
Abstract
The human brain displays oxidant and antioxidant markers with regional specificity that directly impinges on neuronal function in aging and in disease states. Similarly, the antioxidant activities might exhibit differential intracellular distribution rendering subcellular structures differentially vulnerable to toxic insults. To investigate the subcellular distribution of antioxidant activities in the human postmortem brain, we assayed superoxide dismutase (SOD), glutathione (GSH), glutathione peroxidase (GPx), glutathione reductase (GR), and glutathione-S-transferase (GST) in the cytosol and synaptosomal fraction from the frontal cortex (FC) of 45 postmortem human brains. We also tested whether these activities were altered by premortem and postmortem factors, including increasing storage time (11.8–104.1 months), postmortem interval (PMI) (2.5–26 h), age and gender differences, and agonal state [based on Glasgow coma scale (GCS): range: 3–15]. Overall, the antioxidant activities were found to be several folds lower in the synaptosomes compared to cytosol, which could make it more susceptible to degeneration. The activities were significantly affected mainly by age (SOD increased in synaptosomes, p=0.01; GSH decreased in cytosol, p=0.03; GPx decreased in cytosol and increased in synaptosomes, p=0.05; GST decreased in synaptosomes, p=0.05) and to a lesser extent by other premortem (GST decreased with GCS in synaptosomes, p=0.02) and postmortem factors (GSH decreased with PMI in cytosol, p=0.04). Increasing storage time or gender difference did not affect the antioxidant activities. We infer that premortem and postmortem factors in general, and increasing age in particular, significantly alter the antioxidant activities in subcellular fractions of postmortem brain with implications for studies on brain pathology employing stored human samples.
Introduction
Studies on markers of antioxidant function and oxidative stress in human material require use of postmortem brain tissues. Prior to embarking on utilizing postmortem human tissues for study of disease states, it is critical to determine if the preservation and subcellular distribution of antioxidant markers in the human brain could be influenced by premortem parameters (age, gender, medication received, and the agonal state) and postmortem parameters [postmortem interval (PMI), tissue preservation and duration, and the conditions of storage]. The effects of these factors on RNA stability, gene expression,13–19 and proteins 20 in the human brain have been studied, but their influence on antioxidant activities and relative distribution in subcellular compartments have not been elucidated. We have demonstrated that glutathione (GSH) metabolism and markers of oxidative damage and astrogliosis are affected by premortem and postmortem factors.21,22 Similarly, if the status of antioxidant markers in the subcellular compartments of the human brain is altered independent of disease-related changes, this could influence the interpretation of the experimental results.
We analyzed the status of antioxidant markers in cytosol and synaptosomes of the frontal cortex (FC) in postmortem brains (n=45) stored at the Human Brain Tissue Repository (HBTR), National Institute of Mental Health and Neurosciences (NIMHANS), Bangalore, India, and determined the effects of increasing PMI (2.5 to 26 h), age (2 days to 80 years), gender, agonal state [based on Glasgow coma scale (GCS): range: 3–15], and prolonged storage time (11.8–104.1 months) at−80°C.
Materials and Methods
All the chemicals used were of analytical grade. Bulk chemicals were obtained from Merck (Whitehouse Station, NJ) and Sisco Research Laboratories Pvt. Ltd. (Mumbai, Maharashtra, India). Fine chemicals and protease inhibitor cocktail were procured from Sigma (Eugene, OR).
Human tissue samples
Human brain samples were sourced from the Human Brain Tissue Repository (HBTR), a National research facility, Department of Neuropathology, National Institute of Mental Health and Neurosciences (NIMHANS), Bangalore, India. The brain tissues were collected following written informed consent from close relatives of the deceased to store and utilize the material for research. The Institutional Ethics Committee approval was obtained for the study protocol. The brains were from subjects who succumbed to road traffic accidents (nonalcoholics, nondiabetics, not on any medication, and with no known neurological or psychiatric disorders). Demographic and clinical details of all the subjects and the agonal state scored by the Glasgow coma scale (GCS) were collected from medical records. The GCS score of the subjects was based on (i) best eye response (E; range: 1–4) (ii) best verbal response (V; range: 1–5) and (iii) best motor response (M; range: 1–6) with a total GCS score range of 3 to 15 (noted at the time of hospital admission), with score of 3 corresponding to the most severe agonal state. The hospital stay following admission varied among the patients. During the hospital stay, assisted ventilation and parenteral administration of mannitol, hydrocortisone, and ionotropic drugs were administered to maintain the vital parameters whenever necessary.
