
Abstract
Background:
Efficient cryopreservation of human hepatocytes is essential for their use in cell therapy. This study investigated the effects of adding melatonin and/or dimethyl sulfoxide (DMSO) to pre-incubation and/or freezing solutions on the viability and function of thawed human hepatocytes.
Methods:
Isolated human hepatocytes were pre-incubated for 90 min at 4°C in Williams' Medium E (WEM), WEM containing 5 mM melatonin dissolved in DMSO, or WEM containing the equivalent amount of DMSO (1%). The hepatocytes were frozen in University of Wisconsin solution (UW) and 10% DMSO, with or without 5 mM melatonin. After thawing, viability, plating efficiency, mitochondrial dehydrogenase activity (MTT), and albumin and urea production were analyzed.
Results
: Viability and plating efficiency were not affected by melatonin or DMSO in pre-incubation media. Unexpectedly, hepatocytes pre-incubated with DMSO had significantly higher MTT (29.7% vs. control, p<0.01), albumin (82.8% vs. control, p<0.05), and urea amounts (26.2% vs. control, p=0.06) than those incubated only with WEM. Hepatocytes pre-incubated in media containing melatonin had amounts between those of cells incubated with DMSO or only with WEM (p<0.05 for MTT and p>0.05 for albumin and urea values). Also, the addition of melatonin to the freezing media did not significantly improve any of the studied parameters (p>0.05).
Discussion:
Adding 1% DMSO to pre-incubation media prior to the cryopreservation of human hepatocytes preserves hepatocyte function after thawing. These findings could be considered in current hepatocyte cryopreservation protocols.
Introduction
Currently, the only method for the long-term preservation of hepatocytes is cryopreservation. However, the procedure seriously compromises cell viability, attachment efficiency, and functionality.3,4 Efforts have been made to optimize the cryopreservation protocol by reducing the harmful effects of freezing/thawing.5–8
Prior to cryopreservation, hepatocyte isolation also results in cell impairment, to a large extent due to oxidative stress. 9 Thus, the condition of hepatocytes is already delicate after isolation. Pre-incubation of isolated hepatocytes prior to cryopreservation is a method to partially recover cell function and, as a result, obtain better-quality cryopreserved hepatocytes.5,6,10 Adding compounds to the pre-incubation media to boost cellular ATP content,7,8 or antioxidants to reduce/repair any damage resulting from the isolation procedure, may enhance the beneficial effects of pre-incubation. 7
During the cryopreservation/thawing process, cell death occurs due to cell rupture caused by intracellular ice formation and cell dehydration, necrosis, or apoptosis.11,12 Different compounds have been shown to reduce the harmful effects of cryopreservation, mainly osmotic shock, when added to the freezing solution.13,14 Substances that protect the mitochondrial membrane could also decrease the harmful effects of cryopreservation on primary human hepatocytes.
Melatonin, a pineal gland hormone with important direct and indirect antioxidant effects, can easily cross cell membranes. 15 Melatonin prevents the breakdown of the mitochondrial membrane potential and decreases electron leakage, thereby reducing the formation of superoxide anions. 16 The hormone is also a powerful scavenger and inhibitor of the free radicals that arise in cells after physical damage, 17 and it has been shown to have beneficial effects on the liver under oxidative stress situations.18–23
Melatonin may be able to partially recover the condition of isolated hepatocytes in order to better survive cryopreservation, or even to reduce the physical and mitochondrial damage caused by the cryopreservation and thawing process.
Dimethyl sulfoxide (DMSO) is one of the most widely used solvents as a vehicle for water-insoluble substances. Among other properties, it has been shown to penetrate cell membranes easily and to remove free radical products, working as an effective antioxidant. 24 In fact, it has been shown to reduce reactive oxygen species (ROS) formation and to enhance hepatocyte differentiation and function in rat hepatocytes in culture at 37°C and 5% CO2.25,26
The aim of this study was to evaluate the effects of incubating human hepatocytes with melatonin or DMSO prior to cryopreservation and/or the addition of melatonin to the freezing solution on hepatocyte viability and function after thawing.
