 or 10 min, followed by cryopreservation, resulted in similar percentages of viable follicles (60% and 62%, respectively), when compared to the control (79%; Fig. 3). However, exposure to 1.0 M PROH for 20 min decreased (p<0.05) viable follicles (46%).
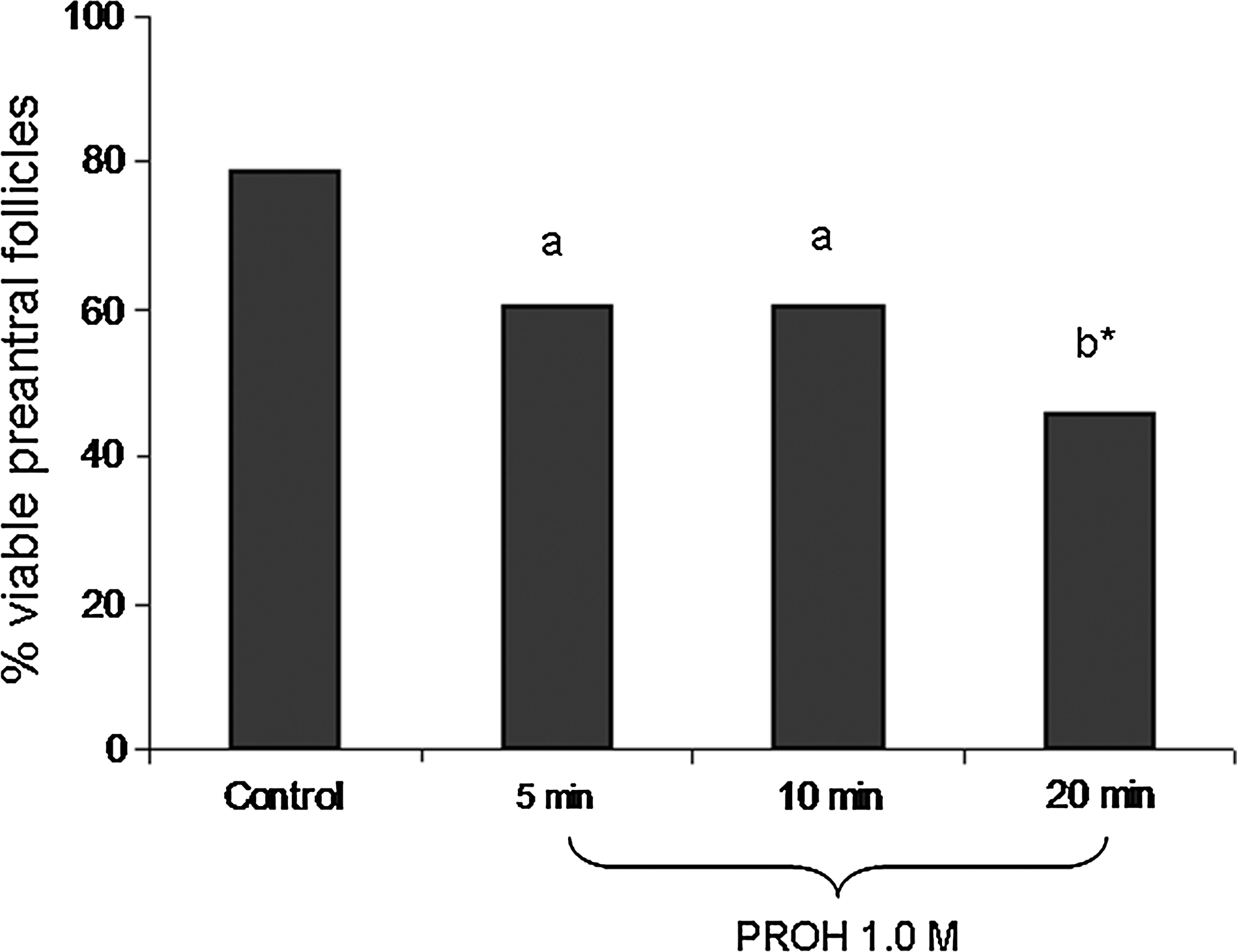
Percentage of viable preantral follicles after exposure to PROH 1.0 M for 5, 10, or 20 min, followed by freezing. *Means with an asterisk differed from the control, p<0.05;
Viability analysis of ovarian tissue cryopreserved in the presence or absence of catalase (5, 10, or 20 IU/mL) or Trolox® (0.1, 0.5, or 1 mM) is shown (Table 1). Percentages of viable follicles were similar (p>0.05) to the control (fresh tissue) when ovarian tissue was cryopreserved in a solution containing 1.0 M PROH with the addition of catalase (10 or 20 IU/mL) or Trolox® (0.1 mM). Conversely, cryopreservation in a solution containing 1.0 M PROH alone decreased (p<0.05) percentages of viable follicles relative to the control. Follicular viability was not preserved with catalase (5 IU/mL), but was preserved with the lowest concentration of Trolox® (0.1 mM). Ovarian tissue cryopreserved with PROH plus 20 IU/mL catalase had less MDA (p<0.05) than other treatments (Table 2). Furthermore, MDA values in cryopreserved ovarian tissue were not significantly different following cryopreservation without a cryoprotectant, or with PROH, with or without 10 IU/mL catalase or 0.1 mM Trolox®.
Differs from control; Ø differs from cryopreserved tissue without antioxidant (p<0.05).