Following the death of the patients, the PMI (the time interval between the time of death and placing the dissected brain slices into the freezer maintained at−80 °C) was recorded. Within one hour of death, routinely, the body was transferred to a refrigerator maintained at 2°C–4°C with a recorder and uninterrupted power supply. Following autopsy, the brains were recovered and sliced coronally and kept flat on salt-ice mixture (-15°C to−18°C) during dissection and then transferred in plastic zip lock bags into a box to be stored at−80°C in the HBTR. The procedure of dissection took ∼30–45 min and the brain slices transported on salt-ice mixture were transferred immediately into the deep freezer. The brain areas chosen for the study were anatomically farthest from the site of injury and without distinct edema or grossly apparent pathology. While the major portion of the tissue was frozen for biochemical studies, a portion corresponding to the mirror image of the stored tissue bits were fixed in buffered formalin. These tissues were subjected to histopathological assessment and the samples that maintained tissue integrity were utilized for the study (data not shown). The protocol of autopsy, tissue handling, and other procedures were uniform for all the samples. Such tissues have been earlier utilized extensively as control samples in PD research23–25 and other studies.3,20,21,26–28
In the current study, frontal cortex (FC) was analyzed from 45 postmortem brains (Table 1). The gender distribution (30 males; 15 females), age (range: 2 days to 80 y), PMI (range: 2.5–26 h), storage time (11–104.1 months), and agonal state (GCS score: 3–15) were noted in all the cases.
PMI, postmortem interval; GCS, Glasgow Coma Scale; NA, Not available.
Isolation of cytosol and synaptosomes from brain tissue 29
Approximately 200 mg of brain tissue was minced in 1000 μl of isolation buffer (350 mM sucrose, 5 mM TES, and 1 mM EGTA, pH 7.2) containing 100 μl of protease inhibitor cocktail and manually homogenized (16 strokes). The homogenate was centrifuged at 1000 g for 5 min at 4°C, and the supernatant was stored. The pellet was homogenized again (16 strokes) on ice and centrifuged (1000 g, 5 min at 4°C). The supernatants from both the steps were pooled and centrifuged at 8500 g for 10 min at 4°C to obtain the crude mitochondrial/synaptosome pellet. The post-mitochondrial supernatant that corresponded to the cytosol fraction was aliquoted and stored. The pellet was re-suspended in 300 μl of isolation buffer and overlaid on a discontinuous Ficoll gradient consisting of 6% (w/v Ficoll, ρ=1.065 g/ml, 3 ml), 9% (w/v Ficoll, ρ=1.075 g/ml, 2 ml), and 12% (w/v Ficoll, ρ=1.085 g/ml, 3 ml). The gradient was centrifuged at 75,000 g for 1 h at 4°C, and the synaptosomes which formed a whitish ring in the middle of the gradient were collected, resuspended in reconstitution buffer (250 mM sucrose and 10 mM TES, pH 7.2) and stored as aliquots in−80 °C. The purity and integrity of the isolated synaptosomes were confirmed by electron microscopy (TEM-Tecnai G2) (Fig. 1) as described previously (30). The cytosol and synaptosomes were utilized for the following biochemical experiments after protein estimation (31).
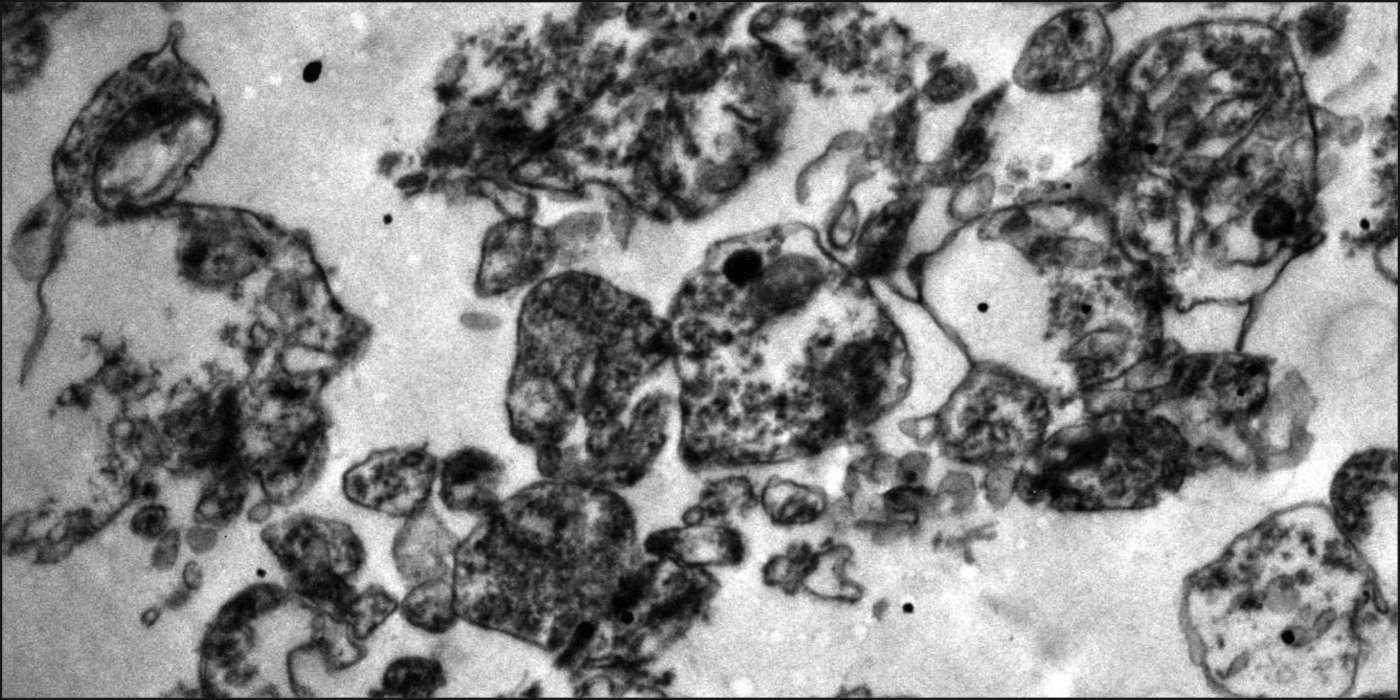
Representative electron micrograph showing synaptosomes (magnification=1X 13,000) purified from postmortem human brains samples (frontal cortex).