Material and Methods
Isolation of human hepatocytes
Adult normal human liver samples were obtained from seven patients (2 males/5 females; mean age=63.6±3.03 years), undergoing partial hepatectomy because of tumor disease. Four of the seven patients had previously received chemotherapy treatment. All samples were obtained and processed in compliance with the Clinical Research Ethics Committee of Aragon. Liver samples were maintained in cold University of Wisconsin Solution (UW) (ViaSpan, Bristol-Myers Squibb Pharma Ltd., Madrid, Spain) until cell isolation. Hepatocytes were isolated using a two-step collagenase perfusion technique.6,27 After each isolation, cell viability was assessed by the Trypan-blue exclusion method. Both pre-incubation and freezing media effects were studied for each isolated liver sample. The experimental design of the study is shown in Figure 1.

Schematic flowchart of the experimental design. A color version of this figure is available in the online article at www.liebertonline.com/bio
Pre-incubation procedure
Melatonin is not water-soluble and is usually dissolved in organic solvents, such as ethanol or DMSO. Because of the toxic effects of ethanol on hepatocytes, 28 DMSO was chosen as the solvent. Melatonin was dissolved in DMSO at a stock concentration of 500 mM. Because of the properties of DMSO, two different control pre-incubation treatments were included: without any substance added (control) or with DMSO).
Because of the limited amount of cells obtained from each isolation process and the number of samples to be run at the same time, only a single dose of melatonin was studied. The melatonin dose was chosen according to previous data showing a beneficial effect on hepatic cells under oxidative stress conditions in vitro 29 and in vivo. 20
Before cryopreservation, freshly isolated hepatocytes were suspended in Williams' Medium E (WEM) (Lonza, Verviers, Belgium) including 5 mM melatonin (Fagron Ibérica, Spain) or 1% DMSO (Sigma-Aldrich, St. Louis, MO), which is equivalent to the DMSO added with melatonin, at a density of 1.5×106 viable hepatocytes per milliliter pre-incubation media at 4°C for 90 min. As conclusive data about optimal pre-incubation times do not exist in the literature,5–7,10 a 90 min pre-incubation was chosen as a practical and reasonable amount of time to see the potential effects on hepatocytes of the addition of melatonin or DMSO before freezing. As a control, the same number and density of hepatocytes were incubated in WEM alone (Fig. 1).
Freezing protocol
After pre-incubation, cells were centrifuged at 50 g at 4°C for 5 min and the pellets suspended in freezing media. For half of the cells, the freezing media was ice-cold UW solution containing 10% DMSO, added to the cell pellet in a drop-wise manner. For the remaining cells, the same freezing medium was used but containing 5 mM melatonin (Fig. 1). The final DMSO concentration was maintained at 10%. Cells in cryovials were immediately placed in Nalgene Cryo 1°C Cell Cooler (Nalge Nunc International, Rochester, NY) and into a −80°C freezer, providing a temperature decrease rate of −1°C/min. After 24 h, the cryovials were moved to liquid nitrogen until thawing. For each treatment, two cryovials were frozen/thawed with 1 ml of freezing solution per cryovial at a cell density of 3×106 cells per milliliter of freezing solution.
Thawing protocol
Cell vials were rapidly thawed by gentle agitation in a 37°C water bath for 1 to 2 min. 29 The cryoprotectant was immediately removed by successive dilution with thawing media consisting of ice-cold WEM containing 10% fetal calf serum (Lonza, Verviers, Belgium). For every 1 ml of cell suspension, 0.5, 1, 2, 3, and 6 ml of thawing media was added drop-wise, with 5 min on ice between each addition. Thereafter, cells were pelleted by centrifugation at 50 g for 5 min at 4°C, and the pellet suspended in a known volume of cell seeding culture media consisting of Ham's F-12/WEM (1:1,v/v) supplemented with 2% newborn calf serum (NCS), 0.1% bovine serum albumin fraction V, 10 nM insulin, 25 μg/ml transferrin, 26 mM sodium bicarbonate, 66.8 μM ethanolamine, 7.2 μM linoleic acid, 7 mM glucose, 0.62 mM ascorbic acid, 2 mM L-glutamine, 0.64 mMNω-Nitro-L-arginine methyl ester, 100 U/ml penicillin, 100 μg/ml streptomycin, and 0.25 μg/ml amphotericin B (all purchased from Sigma-Aldrich, except NCS and the antibiotic/antimycotic solution, which were purchased from Lonza).
