
Abstract
In order to create a bank of goat spermatozoa, it was important to evaluate the protective effect of five different cryoprotectant extenders. Additionally, this study determined whether the cryoprotective effect of trehalose is superior to that of other disaccharides. The data indicate that the motility (∼20%) and path velocity (∼15 μm/s) of spermatozoa frozen with extenders containing disaccharide were significantly less than that of spermatozoa frozen with extender containing no disaccharide (45.21±3.51% and 28.66±13.57 μm/s, respectively; p<0.01). Moreover, in the presence of disaccharide, the percentage of goat spermatozoa with exposed phosphatidylserine following cryopreservation was over 35% and significantly higher than spermatozoa frozen in the absence of disaccharide (4.88±3.41%, p<0.01). In addition, the presence of disaccharide did not increase the thermal resistance or protect membrane integrity of frozen spermatozoa. However, the percentage of cells with an intact acrosome following cryopreservation in the presence of disaccharide was significantly greater than that of cells frozen with no disaccharide (p<0.05). Finally, the protective effect of trehalose on frozen goat spermatozoa was similar to that of sucrose, maltose, or lactose (p>0.05). In conclusion, the effect of trehalose on frozen goat spermatozoa is not superior to that of other disaccharides including sucrose, maltose, or lactose. Moreover, disaccharide used as the main solute does not improve the quality of frozen goat spermatozoa except for protection of acrosome integrity. The role of disaccharide on frozen goat spermatozoa still needs further elucidation.
Introduction
The role of sugar is critical for survival of goat spermatozoa during cryopreservation.1,7 Generally, glucose or fructose is a necessary component of freezing extenders which are used as the primary substrate for glycolysis of goat spermatozoa.1,8,9 In addition, disaccharides can also be used for cryoprotection of goat spermatozoa. However, disaccharides act primarily as the extracellular cryoprotectant. Currently, trehalose is thought to be the most effective disaccharide.7,10–20 Aboagla and Terada found that cryoprotection of goat spermatozoa correlated with enhancement of membrane fluidity caused by trehalose. 7 However, other disaccharides, such as sucrose or lactose, can also be used for cryoprotection of mammalian spermatozoa.8,15–17,19–22
The exact mechanism by which disaccharides protect spermatozoa during cryopreservation is currently unknown. Disaccharides can create an extracellular hypertonic environment that can induce cell dehydration and decrease lethal intracellular ice formation. 1 According to the “water replacement hypothesis”, disaccharides may replace the water shell of macromolecules through linkage by hydrogen bonding and prevent injury caused by freezing or drying.23–25 Similarly, disaccharides may interact with the phospholipids of the plasma membrane, reorganizing the membrane and allowing spermatozoa to survive through the freezing process.7,12,22 Finally, disaccharides may insert into the membrane and limit excessive dehydration of spermatozoa during freezing and thawing, so that physical damage due to cell volume changes is decreased. 26
In order to select an optimal freezing extender for creation of a goat spermatozoa bank and to determine whether the cryoprotective effect of trehalose is superior to that of other disaccharides, five different freezing extenders were used to cryopreserve goat spermatozoa using a newly developed freezing method. Yunling black goats, a local breed of Yunnan province, were used as the source of semen. The motility, thermal resistance, acrosome integrity, membrane integrity, and PS (phosphatidylserine) distribution of frozen goat spermatozoa were determined in this study.
Materials and Methods
Freezing extenders
Unless otherwise stated, all chemicals were analytical reagent grade. The disaccharides trehalose, sucrose, maltose, and lactose were purchased from Sigma Chemical Co. (St. Louis, MO). The Annexin V-FITC Apoptosis Detection Kit containing propidium iodide (PI) was purchased from KeyGEN Biology (Nanjing, Jiangsu province, China).
Five different freezing extenders were used in this study. Extender NS, which contained no disaccharide, is composed of the basic extender TCG, as defined by Salmon and Ritar 27 (375 mM Tris; 124 mM citric acid; 41.0 mM glucose), and 20% (V/V) egg yolk, 0.05%(W/V) sodium dodecyl sulfate (SDS), 5% (V/V) glycerol, 100,000 IU penicillin, and 100,000 IU streptomycin. The other four extenders were composed of 375 mM disaccharide (either trehalose (T), sucrose (S), maltose (M), or lactose (L)), 20% (V/V) egg yolk, 0.05%(W/V) SDS, 5% (V/V) glycerol, 100,000 IU penicillin, and 100,000 IU streptomycin. All of the extenders were adjusted to pH 7.0 using Tris. The freezing extenders were centrifuged at 15,000 g for 1 h and the supernatants filtered through a 0.45 μm membrane filter (Millipore Corporation, Bedford, MA).