Superoxide dismutase (SOD) assay
SOD activity was assayed using its inhibitory action on quercetin oxidation based on the method described earlier, 32 with minor modifications. The final reaction mixture contained 30 mM Tris HCl (pH 9.1), 0.5 mM EDTA, 50 mM TEMED, 0.05 mM quercetin, and 10 μl of mitochondria containing 10 μg of protein. The reaction was monitored at 406 nm for 10 min. One unit of SOD activity was defined as the amount of enzyme (per mg protein) that inhibits quercetin oxidation reaction by 50% of maximal value.
Estimation of total glutathione (GSH+GSSG)
Total glutathione estimations in the brain extracts were carried out by the DTNB recycling method, as described earlier. 33 All estimations were conducted in triplicate, and total glutathione concentrations were normalized per mg protein.
Glutathione-S-transferase (GST) assay
GST was assayed by the 1-chloro 2-4-dinitro benzene (CDNB) method. 34 To 1 ml reaction mixture containing phosphate buffer (0.1 M, pH 6.5; 0.5 mM EDTA), CDNB (1.5 mM), and 50 μl GSH (1 mM), 30 μg protein (sample) were added and the increase in absorbance at 340 nm was monitored for 5 min. The enzyme activity was expressed as nmoles of S-2,4, dinitrophenyl glutathione formed /min/mg protein (MEC=9.6 mM-1cm−1).
Glutathione peroxidase (GPx) assay
GPx activity was determined by t-butyl hydroperoxide (tbHP) method. 35 The reaction mixture containing 150 μg protein (sample), phosphate buffer (0.1 M), 0.5 mM EDTA, 100 μl glutathione reductase (0.24 U), 100 μl GSH (1 mM), and 100 μl NADPH (0.15 mM) was incubated at 37°C for 3 min and the reaction was initiated by the addition of 100 μl tbHP (0.12 mM). Change in absorbance at 340 nm was monitored for 5 min spectrophotometrically and the activity was expressed as nmoles of NADPH oxidized /min/mg protein (MEC=6.22 mM−1cm −1).
Glutathione reductase (GR) assay
Solubilized brain protein extracts (100 μg) was assayed at 25°C in 0.1 M Tris–HCl (pH 8.1) and 0.2 mM NADPH, and the reaction was initiated by the addition of 1 mM GSSG and followed spectrophotometrically at 340 nm. 36
Statistical analysis
Quantitative data were accumulated from at least three independent experiments and expressed as mean±SD, followed by analysis of variance (ANOVA) and Pearson's correlation of linear regression (r value). In all the experiments, data with p<0.05 were considered to be statistically significant.
Results
In the current study, we investigated the distribution of antioxidant activities in the subcellular fractions of postmortem human brains and tested whether they are influenced by increasing storage time at−80°C (11.8–104.1 months), PMI (range: 2.5–26 h), age of the donors (2 days to 80 y), gender of the subject, and agonal state (GCS score: 3–15). This was carried out by assaying the SOD, GSH, GPx, GR, and GST in cytosol and synaptosomes fractions of FC from human brain samples (n=45).
The SOD activity was>5 fold lower in synaptosomes compared to the cytosol. The SOD activity was unaffected by storage time (cytosol: r=− 0.12, p=0.2; synaptosomes: r=− 0.10, p=0.29), PMI (cytosol: r=0.02, p=0.46; synaptosomes: r=− 0.11, p=0.24), agonal state (cytosol: r=− 0.12, p=0.23; synaptosomes: r=0.04, p=0.40), or gender (cytosol: p=0.89; synaptosomes: p=0.37) (Fig. 2). However, SOD activity increased with age in the synaptosomes, while it remained relatively unaltered in the cytosol fraction (cytosol: r=0.20, p=0.10; synaptosomes: r=0.37, p=0.01) (Fig. 2D).