Cell viability and function assays
The Trypan blue exclusion method was used to evaluate viability and determine the cell count. For functional assays, cells were seeded at a density of 30,000 viable cells per well in 96-well, flat-bottomed, collagen-coated plates (Biocoat™, BD Bioscience, Belgium).
Mitochondrial dehydrogenase activity (MTT) of cultured cells was assessed using a method based on the reduction of a tetrazolium salt, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, into a purple formazan product. After 16 h in culture, the media was removed and the cells incubated for 4 h with the reagent (at a final concentration of 500 μg/ml; Sigma-Aldrich) in serum-free culture media (without NCS and supplemented with 10 nM dexamethasone). The supernatant was removed, and the formazan crystals in each well were dissolved in 200 ml DMSO. The absorbance was read at 570–630 nm using a microplate reader.
Plating efficiency (PE) was examined by seeding the same number of thawed hepatocytes and determining the protein content of attached cells 16 h after plating compared with the initial number of cells. Protein content was measured using the Bicinchoninic Acid Protein Determination kit (Sigma-Aldrich).
Albumin production was determined in 16-h supernatants using a Human Albumin ELISA Quantification Kit (Bethyl Laboratories, Inc., Montgomery, TX). An albumin standard curve (0–1500 ng/ml) was used to calculate the albumin production expressed as micrograms albumin produced per milligram of adhered cell protein. The urea concentration was measured in the same supernatants using a colorimetric urea kit (QuantiChrom™ Urea Assay Kit, DIUR-500, Bioassay Systems, Hayward, CA) as previously described. 31
Calculations and statistical analysis
The MTT activity for each isolation was obtained as an average of the values in eight wells. For cell adhesion, albumin, and urea determinations, supernatants were pooled from eight wells; adhered protein content was determined in eight separate wells. Protein, albumin, and urea were analyzed in duplicate for each sample. Data were analyzed in a randomized block design using two-way analysis of variance in which the isolation effect was considered as the random blocking effect, and pre-incubation (control, with DMSO, or with melatonin) and cryopreservation (control or with melatonin) effects were considered fixed factors. When resulting pre-incubation or cryopreservation effects were significant (p<0.05), means were compared post hoc using the test of least significant difference (LSD). Calculations were performed using the SAS software package (SAS Institute Inc., 2000).
Results
Viability and functionality values for each cryopreserved/thawed hepatocyte group are presented in Table 1. No significant (p>0.05) interactions were found between pre-incubation and cryopreservation effects in any of the studied parameters; pre-incubation effects were not affected by cryopreservation effects or vice versa. Accordingly, the results are presented separately for each main effect, for the effect of pre-incubation media (Fig. 2) and for the effect of the freezing solution used (Fig. 3).