Semen collection, evaluation, and dilution
Semen samples were collected from 6 mature Yunling black bucks (3–4 years of age) using an artificial vagina at the farm of Yunnan Animal Science and Veterinary Institute. Immediately following collection, the ejaculates were placed in a 37°C water bath. The assessment of semen quality was performed immediately after collection. The ejaculates used in the study met the following criteria: volume of 0.75–2.0 ml; minimum spermatozoa concentration of 3×109 spermatozoa/ml; motility greater than 70%.
Unlike semen of other domestic animals, goat semen contains phospholipase A which can hydrolyze egg yolk lecithin into fatty acids and lysolecithin.1–3 This hydrolysis causes the spermatozoa membrane to be more fusogenic and induce the acrosome reaction. 3 In order to preclude the detrimental interaction between goat semen plasma and egg yolk, it is necessary to remove the seminal plasma by centrifugation.1,4,5 The semen was diluted with basic TCG extender and centrifuged at 1500 g for 15 min to remove the semen plasma. The washing step was repeated once. The spermatozoa pellet was suspended in the basic TCG extender to its initial semen volume.
The spermatozoa suspension was divided into five equal parts, with each diluted in one of the extenders -NS, T, S, M, or L. The final concentration of spermatozoa in the diluted semen was approximately 5×108/ml.
Freezing and thawing
The diluted semen divided into the 0.25 ml plastic straws (IMV, France) was cooled to 5°C at −0.01°C/s and then equilibrated for 2 hours. Following equilibration at 5°C, the straws containing diluted spermatozoa were put on a self-made aluminum plate and frozen in liquid nitrogen vapor for 5 min. The freezing rate was approximately −1.5°C/s. The distance between the aluminum plate and the liquid nitrogen was 4 cm. Following the above treatment, the straws were directly plunged into liquid nitrogen. The cooling/freezing rate was measured with a thermometer (Testo 925, TESTO, Germany). The thermocouple was placed in the middle of the plastic straw. The measurement of the cooling rate was made between 25°C and 5°C. The measurement of the freezing rate was made between 5°C and −50°C. After cryopreservation for 1 month, the frozen spermatozoa were thawed in a 37°C water bath for 30 sec. The thawed spermatozoa were diluted with the basic TCG extender in a ratio of 1:9 (V/V), allowed to equilibrate at 37°C for 10 min, and then evaluated for motility and path velocity (VAP) using a computer-assisted spermatozoa analysis system (CASAS) (MaiLang, Songjingtianlun Co., Nanning, Guangxi Province, China). The CASAS settings were as follows: frame required=25, fields of vision acquired=10, gray threshold=60, minimum size of spermatozoa=5, maximum size of spermatozoa=50.
Thermal resistance test
Owing to the lower motility of frozen-thawed goat spermatozoa, more time may be needed to finish oocyte–spermatozoa fusion. However, the thermal resistance test can be used to evaluate the stability of goat spermatozoa at a relatively higher temperature. 7 Following cryopreservation and thawing, the goat spermatozoa were diluted with the basic TCG extender in a ratio of 1:3 (V/V), incubated in a 37°C water bath for 0 h, 1 h, 3 h, or 5 h, and motility parameters determined using the CASAS.
Acrosome integrity
The acrosome reaction of mammalian spermatozoa is a prerequisite in order for spermatozoa to fuse with oocytes. Maintenance of the acrosome is critical for cryopreservation and the integrity can be assessed using FITC-labeled lectin from pisum sativum (FITC-PSA). 28 Briefly, aliquots (15 μl) of semen were fixed in 4% paraformaldehyde for 30 min at room temperature. Thirty μl of FITC-PSA (Sigma Chemical Co.) diluted in phosphate buffered saline (200 μg/ml) was added to the upper suspension. The mixed suspensions were incubated in a dark, moist chamber for 30 min at 37°C.The percentage of spermatozoa labeled with FITC-PSA was analyzed by flow cytometry. Spermatazoa with high fluorescence (>1000) were classified as having intact acrosomes; spermatozoa with low fluorescence (<1000) were classified as having damaged acrosomes. 28
Phosphatidylserine distribution and membrane integrity
Apoptosis is regarded as a major cause of freezing-induced cell death.29,30 Exposure of phosphatidylserine (PS) is considered a marker of apoptotic cells. In order to evaluate the effect of cryopreservation on the distribution of PS in goat spermatozoa, annexin V-FITC was used to label exposed PS. After cryopreservation and thawing, approximately 1–5×105 goat spermatozoa were suspended in 500 μl of binding buffer. Five μl of PI and annexin V-FITC were added to the upper suspension and mixed. After incubation for 15 min in the dark at room temperature, the percentage of spermatozoa labeled with annexin V-FITC and PI was analyzed using flow cytometry. Spermatozoa labeled with PI were classified as those with damaged membranes; spermatozoa labeled with annexin V-FITC without PI were classified as early apoptotic; spermatozoa labeled with both annexin V-FITC and PI were classified as late apoptotic; spermatozoa with no label were classified as normal.