Effect of premortem and postmortem factors on superoxide dismutase (SOD) activity in cytosol and synaptosomes of the frontal cortex (FC) region of human postmortem brains (n=45). SOD activity in cytosolic and synaptosomes from FC was assayed in triplicate, and the average for each sample was plotted as mean±SD followed by regression analysis. The r and p values are shown. Scatter plot graph with linear regression line for SOD levels with increasing
The total GSH content was ∼5 fold lower in the synaptosomes compared to cytosol. The total GSH content was unaffected by storage time (cytosol: r=0.21, p=0.11; synaptosomes: r=− 0.11, p=0.27), agonal state (cytosol: r=− 0.12, p=0.24; synaptosomes: r=− 0.23, p=0.09), or gender (cytosol: p=0.35; synaptosomes: p=0.15) (Fig. 3). However, total GSH decreased in the cytosol with increasing PMI (cytosol: r=− 0.28, p=0.04; synaptosomes: r=0.05, p=0.38) (Fig. 3B) and age, while it was unaltered in synaptosomes (cytosol: r=− 0.28, p=0.03; synaptosomes: r=0.11, p=0.24) (Fig. 3D).
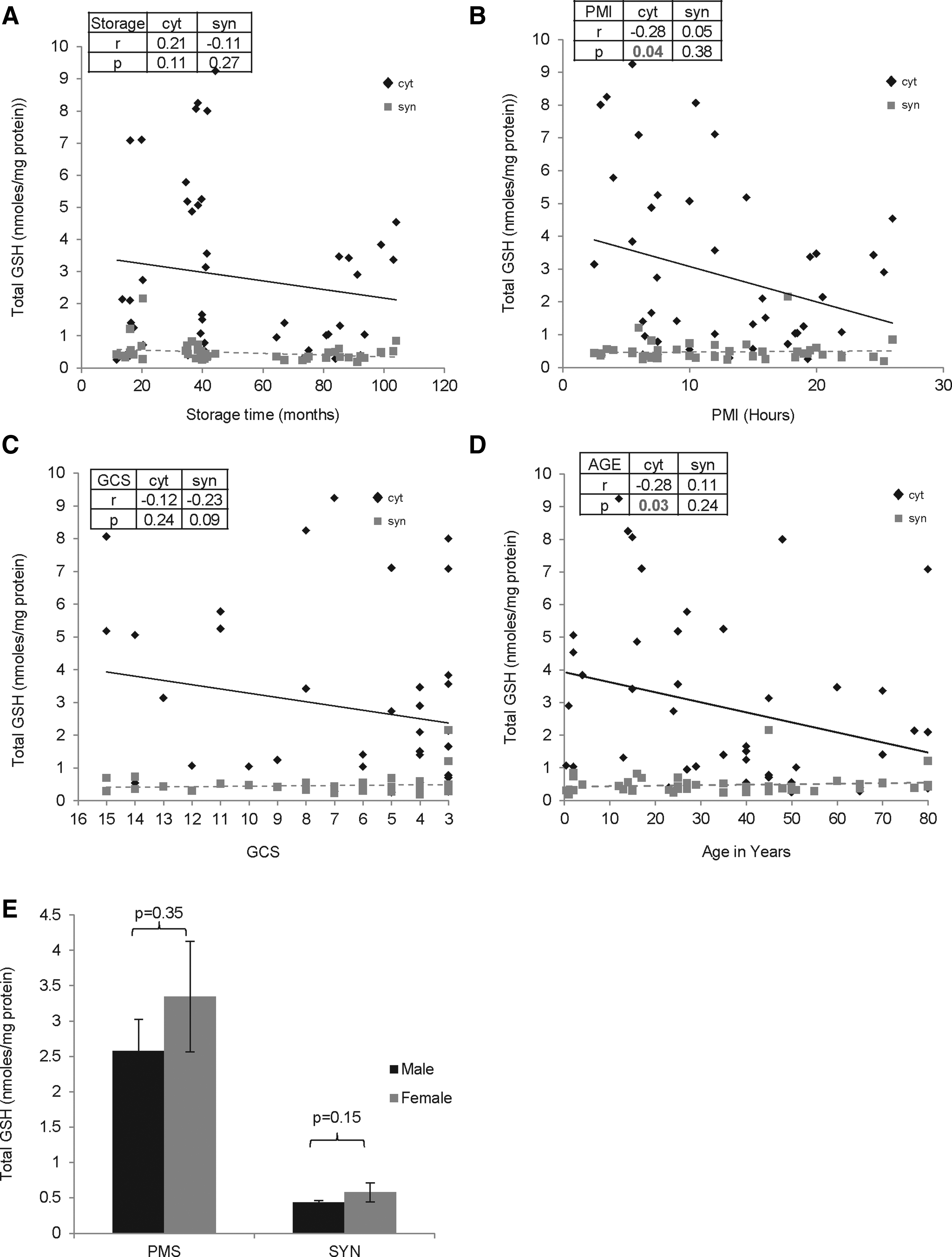
Effect of premortem and postmortem factors on the total glutathione (GSH) content in cytosol and synaptosomes of the frontal cortex (FC) region of human postmortem brains (n=45). Total GSH content in cytosol and synaptosomes from FC was assayed in triplicate, and the average for each sample was plotted as mean±SD followed by regression analysis. The r and p values are shown. Scatter plot graph with linear regression line for GSH levels with increasing
On the other hand, GPx activity was ∼ 8 fold lower in synaptosomes compared to cytosol. GPx activity was unaffected by storage time (cytosol: r=0.05, p=0.38; synaptosomes: r=− 0.15, p=0.18), PMI (cytosol: r=− 0.11, p=0.25; synaptosomes: r=− 0.11, p=0.24), agonal state (cytosol: r=0.13, p=0.23; synaptosomes: r=− 0.13, p=0.22) or gender (cytosol: p=0.93; synaptosomes: p=0.23) (Fig. 4). Interestingly, while the GPx activity decreased with age in cytosol, the activity significantly increased in synaptosomes (cytosol: r=− 0.26, p=0.05; synaptosomes: r=0.25, p=0.05) (Fig. 4D).