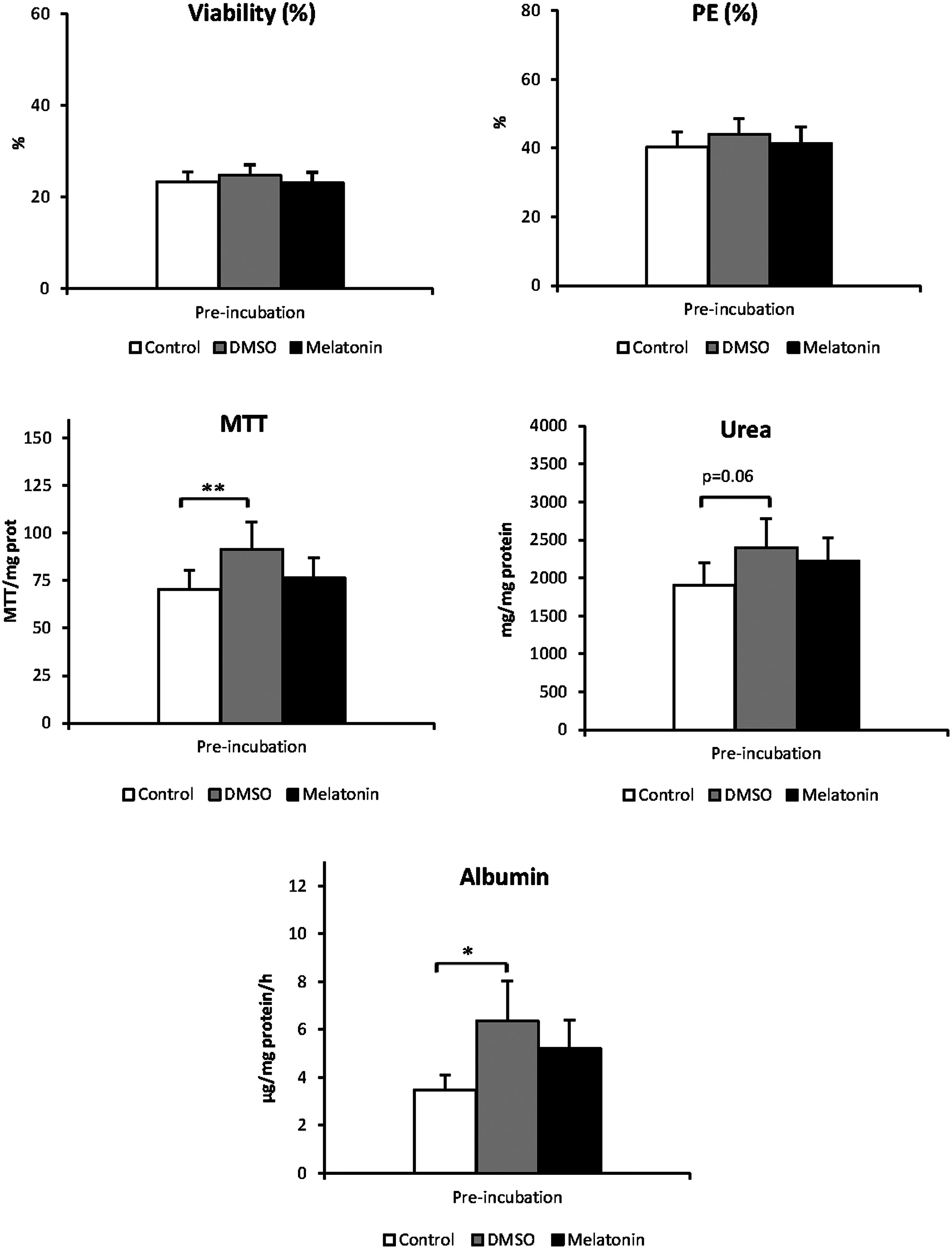
Effect of DMSO and melatonin addition to pre-incubation media prior to cryopreservation on thawed human hepatocyte viability and function. Results are presented for each pre-incubation treatment (Control, white, pre-incubated with WEM; DMSO, gray, pre-incubated with WEM+1% DMSO; Melatonin, black, pre-incubated with WEM+5 mM Melatonin in DMSO (1%)) as means and standard error of samples of 7 different isolations and 2 types of cryopreservation (n=14). MTT, mitochondrial dehydrogenase activity; PE, plating efficiency; Values are compared to those of standard hepatocytes without DMSO or melatonin in the pre-incubation media. Significance: *p<0.05; **p<0.01.

Effect of melatonin addition to freezing medium on thawed human hepatocyte viability and function. Results are presented for control, white (Freezing medium composed of UW solution and 10% DMSO) and melatonin, black (Freezing medium composed of UW solution+10% DMSO+5 mM melatonin) treatments as means and standard error of samples of 7 different isolations and 3 types of pre-incubation media (n=21). No significant differences in any studied variable were found between hepatocytes cryopreserved with or without melatonin.
After isolation, the primary human hepatocytes had an average viability of 72±3.4%. We did not assess cell viability after pre-incubation before freezing the cells because pre-incubation at 4°C is a method that has been shown to improve human hepatocyte cryopreservation, 7 and it is currently used in cryopreservation protocols. 30 Cryopreservation considerably reduced viability after thawing. The viability of thawed hepatocytes averaged 23.7±1.28%.
The addition of DMSO or melatonin to the pre-incubation media did not alter hepatocyte viability after cryopreservation. Cell adhesion to the collagen surface of culture plates did not differ among hepatocytes pre-incubated with DMSO, melatonin, or WEM alone (average 41.9±2.61%). Hepatocytes pre-incubated with 1% DMSO or 5 mM melatonin exhibited higher MTT activity per milligram of adhered cell protein than control hepatocytes (p<0.01 vs. DMSO, p>0.05 vs. melatonin; Fig. 2). The same trend was observed in the other parameters studied (Fig. 2). Hepatocytes pre-incubated with 1% DMSO had higher albumin production (p<0.05) and secreted more urea (p=0.060) compared to control hepatocytes. Similar albumin and urea concentrations were measured in supernatants from melatonin pre-incubation as those measured for 1% DMSO (p>0.05). Although, hepatocytes pre-incubated with melatonin had 49.7% and 17.1% higher values for albumin production and urea secretion, respectively, than hepatocytes pre-incubated in WEM alone, differences between both treatments were not significant (p>0.05).