Flow cytometry
The percentage of spermatozoa labeled with FITC-PSA, PI, or annexin V-FITC was analyzed using a FacStar-plus flow cytometer (FACSCalibur, Becton Dickinson and Co., Franklin Lakes, NJ). The excitation wavelength was 488 nm and the emission wavelength was 530 nm. The green fluorescence of FITC was detected by the FL1 detector, while the red fluorescence of PI was detected by the FL2 detector. Data were collected from 20,000 events for further analysis with Cell-Quest software (Becton Dickinson).
Statistical analysis
The data were collected from six bucks and analyzed using one-way ANOVA with calculation of the least significant difference (LSD). All results with a value of p<0.05 or p<0.01 were considered statistically significant. The data were expressed as mean±SD.
Results
Motility parameters of goat spermatozoa following cryopreservation and thawing
As seen in Figure 1, following cryopreservation and thawing, the motility of spermatozoa frozen with the NS extender was significantly higher (45.21±3.51%, p<0.01) than that of spermatozoa frozen with either T, S, M, or L extenders (approximately 20%). In addition, the effect of the four disaccharides (trehalose, sucrose, maltose, and lactose) on motility of frozen spermatozoa was similar (p>0.05).
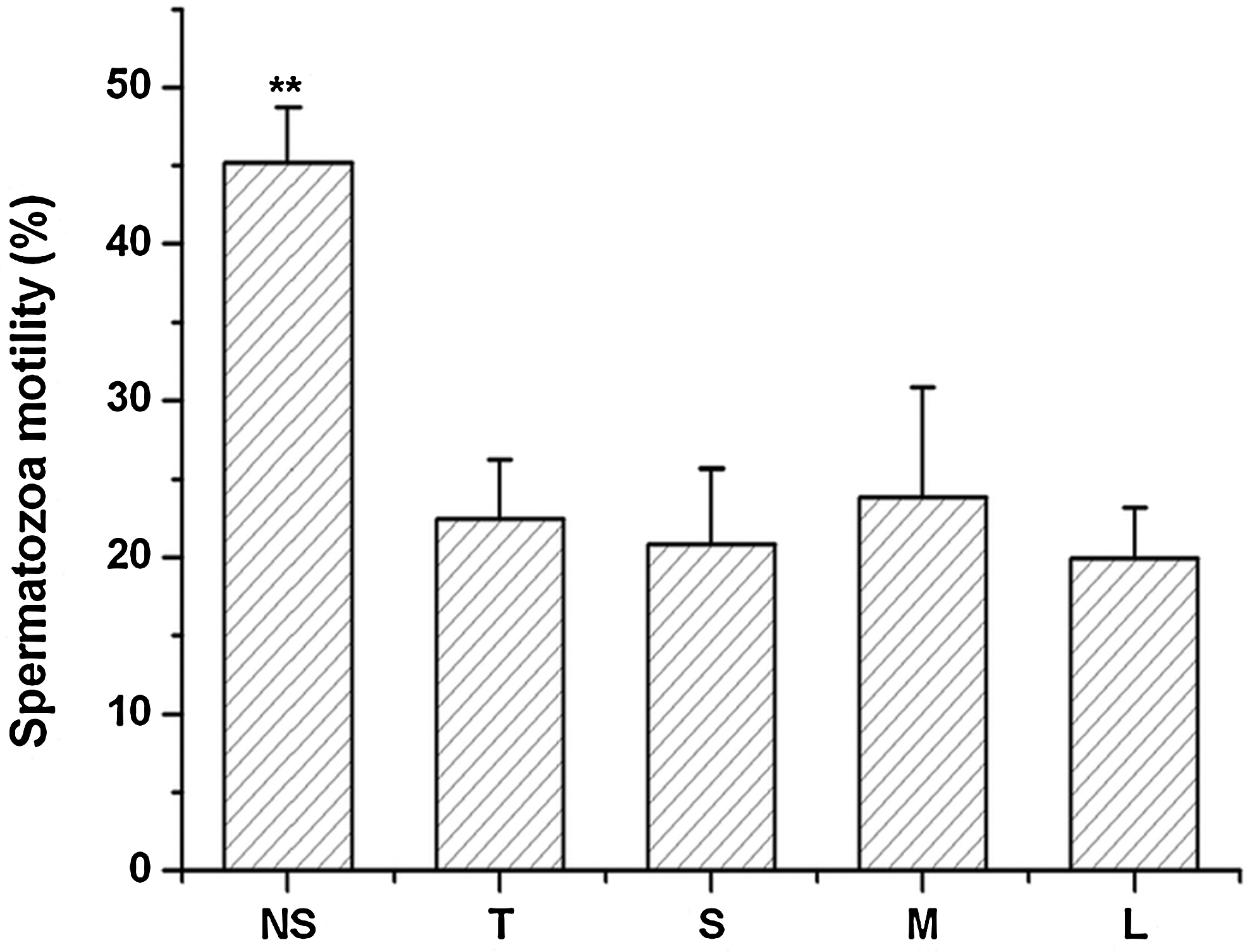
The effect of cryopreservation on motility of goat spermatozoa. NS represents spermatozoa cryopreserved in the TCG extender free of disaccharides; disaccharides used for cryopreservation: T, trehalose; S, sucrose; M, maltose; and L lactose. The data points are expressed as means±SD, n=6 in each group. **p<0.01 vs. the other groups. The data were statistically analyzed by one-way ANOVA and LSD multiple comparison.