Treatments without a common superscript differed, p<0.05.
Figure 4 illustrates viable and nonviable preantral follicles after trypan blue analysis.
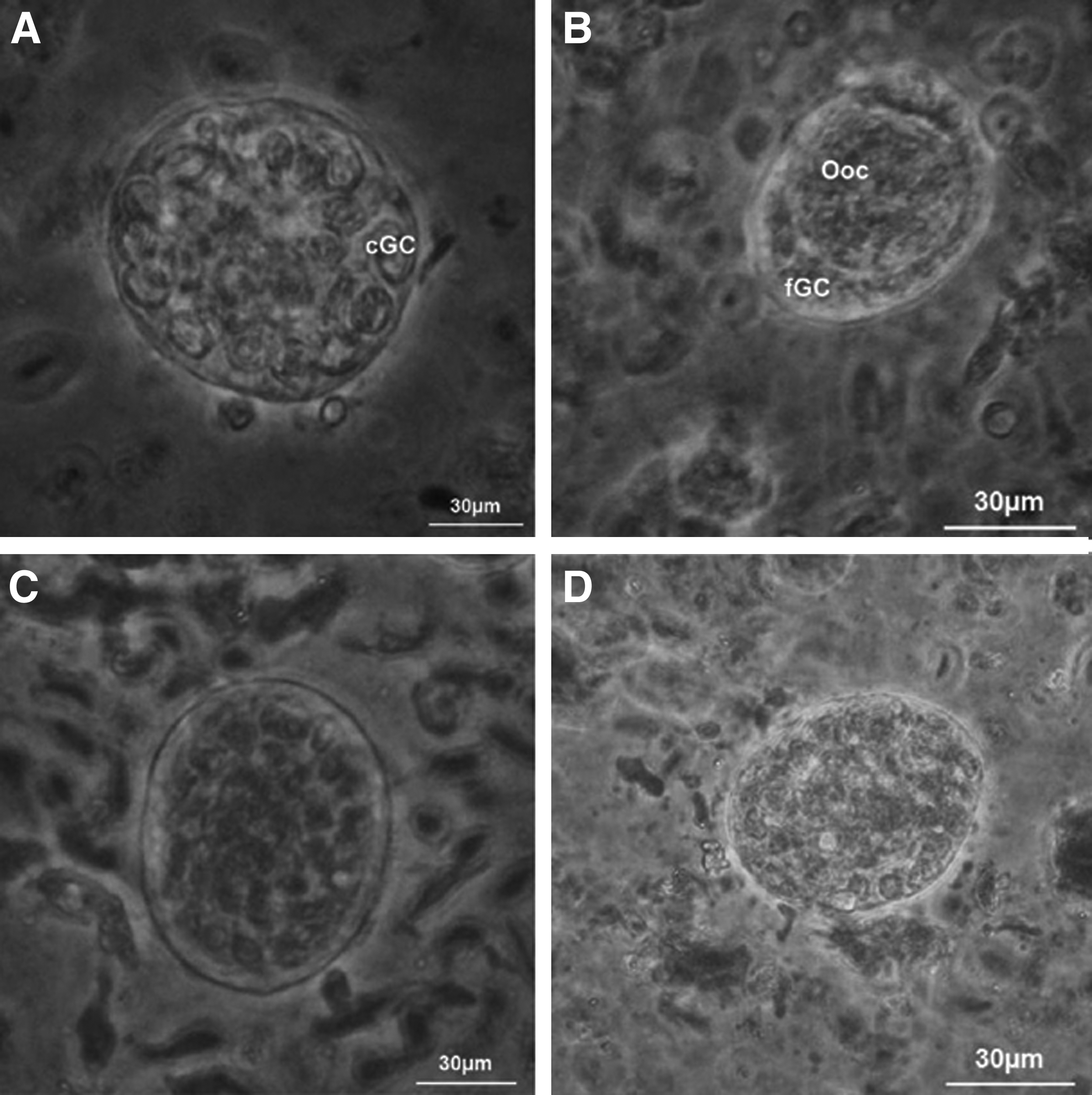
Representative images of viable
Discussion
Follicular morphology was not affected by the exposure to PROH, independently of the cryoprotectant concentration (1.0 or 1.5 M) and exposure time (5, 10, or 20 min). When tissue was exposed for 20 min to PROH, independently of its concentration, and subsequently cryopreserved, a negative effect of the freezing process was observed. It has been shown previously that ovine ovarian tissue exposed to 1.5 M PROH for 20 min had irreversible follicular damage after cryopreservation, 7 whereas isolated ovine primordial follicles were successfully cryopreserved after exposure to 1.0 M PROH for 5 min. 23 We concluded that prolonged exposure to PROH may lead to cell damage detectable only after freezing.
Percentages of morphologically normal follicles were increased when catalase (10 or 20 IU/mL) or Trolox® (0.1 mM) were added to a solution of 1.0 M PROH. Follicular survival, however, was impaired when Trolox® (0.5 or 1.0 mM) was added to the cryopreservation medium. Therefore, 0.5 or 1.0 mM Trolox® seemed to be toxic. In that regard, the concentration-dependence of Trolox® (0.1–0.8 mM) on mouse embryo development was tested, and it was found that Trolox® used at concentrations above 0.6 mM results in embryotoxicity. 