When hepatocytes were frozen in media containing 5 mM melatonin, none of the studied parameters were significantly improved (p>0.05, Fig. 3) compared to hepatocytes frozen with the standard solution.
Discussion
Different cryopreservation protocols have been described for human hepatocytes,30,32 but cell viability and function are often compromised during the process,5,8 making hepatocyte cryopreservation a considerable constraint for the use of human hepatocytes in different applications.
In this study, contrary to expectation, the pre-incubation of hepatocytes with melatonin did not improve viability, adhesion to the collagen surface, metabolic activity (MTT/mg protein), or the specific functions measured. However, DMSO addition to the pre-incubation media improved all functional parameters (MTT, albumin production, and urea secretion) compared to hepatocytes pre-incubated with media alone. Although the melatonin and DMSO treatments did not differ significantly, melatonin treatment generally provided values that fell between the DMSO and control treatments. Taking into account the fact that the melatonin treatment contained the same DMSO concentration as the DMSO treatment, the observed results could suggest an inhibitory influence of melatonin on the DMSO effect.
DMSO (1%–2%) in culture media has been shown to support hepatocyte differentiation25,26 and enhance liver-specific functions, 34 and it is known to penetrate cell membranes easily and remove free radical products, working as an effective antioxidant.24,25 Although the effect of DMSO inclusion in human hepatocyte culture media has been previously reported,25,26,33 this is the first time the effect of DMSO used as a preconditioning agent to diminish the damage of human hepatocytes caused during the freeze/thaw process has been studied.
The effects of DMSO seen here could be related to the antioxidant properties of DMSO, 25 which could have enhanced the condition of the hepatocytes to endure later cryopreservation. It has also been pointed out by Stéphenne et al. 12 that a pre-adaptation period of hepatocytes to the lower doses of DMSO used during pre-incubation than those used in the cryopreservation media could have reduced the osmotic stress and intracellular ice-crystal formation during freezing and thawing. However, in accordance with this hypothesis, viability should have been higher than in control hepatocytes; we found no significant differences between the pre-incubation groups in regards to viability. The improvement of hepatocyte activity by pre-incubation with DMSO, despite cell death due to physical damage during cryopreservation and thawing, could also be related to the observed differentiation properties of DMSO on hepatocytes by boosting the expression of hepatocyte phenotype. 33
In regard to melatonin, it produced an unexpected effect in the pre-incubation media. Data in the literature point to a potentially beneficial effect of melatonin on cells in vitro and in vivo, acting both as a free radical scavenger and by stimulating the production/activity of intracellular antioxidant enzymes 15 in a way that reduces oxidative stress within cells. In the liver, the antioxidant properties of melatonin prevent both acute liver injury induced by ischemia–reperfusion, 28 irradiation, 19 cholestasis, 20 or toxins, 21 and liver degeneration, such as cirrhosis induced by thioacetamide 22 or age-induced changes. 23
Despite the well-documented beneficial effects of melatonin, other considerations could explain our current results. First, experiments in which melatonin has been found to have beneficial effects on hepatocytes under oxidative stress conditions involved the administration of melatonin for days in in vitro studies 29 or weeks in in vivo studies.20,23 Thus, melatonin activity could require some time before being effective. Reviewing the literature, there are no data on the effect of melatonin on hepatocytes when it is added for a short period of time. Radogna et al. 34 found ROS production induced by melatonin in normal and tumor leukocytes rapid and transient from the first minute, peaking at 2–3 h, and disappearing 6 h after melatonin addition at concentrations higher than 10 μM. Blackman et al. 35 investigated the effect of melatonin on hepatocyte gap junctions, only observing changes in the intercellular communication of cultured mouse hepatocytes 10 hours after melatonin treatment, reaching a maximum at 12 h. In studies in which melatonin has shown positive effects on cultured hepatocytes 29 or liver function,20,23 the hormone was administered for days (in vitro studies), or even weeks (in vivo studies). Time could explain the positive results seen with melatonin at supraphysiological levels. Therefore, we can presume that the pre-incubation time with melatonin was not enough to see the expected effects on hepatocytes,