The effect of cryopreservation on the VAP of goat spermatozoa is shown in Figure 2. The VAP of spermatozoa frozen in the NS extender was 28.66±13.57 μm/s, significantly higher than that of spermatozoa frozen with any of the disaccharide extenders (p<0.01), which was approximately 12 μm/s. Furthermore, there is no significant difference in motility of goat spermatozoa when any of the four disaccharides were used for cryopreservation (p>0.05).

Effect of cryopreservation on path velocity (VAP) of goat spermatozoa. NS represents spermatozoa cryopreserved in the TCG extender free of disaccharides; disaccharides used for cryopreservation: T, trehalose; S, sucrose; M, maltose; and L, lactose. The data points are expressed as means±SD, n=6 in each group. **p<0.01 vs. the other groups. The data were statistically analyzed by one-way ANOVA and LSD multiple comparison.
Thermal resistance of goat spermatozoa following cryopreservation and thawing
The results of thermal resistance of frozen-thawed goat spermatozoa are shown in Figures 3 and 4. Figure indicates that the incubation time at 37°C greatly influences the motility of spermatozoa following cryopreservation and thawing. As incubation time increases from 0 to 5 hours, the motility of frozen-thawed spermatozoa steadily decreases. However, the motility of spermatozoa frozen in the NS extender was significantly higher than that of spermatozoa cryopreserved in the T or S extender at the same incubation time points (p<0.05). Moreover, Figure 3 shows that the effect of trehalose was similar to that of sucrose on the motility of frozen goat spermatozoa (p>0.05). Figure 4 indicates that the VAP of frozen-thawed spermatozoa decreased steadily with prolonged incubation time. In addition, the negative effect of trehalose (T) or sucrose (S) on the VAP of frozen goat spermatozoa was significantly greater than that of spermatozoa frozen in the NS extender at each time point (p<0.05). Finally, the effect of trehalose was similar to sucrose on the VAP of frozen goat spermatozoa (p>0.05).