20 Ovarian tissue cryopreserved with PROH plus 20 IU/mL catalase resulted in the lowest MDA levels. The present study highlighted the importance of catalase as a supplement in a PROH-based cryopreservation medium. In previous studies, PROH was either deleterious6,7 or ineffective 8 as a cryoprotectant. Therefore, cryopreservation of ovarian tissue from small ruminants has primarily used solutions containing dimethyl sulfoxide 24 or ethylene glycol.2,20 However, in the present study, cryopreserved ovarian tissue previously exposed to PROH for 5 min had MDA values similar to those in ovarian tissue frozen in the absence of cryoprotectant. Therefore, we inferred that exposure of ovarian tissue to 1.0 M PROH for 5 min before freezing was sufficient to maintain follicular morphology, but not to protect the tissue from cryoinjuries.
In conclusion, caprine follicular morphology was successfully preserved when ovarian tissue was exposed for 5 min to 1.0 M PROH before cryopreservation, with an enhanced preservation of follicular viability when freezing medium was supplemented with 0.1 mM Trolox® or 10 or 20 IU/mL catalase. However, 20 IU/mL catalase minimized lipid peroxidation.
Footnotes
Acknowledgments
This research was supported by CNPq and FUNCAP, Brazil. H. K. M. Luz is supported by a grant from CNPq. In addition, CNPq also supports APR Rodrigues (Project no. 471.739/2008-1), RR Santos (Project no. 483439/2009-6), and JR Figueiredo (Project no.554.812/2006-1).
Author Disclosure Statement
No competing financial interests exist.