Effect of premortem and postmortem factors on glutathione peroxidase (GPx) activity in cytosol and synaptosomes of the frontal cortex (FC) region of human postmortem brains (n=45). GPx activity in cytosol and synaptosomes from FC was assayed in triplicate, and the average for each sample was plotted as mean±SD followed by regression analysis. The r and p values are shown. Scatter plot graph with linear regression line for GPx levels with increasing
The GR activity was ∼ 20 % lower in synaptosomes compared to cytosol. GR activity was unaffected by all factors including storage time (cytosol: r=0.11, p=0.27; synaptosomes: r=− 0.18, p=0.14), PMI (cytosol: r=− 0.13, p=0.20; synaptosomes: r=− 0.05, p=0.36), agonal state (cytosol: r=0.13, p=0.23; synaptosomes: r=0.19, p=0.13), age (cytosol: r=− 0.10, p=0.26; synaptosomes: r=− 0.08, p=0.30) and gender (cytosol: p=0.57; synaptosomes: p=0.11) (Fig. 5).
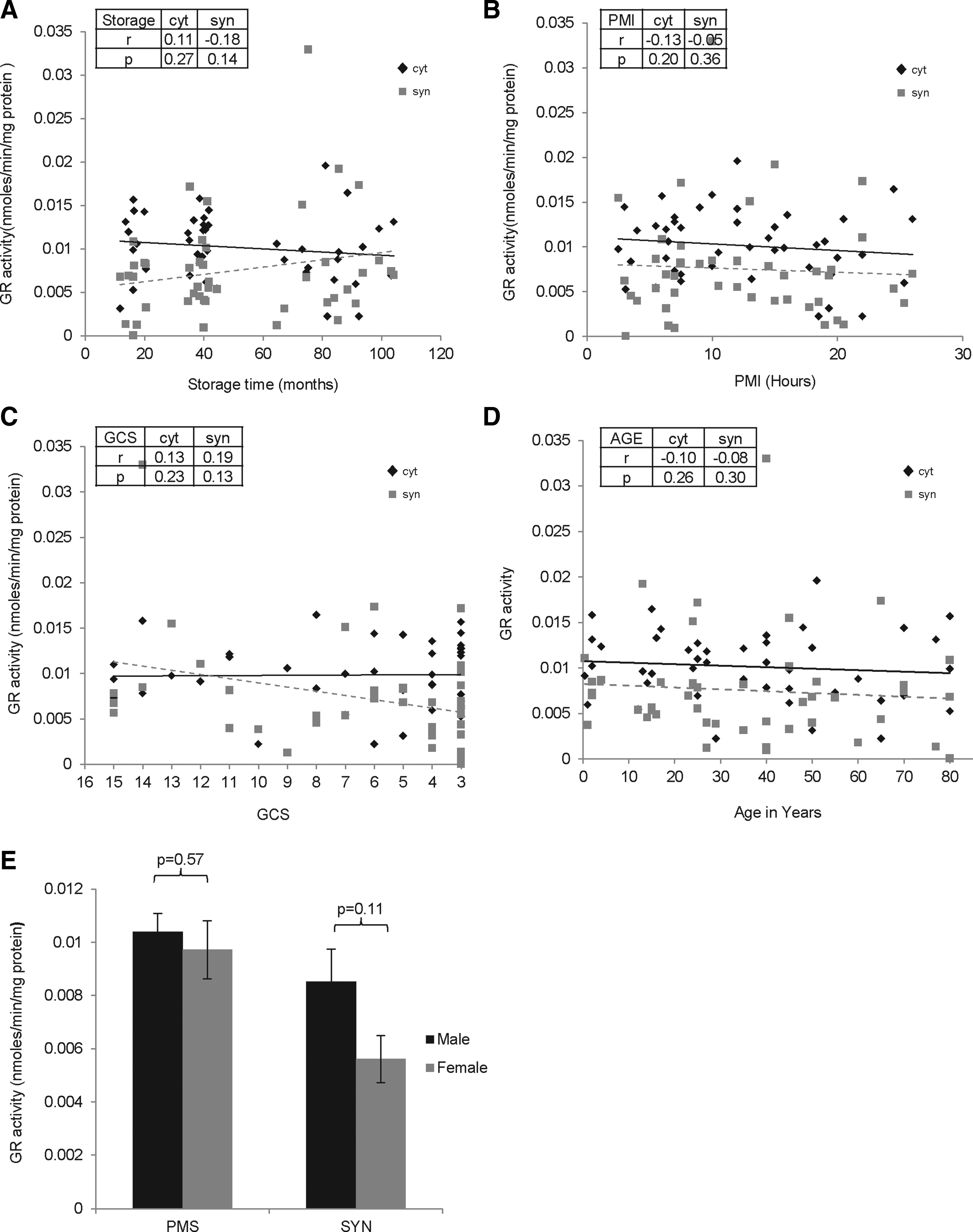
Effect of premortem and postmortem factors on glutathione reductase (GR) activity in cytosol and synaptosomes of the frontal cortex (FC) region of human postmortem brains (n=45). GR activity in cytosol and synaptosomes from FC was assayed in triplicate, and the average for each sample was plotted as mean±SD followed by regression analysis. The r and p values are shown. Scatter plot graph with linear regression line for GR levels with increasing
The total GST activity was>20 fold lower in synaptosomes compared to cytosol. The GST activity was unaffected by storage time (cytosol: r=0.07, p=0.34; synaptosomes: r=− 0.18, p=0.15), PMI (cytosol: r=− 0.01, p=0.47; synaptosomes: r=0.07, p=0.33), or gender (cytosol: p=0.72; synaptosomes: p=0.91) (Fig. 6). However, GST activity decreased in synaptosomes with agonal state (cytosol: r=0.12, p=0.23; synaptosomes: r=− 0.27, p=0.05) (Fig. 5C) and age, while it was unaltered in cytosol (cytosol: r=0.05, p=0.38; synaptosomes: r=0.32, p=0.02) (Fig. 6D).
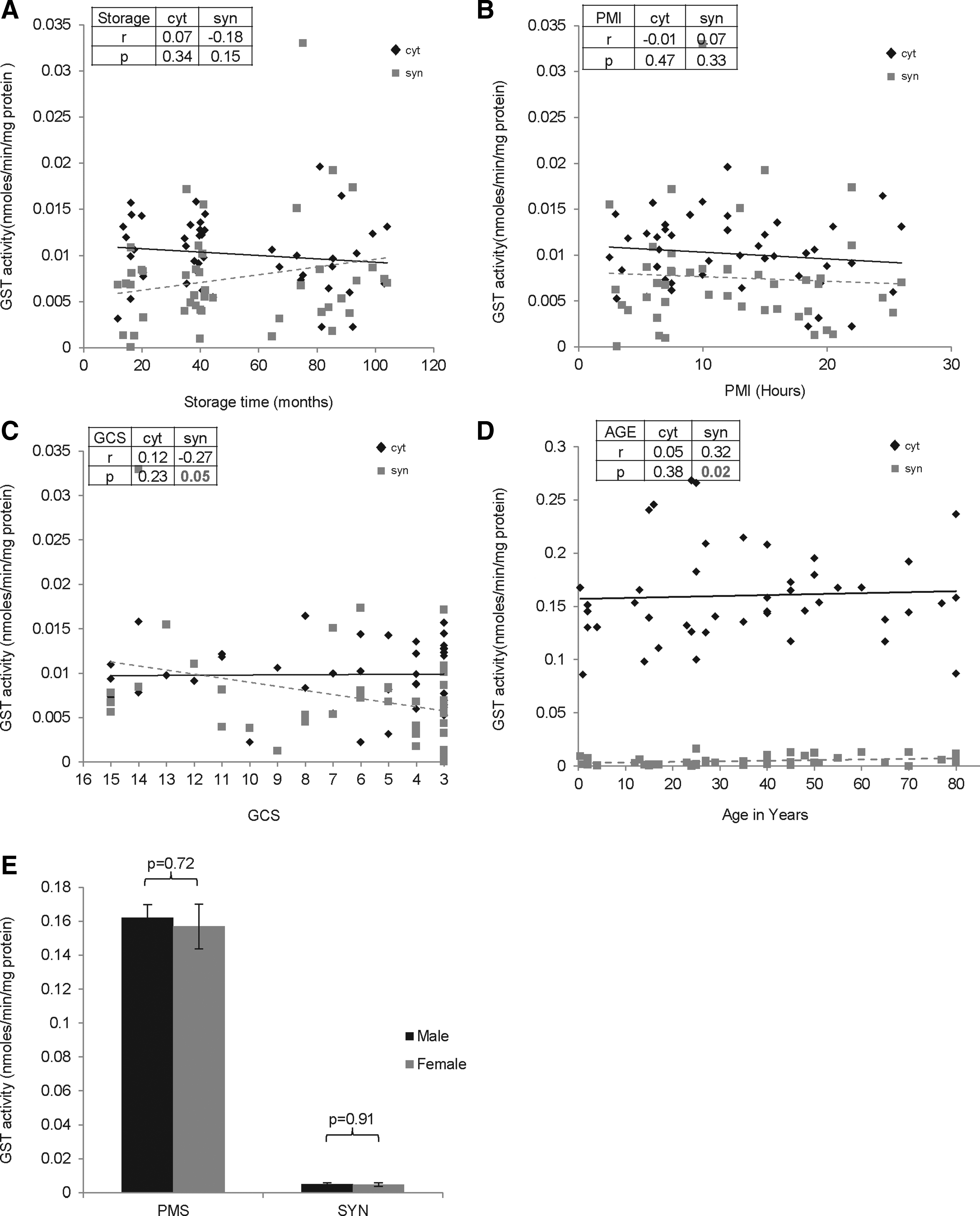