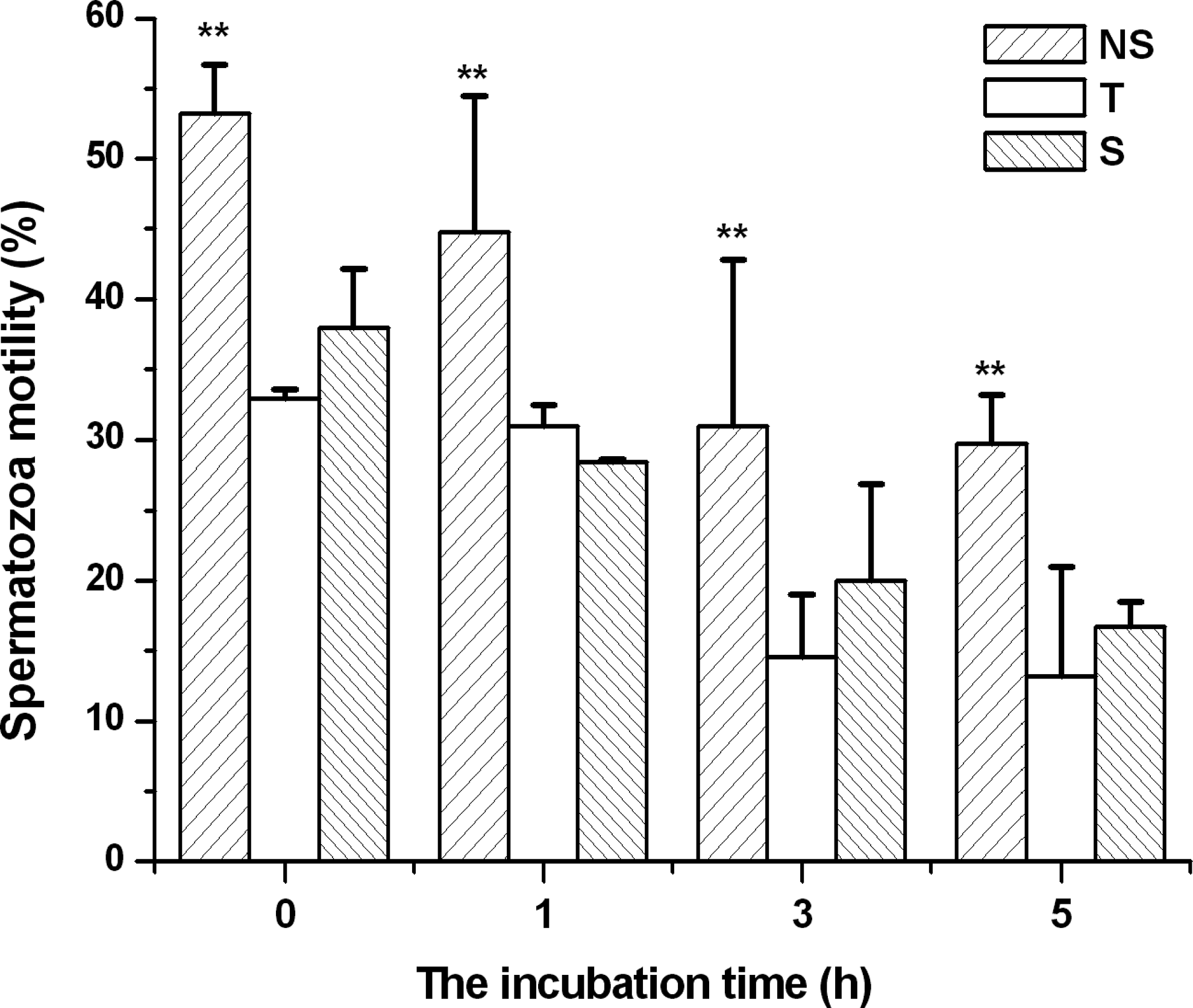
The effect of incubation time on motility of goat spermatozoa following cryopreservation and thawing. NS represents spermatozoa cryopreserved in the TCG extender free of disaccharides; T and S represent spermatozoa cryopreserved in the trehalose or sucrose extender, respectively. The data points are expressed as means±SD, n=6 in each group. **p<0.01 vs. the other groups after incubation for the same time. The data were statistically analyzed by one-way ANOVA and LSD multiple comparison.
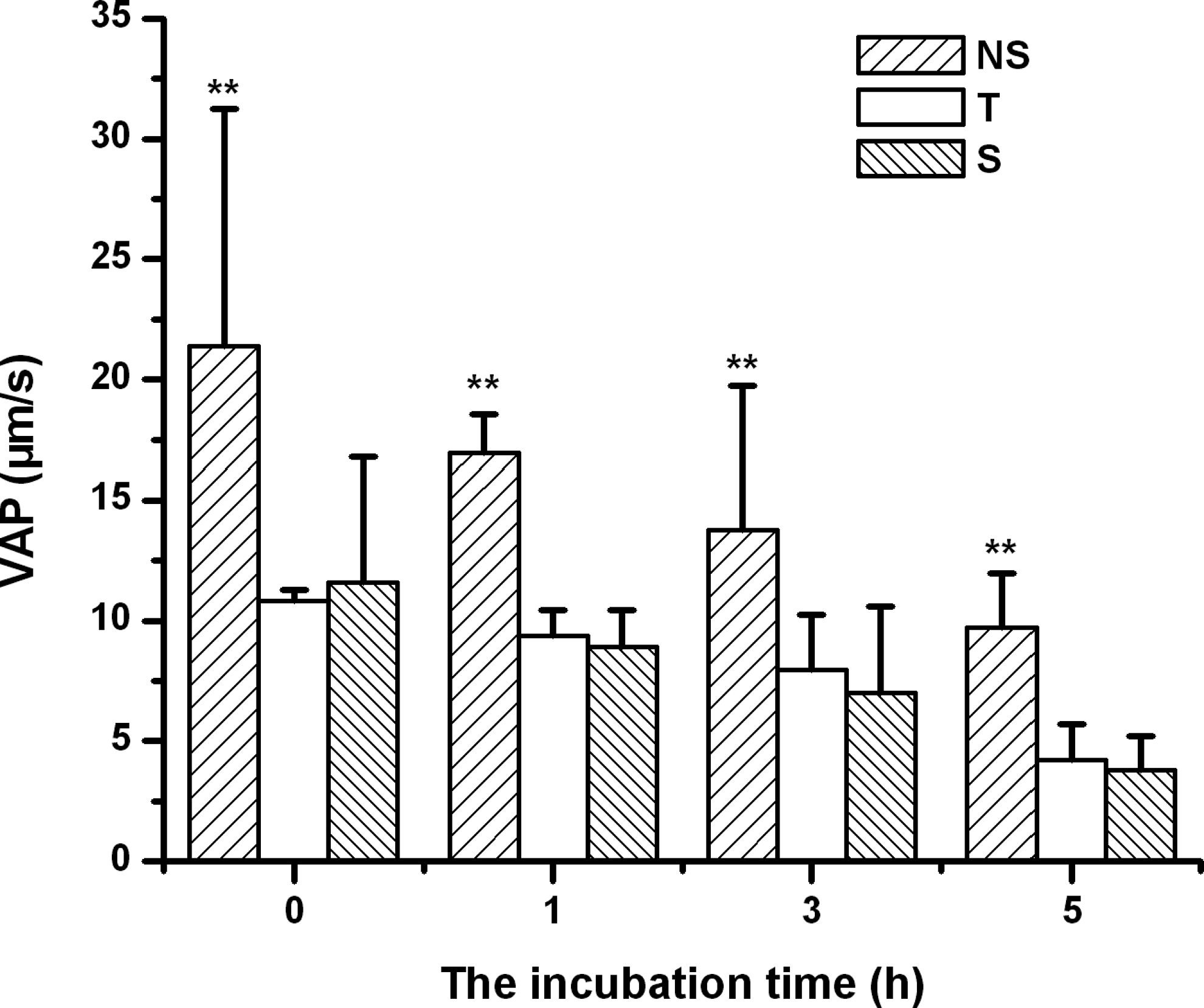
The effect of incubation time on VAP of goat spermatozoa following cryopreservation and thawing. NS represents spermatozoa cryopreserved in the TCG extender free of disaccharides; T and S represent spermatozoa cryopreserved in the trehalose or sucrose extender, respectively. The data points are expressed as means±SD, n=6 in each group. **p<0.01 vs. the other groups after incubation for the same time. The data were statistically analyzed by one-way ANOVA and LSD multiple comparison.
Acrosome integrity of goat spermatozoa following cryopreservation and thawing
Acrosome status directly affects the fertility of goat spermatozoa following cryopreservation. After labeling with FITC-PSA, the acrosome status of frozen-thawed spermatozoa using different extenders was analyzed by flow cytometry 28 (Fig. 5). In Figure 5A, the M1 field represents spermatozoa with intact acrosomes, while the M2 field represents spermatozoa with damaged acrosomes. As shown in Figure 5B, the