Second, different concentrations of melatonin were used in the different studies, from physiological (nanomolar concentrations) to supraphysiological ranges, so the effect of melatonin on cells could be influenced by its concentration. Radogna et al. 34 found a pro-oxidant activity of melatonin when used in the micromolar range (10 μM–1 mM) but not at lower concentrations (10 nM–1 μM). Blackman et al. 35 found little or no effect of melatonin on cultured hepatocytes at supraphysiological concentrations. 35 Given this information, we suspect that both melatonin dose and its time in contact with cells played an important role in the results we obtained. However, only one melatonin dose was chosen for the present study because the number of freshly isolated human hepatocytes was not enough to test two or more melatonin doses at the same time in each isolation. The 5 mM dose was an approximate mean value derived from doses found to be effective in the liver in other oxidative stress-related studies.19,20,22,29 Regarding the time of melatonin contact with the cells, we had to consider that freshly isolated human hepatocytes should be cryopreserved as soon as possible because function deteriorates over hours; thus, the pre-treatment of hepatocytes prior to cryopreservation should not be longer than 24 h. 10 Therefore, the pre-incubation time was that recommended and used by other authors.7,30,32
On the other hand, melatonin addition to the cryopreservation media did not modify any studied parameter in the thawed hepatocytes. This observation can be explained by the reasons mentioned above. Other studies on the addition of antioxidants to the hepatocyte freezing solution found very little or no effect, despite reporting positive results when adding them to the culture media. 36 Nevertheless, the addition to the cryopreservation media of membrane stabilizers such as trehalose, 8 or osmotic modulators such as wheat protein extracts, 14 have been shown to improve the quality of hepatocytes post-thawing. The damage of cryopreservation/thawing on hepatocytes may have a more physical origin, so that antioxidants are not effective enough at protecting human hepatocytes during the process.
One of the limitations of our study could be that cryopreservation drastically reduced viability after thawing (average 23.7±1.28%), Nevertheless, the viability of thawed hepatocytes was comparable to that obtained by other authors, considering sample origin (all of them came from patients ranging in age from 52 to 75 years, most of whom had received previous chemotherapy treatment), which significantly affects sample quality and consequently the condition of the cells to be cryopreserved. We did not perform percoll purification after thawing15,31 and we did not use a controlled rate freezer which provides optimal temperature conditions during freezing, resulting in better post-thawing viabilities. 30 While we were aware of the importance of using a controlled rate freezer during the study, as we did not have one (similar to many other centers), we performed the cryopreservation with a Nalgene Cryo 1°C Cell Cooler that “guarantees” a temperature decrease inside the cryovial chamber of 1°C per minute in a −80°C freezer. This was the device available to us that most closely resembled a controlled rate freezer and it has been used in other studies cryopreserving human hepatocytes. 6
This study constituted an experimental approach to determine how DMSO or melatonin addition to pre-incubating and freezing media could affect cryopreservation of human hepatocytes. However, it should be pointed out that, if clinical-grade hepatocytes are required, they should be prepared under strict sterile conditions (in an accredited good manufacturing practice unit), FBS should be changed (because it is of animal origin) to Human Serum Albumin, and WEM could be substituted by UW during pre-incubation or thawing. In any case, DMSO or melatonin (at the concentration tested here) could be used.
Briefly, pre-incubating human hepatocytes with 1% DMSO at 4°C prior to their cryopreservation improves the quality of hepatocyte function and could be introduced in current hepatocyte cryopreservation protocols. Pre-incubation with 5 mM melatonin does not significantly improve hepatocyte activity, nor does the inclusion of 5 mM melatonin in cryopreservation media, although more studies using different melatonin concentrations at physiological levels and with longer incubation times should be carried out to plainly elucidate the effect of melatonin on the human hepatocyte cryopreservation process.
Footnotes
Acknowledgments
The authors are very grateful to Dr A. Lanas for allowing us to use his laboratory facilities. Dr. Solanas's work was supported by the Aragon Regenerative Medicine Program (IIS Aragón) and the CIBER Enfermedades Hepáticas y Digestivas (CIBERehd).
Author Disclosure Statement
No competing financial interests exist.