Effect of premortem and postmortem factors on glutathione-S-transferase (GST) activity in cytosol and synaptosomes of the frontal cortex (FC) region of human postmortem brains (n=45). GST activity in cytosol and synaptosomes from FC was assayed in triplicate, and the average for each sample was plotted as mean±SD followed by regression analysis. The r and p values are shown. Scatter plot graph with linear regression line for GST levels with increasing
Overall, we observed that the antioxidant activities were several fold lower in synaptosomes as compared to cytosol in the postmortem human brains. The activities were affected mainly by the age and to a lesser extent by other premortem and postmortem factors (Table 2).These results should be considered during interpretation of biochemical data in a meaningful manner.
m, mitochondria; c, cytosol (cyt); s, synaptosome (syn); SOD, superoxide dismutase; GSH, glutathione; GPx, glutathione peroxidase; GR, glutathione reductase; GST, glutathione-s-transferase; PMI, postmortem interval; GCS, Glasgow coma scale; ns, not significant.
Previous study (22); #Recent study (Harish et al, 2011, manuscript submitted); $Current study;
The r and p values for the parameters with significant change are indicated in boldface.
Discussion
The differential distribution of antioxidant markers in subcellular compartments must be studied to understand its influence on brain function and pathology. Among these markers, intracellular distribution of total GSH has been studied extensively in animal models. Schnellmann et al. 37 examined the intracellular distribution of total GSH in rabbit renal tubules and found that irrespective of the in vivo GSH levels in the tubules, the mitochondrial GSH pool was unaltered. Although the mitochondrial GSH concentration accounted for ∼10% of the total cellular GSH, depletion of total GSH in the tubules caused selective depletion of the cytosolic pool without affecting the mitochondrial GSH pool. Further,>40% loss of mitochondrial GSH caused mitochondrial dysfunction, reflecting complicated GSH dynamics between cytosol and mitochondria and their role in mitochondrial function. In a related study, Roychowdhury et al. 38 