presence of disaccharide improved acrosome status of spermatozoa following cryopreservation. In the presence of trehalose, sucrose, or maltose, the percentages of spermatozoa with intact acrosomes were 22.33±3.56%, 27.08±3.39%, and 27.18±1.47%, respectively, which was significantly higher than for spermatozoa frozen in the NS extender (16.38±2.00%, p<0.05). In addition, the percentage of spermatozoa with intact acrosomes following cryopreservation with sucrose or maltose was significantly higher than that of spermatozoa frozen with trehalose (p<0.05).
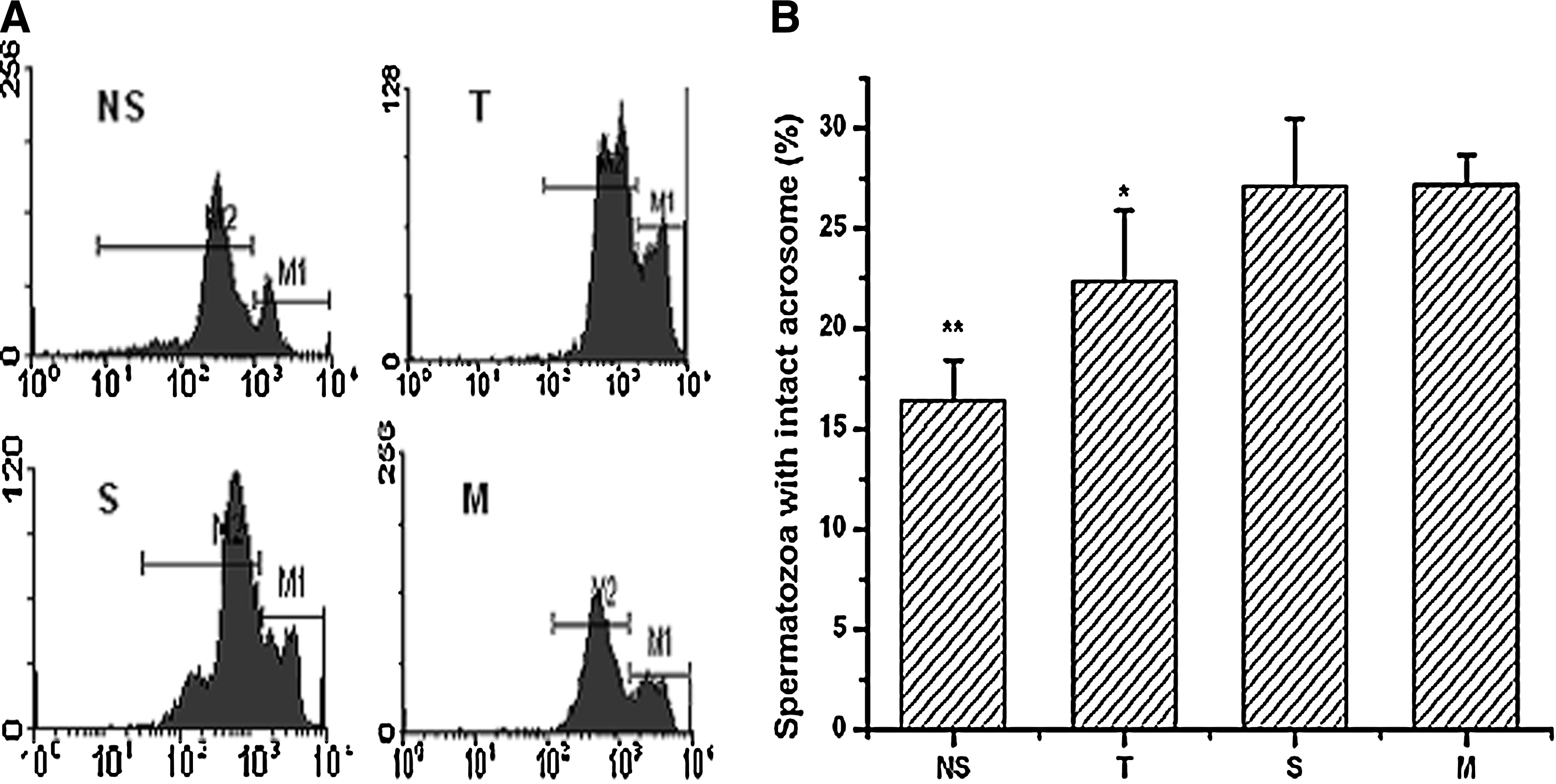
The effect of cryopreservation on acrosome status of goat spermatozoa.
Phosphatidylserine distribution and membrane integrity of goat spermatozoa following cryopreservation and thawing
Exposure of PS on the outside of the membrane is a typical feature of cell apoptosis. In this experiment, annexin V-FITC and PI were used to determine the distribution of PS of frozen spermatozoa. The results are shown in Figures 6A and 6C. As shown in Figure 6A, the dot plots of cells labeled with annexin V-FITC and PI can be divided into four fields. The upper-left field represents damaged cells labeled with only PI; the upper-right field represents late apoptotic cells labeled with Annexin V-FITC and PI; the lower-left field represents unlabeled cells; and the lower-right field represents early apoptotic cells labeled only with annexin V-FITC membranes. The detailed data are shown in Figure 6C. After cryopreservation in disaccharide extender, the percentage of cells with exposed PS was greater than 35%, and significantly higher than that of cells following cryopreservation in the NS extender (4.88±3.41%, p<0.01). In addition, the effect of trehalose was not significantly different from that of sucrose or maltose on PS distribution of frozen cells (p>0.05).

The effect of cryopreservation on membrane integrity and PS distribution of goat spermatozoa.
Cryopreservation and thawing can damage the membrane of mammalian cells. 13 In this experiment, PI was used to label spermatozoa with damaged membranes shown in Figures 6A (upper fields: damaged) and B (lower fields: intact). As seen from Figure 6B, after cryopreservation in the NS, T, S, or M extenders, more than 40% of spermatozoa were labeled with PI. Moreover, the effect of the four extenders on the membranes of frozen spermatozoa were similar (p>0.05).
Discussion
Development of an efficient and effective freezing method is very important for the creation of a goat spermatozoa bank. At present, there are two approaches to using disaccharides for cryoprotection of mammalian spermatozoa which result in differences in the osmolarity of the freezing extender, First, the disaccharide is added directly to the basic extender, which can result in increasing osmolarity of the freezing extender.12–14,16 In order to avoid the effect of osmotic differences, some investigators substitute the solutes in the basic extender with disaccharide.7,10,11 Aboagla and Terada adopted the second approach to evaluate the protective effect of trehalose on frozen goat spermatozoa. They found that trehalose significantly improved the quality of frozen goat spermatozoa.7,10,11 Moreover, compared with sucrose, trehalose enhanced the fluidity of the goat spermatozoa membrane, which might correlate with the protection by trehalose during freezing. 7 However, in their study, the protective effect of sucrose on frozen goat spermatozoa was not evaluated. 7 So whether the protective effect of trehalose is better than that of other disaccharides is unknown.