studied whether the mitochondrial and cytosolic GSH are differentially affected by endogenous nitric oxide (NO) in microglial cultures. Activated inducible nitric oxide synthase (iNOS) significantly depleted total GSH but the mitochondrial GSH was relatively unaffected. NO-mediated oxidative damage and mitochondrial dysfunction were more pronounced in cells where mitochondrial GSH was depleted compared to cytosolic GSH. Further, endogenous NO production did not compromise the mitochondrial GSH pool which, in turn, might explain the ability of these cells to resist the damaging effects of high-output NO production. Previously, we demonstrated that total GSH content in whole tissue extracts of FC from postmortem human brains was unaffected by premortem and postmortem factors. Similarly, total GSH in mitochondria (Harish et al, manuscript submitted) and synaptosomes (current study) in FC from human brains was unaffected by premortem and postmortem factors. In contrast, cytosolic GSH was significantly decreased with increasing age and PMI (Fig. 3). This is consistent with a previous study 9 which investigated the effect of GSH depletion on the viability of synaptosomes during brain aging in mice. Although aging did not influence the GSH and ATP levels and the viability of mouse synaptosomes, depletion of cytosolic GSH decreased the ATP levels and viability of synaptosomes in aged mice but not in young animals. These results emphasize the importance of the cytosol GSH for the maintenance of the membrane integrity of synaptosomes during brain aging in mice. The antioxidant activities in cytosol and synaptosomes were mainly affected by age and to a lesser extent by other premortem and postmortem factors. Gender had the least effect on antioxidant activities tested (Table 2). On the other hand, other antioxidant activities such as SOD, GPx, and GST were altered in the subcellular fractions with increasing age. We observed opposite trends in the GPx activity with increasing age, with declining activity in cytosol probably compensated by an increase in synaptosomes (Fig. 4D).
The current study also observed several fold lower antioxidant activity in synaptosomes compared to cytosol. All activities were≥5 fold higher in the cytosol compared to synaptosomes, while GR activity was ∼20% increased in cytosol. This also indicates the selective vulnerability of synaptosomes to oxidative damage with implications for pathogenesis of degenerative pathology. This was explained by a recent study, 39 which quantitated the distribution of oxidant and antioxidant markers in various subcellular (neuropil) fractions of FC from control, mild cognitive impairment (MCI), moderate AD, and advanced AD human brains. All the neuropil fractions (cytosol, mitochondria, and synaptosomes) demonstrated pathology-dependent elevation in oxidant markers and depletion of antioxidant activities, but the highest oxidative damage was observed in synaptosomes. This might be considered as the mechanistic basis underlying the selective targeting of synaptosomes during neurodegeneration.
Table 2 shows the distribution of antioxidant activities in whole brain extracts and different subcellular fractions in FC of postmortem human brains. Although the antioxidant activities in the whole extracts of FC remained unaffected by premortem and postmortem factors, the subcellular fractions revealed significant alterations. Hence the data from whole cell or tissue extracts should be used cautiously, as it may not accurately reflect the changes in subcellular structures. For instance, while the SOD activity was unaltered with increasing age in total extracts of FC, the activity increased with age in synaptosomes (Fig. 2D). Similarly, the GSH level which was unaltered in the total extracts of FC with increasing age and PMI was significantly decreased in the cytosolic fraction from the same samples indicating that this decrease was masked due to the other fractions (Figs. 3B and 3D). GPx activity in cytosol and synaptosomes was altered with age, while this was not reflected in the whole extracts (Fig. 4D). Similarly, the GST activity which was significantly increased in the whole extracts with increasing age was due to elevation only in the synaptosomes compared to the cytosolic and mitochondrial fractions (Fig. 6D and Table 2). In some instances, the activity in total extract could be correlated with the opposing trends in subcellular fractions. For example, GPx activity revealed decrease in cytosolic and increase in synaptosomal fraction with age, while total extracts showed the activity to be unaltered (Fig. 4D).
Interestingly, gender of the subjects did not influence any of the activities tested in the current study. In our previous study (Harish et al 2011, manuscript submitted), we observed that the activities of mitochondrial enzymes, such as citrate synthase and succinate dehydrogenase, were significantly higher in brains of female subjects compared to males. Gender of the subject might have significant effects on the neuronal function and synaptic transmission and this might be induced due to the action of hormones such as estrogen in the brain. There are different studies that have demonstrated that exposure to estrogen greatly influences the synaptic functions in the brain. For example, estrogen increases the concentration of neurotransmitters such as dopamine, serotonin, and norepinephrine, and modulates their release, uptake, and enzymatic inactivation. 40 Estrogen is also known to increase the receptors of these neurotransmitters. 41 Estrogens can modulate synaptic functions by inhibiting Na-dependent of Ca2+ efflux from the mitochondria. This causes mitochondrial retention of Ca2+, which modulates the synaptosomal Ca2+ content, thereby affecting the ion homeostasis in nerve terminals. 42 Further, estrogens have been described to induce synaptogenesis in the principal neurons of hippocampus and are shown to be synthesized and released by these neurons. 43
In conclusion, the current study combined with our earlier work 22 [Harish et al, manuscript submitted, and Venkateshappa et al, unpublished observations)], reveal a complex distribution of antioxidant activities in FC of postmortem human brain samples which could also differ in various anatomical areas. Further, these activities could be influenced by some premortem and postmortem factors, and the effect could be different in different subcellular fractions. Compared to cytosol, the synaptosomes showed several fold decrease in antioxidant activity which might make it vulnerable to oxidative stress and contribute to evolution of neurodegenerative diseases.
Footnotes
Acknowledgments
The authors gratefully acknowledge the Human Brain Tissue Repository (HBTR), Department of Neuropathology, NIMHANS, Bangalore, India for providing the human brain tissues for the study. The authors gratefully acknowledge Dr. N. Gayathri and Mrs. Hemavathy from the Electron Microscopy Laboratory, Department of Neuropathology, NIMHANS, for assistance in electron microscopy. The authors thank the donors and their relatives for the kind gift of human brains for neurobiological studies.
Disclosure Statement
The authors declare that there are no conflicts of interest.
This study was financially supported by the Indian Council of Medical Research (ICMR IRIS ID No. 2009-07710) (to MMSB). GH is supported by a senior research fellowship from ICMR, India. VC gratefully acknowledges the financial support from Siddhartha Medical College, Tumkur, India.