In this study, five different freezing extenders were used for cryopreservation of goat spermatozoa. In contrast to the study by Aboagla and Terada, a 0.25 ml plastic straw was used as the spermatozoa-loaded device during cryopreservation. Additionally, the prefreezing procedure was conducted in liquid nitrogen vapor, not on dry ice. Our results indicate that, compared with other disaccharides, trehalose was not particularly effective at stabilizing goat spermatozoa frozen using our developed method. In fact, in this study, the presence of disaccharide aggravated the injury to cells caused by freezing and thawing. Additionally, although the motility of goat spermatozoa frozen in the NS extender was significantly higher than that of goat spermatozoa frozen in the disaccharide extenders (Figs. 1 and 2), the presence of disaccharides efficiently protected acrosome integrity (Fig. 5). This result was similar to those reported by Aboagla and Terada. 7 In their study, as the trehalose concentration was increased, the percentage of goat spermatozoa with intact acrosomes following cryopreservation and thawing steadily increased. 7
At first, our study indicated that the presence of glucose could confer better improvement in forward motility of goat spermatozoa than disaccharide after cryopreservation, which has been shown by other investigators.8,16,31 Glucose is essential for energy utilization by spermatozoa and can support sperm motility and movement patterns.1,8,16 In this study, there was no glucose in the extenders in the presence of disaccharide. In the absence of glucose, serious damage in the motility of frozen goat spermatozoa was demonstrated (Figs. 1 and 2). In addition, the absence of glucose also influenced the PS distribution on the spermatozoa membrane. As seen in Figure 6, after cryopreservation in the absence of glucose, the percentage of spermatozoa with exposed PS was significantly higher than that following cryopreservation in the presence of glucose (p<0.01). The ATP-dependent aminophospholipid translocase can concentrate PS on the inner side of the membrane. 32 However, owing to the absence of glucose, production of ATP in goat spermatozoa may be decreased, which can result in the depression of aminophospholipid translocase activity and lead to exposure of PS. This study further showed that addition of glucose might be necessary for efficient cryopreservation of goat spermatozoa. The osmolarity of the freezing extenders used in this study was approximately 375 mOsm in the absence of glycerol, egg yolk, and antibiotic. So trehalose could not display its predominance to induce cell dehydration and inhibit intracellular ice formation. The buffering solute could also affect the viability of frozen goat spermatozoa. Salamon and Ritar investigated the interaction between Tris and sugars present in the extenders. 27 They found that addition of glucose or fructose in the Tris buffer resulted in higher post-thaw motility. On the contrary, addition of lactose or raffinose in the Tris buffer resulted in lower percentages of motile cells. 27 They concluded that goat spermatozoa prefer fructose or glucose in the presence of Tris during cryopreservation. 27 In this study, glucose in the extenders was fully substituted with alternate disaccharides. Moreover, the pH value was adjusted with Tris, so the interaction between disaccharide and Tris may decrease the quality of frozen goat spermatozoa. Finally, this study indicated that the interaction between disaccharide and cell membrane did not improve the quality of frozen spermatozoa. At present, while the water replacement hypothesis raised by Crowe et al. is often used to explain the protective effect of trehalose during freezing,23,24 it must be noted that trehalose may be effective in preserving dry biomaterials but not those in the frozen state. The reports of the Crowe group22–24 have often been misconstrued by investigators working with freezing. Our results show that there is nothing special about trehalose for freezing biomaterials compared with other disaccharides.
In conclusion, in this study, the cryoprotection afforded by the basic TCG extender, as defined by Salamon and Ritar, 27 was better than that of other extenders containing disaccharide, as defined by Aboagla and Terada. 7 Moreover, the presence of disaccharide did not improve the quality of frozen goat spermatozoa with the exception of acrosome integrity. Glucose is essential for survival of goat spermatozoa during freezing. Moreover, compared with other disaccharides including sucrose, maltose, and lactose, trehalose did not show superiority for cryopreservation of goat spermatozoa using our developed freezing method. The role of disaccharide on goat spermatozoa during cryopreservation still needs further research.
Footnotes
Acknowledgments
The authors would like to thank Mr. Jian Li from Kunming Institute of Zoology for his help with flow cytometry.
Author Disclosure Statement
The authors reported no conflicts of interest.
This study was funded by the Technological Plan of Yunnan Province, China (Grant No. 2008LA017) and the National Infrastructure of Animal Genetic Resources (NIAGR).