
Abstract
Effective stabilization of nucleated cells for dry storage would be a transformative development in the field of cell-based biosensors and biotechnologic devices, as well as regenerative medicine and other areas in which stem cells have clinical utility. Ultimately, the tremendous promise of cell-based products will only be fully realized when stable long-term storage becomes available without the use of liquid nitrogen and bulky, energetically expensive freezers. Significant progress has been made over the last 10 years toward this goal, but obstacles still remain. Loading cells with the protective disaccharide trehalose has been achieved by several different techniques and has been shown to increase cell survival at low water contents. Likewise, the protective effect of heat shock proteins and other compounds have also been explored alone and in combination with trehalose. In some cases, the benefit of these molecules is seen not initially upon rehydration, but over time during cellular recovery. Other considerations, such as inhibiting apoptosis and utilizing isotonic buffer conditions have also provided stepwise increases in cell viability and function following drying and rehydration. In all these cases, however, a low level of residual water is required to achieve viability after rehydration. The most significant remaining challenge is to protect nucleated cells such that this residual water can be safely removed, thus allowing vitrification of intra- and extracellular trehalose and stable dry state storage at room temperature.
Introduction
A promising alternative to storing cells frozen is the possibility that removing cellular water will allow the cells to enter a state of biostasis, or suspended animation. Many plants and animals are capable of accomplishing this task in the natural world, and such anhydrobiotes provide clues as to the types of protection needed to accomplish a similar feat with sensitive cellular samples in the laboratory. 7 Ideally, such samples could be stabilized in the dehydrated state for storage at room temperature, thus obviating the need for freezers or toxic cryoprotectants.
Much progress has been made over the last 15 years with regard to the stabilization of nucleated cells for dry state storage. There remain, however, significant challenges before the ultimate goal of room temperature storage can be realized. In this way, the effort to dehydrate nucleated cells lags behind the advancement achieved in the dehydration of other types of cells and cell fragments. Bacterial cells, for instance, can be freeze-dried with excellent survival following rehydration. 8 Even human blood platelets can be protected for lyophilization, 9 and can survive for 2 years at room temperature in the desiccated state. 10 One important reason that nucleated cells pose additional difficulties is their high level of intracellular complexity. Besides the nucleus, with its obvious importance to post-rehydration cell health and division, the other organelles, each with a different membrane composition, may have significantly different requirements for protection against the severe stresses of dehydration. Thus, the drying of nucleated cells is unquestionably a complicated but important undertaking. This review will highlight both the recent progress in the field, as well as the lingering challenges.
Loading Nucleated Cells with Trehalose
Most anhydrobiotic organisms, naturally able to survive desiccation, accumulate high concentrations of disaccharides (most commonly trehalose or sucrose) in their cells and tissues during drying.7,11–13 And while trehalose does not solve every problem associated with the drying of biological samples, it does have some important properties that enable it to improve significantly post-rehydration survival of nucleated cells. As these protective effects of trehalose have been ably reviewed elsewhere, 14 they are listed here only briefly: (1) trehalose hydrogen bonds with the headgroups of phospholipid bilayers under dehydrated conditions, thus depressing the gel to liquid crystalline phase transition (Tm), and preventing damaging phase changes during rehydration;15–18 (2) trehalose vitrifies at low water contents and is characterized by an extremely high glass transition temperature (Tg), which allows it to remain in the glass even under suboptimal conditions (elevated temperature or humidity);19–24 (3) glassy trehalose is capable of transforming to the crystalline dihydrate in the presence of water vapor, which allows it to “sequester” water and avoid plasticization, thus maintaining a high Tg; 21 (4) trehalose has a highly stable glycosidic bond, which prevents browning reactions to which other sugars are susceptible.25,26 For these reasons, trehalose serves as an excellent lyoprotectant, both in naturally occurring anydrobiotes and in the effort to protect sensitive biomolecules in the laboratory.
Because trehalose is impermeant, and since most tissues of interest lack a trehalose transporter, introducing trehalose to the intracellular milieu is a common first step in any protocol for drying nucleated cells. Several different procedures have been investigated toward this end. One protocol explored early in the effort to load cells with trehalose was to expose the cells to an electric field in the presence of a trehalose solution.27,28 This electroporation (or electropermeabilization) technique was effective at allowing trehalose to cross the plasma membrane, as the intracellular concentration was estimated at 100 mM. 27 However, the treatment also caused some collateral damage to the cells, resulting in cell shrinkage to 60% of the starting volume. 27 More recently, high intensity femtosecond laser pulses have been used to similarly permeabilize membranes in a transient way. 29 Direct microinjection can also be used to load trehalose intracellularly in large cells (∼100 μm), such as oocytes,30–32 but this technique is not efficient for smaller cells.
An alternate method for introducing trehalose inside the cell is to utilize channels in the plasma membrane. At least two types of channels have been investigated, the ATP-stimulated P2-purinergic pore,33,34 and α-hemolysin.35–37 The former is an endogenous membrane protein, present on the surface of immune cells, hematopoetic cells, and epithelia, and is controlled through the presence of ATP.38,39 The latter is a genetically engineered bacterial protein added exogenously to the nucleated cells. In both cases, trehalose was successfully loaded into the cytosol,34,36 and the α-hemolysin pore had the added benefit that it could be closed in the presence of zinc, thus preventing the loss of any important intracellular components. 37
A third general approach has been to genetically engineer cells to either take up or make their own trehalose. For instance, when the genes for trehalose biosynthesis from E. Coli (otsA and otsB) were expressed in human fibroblasts using an adenovirus vector, the cells produced increasing amounts of trehalose with increasing multiplicity of infection (MOI). 40 This trehalose production was nontoxic to the cells until a high MOI was reached. 40 In addition, the trehalose transporter TRET1 from the anhydrobiotic insect P. vanderplanki, when expressed in Xenopus oocytes, was able to function as a high capacity, trehalose-specific transporter loading the cells with disaccharide. 41
Thermally responsive Pluronic-based nanocapsules have emerged recently as a physical means for introducing trehalose intracellularly. 42 The disaccharide is encapsulated within the nanocapsule through a three-step process of incubating in trehalose solution, followed by lyophilization and elevated temperature. After cellular uptake, the trehalose can be released from the nanocapsules by temperature cycling from 37°C to 22°C, then back to 37°C, resulting in intracellular concentrations as high as 300 mM. 42
All of the techniques mentioned above, however, require direct manipulation of the cells, exposing them to nonphysiological conditions, and/or requiring insertion of non-native proteins or carriers. A method that does not require any of these manipulations is based on fluid phase endocytosis, a constitutive process. Using a technique first pioneered in platelets, 9 human mesenchymal stem cells (MSCs) were loaded with trehalose by including the disaccharide in the extracellular milieu. 43 Low temperature inhibits the uptake, likely due to a change in phase state (Fig. 1A). At 37°C, however, trehalose loading correlates with the length of incubation and shows a nonsaturating dependence on extracellular concentration (Fig. 1B), characteristic of fluid phase endocytosis (FPE). 43 Experiments with Lucifer yellow CH (LYCH), a fluorescent dye commonly used for the study of FPE, indicates that solutes taken up by this mechanism are initially sequestered within endosomes, but are also found in the cytosol over time. Using this method, intracellular trehalose concentrations in the range of 30 mM can be achieved by a 24-h incubation in 100 mM trehalose at 37°C with no significant loss in viability (Fig. 1B). 43 This general process can likely be applied to most cell types, as it has been shown proficient in several different lines besides platelets and MSCs, including J774, 293H, HeLa, and murine B cells.44–47
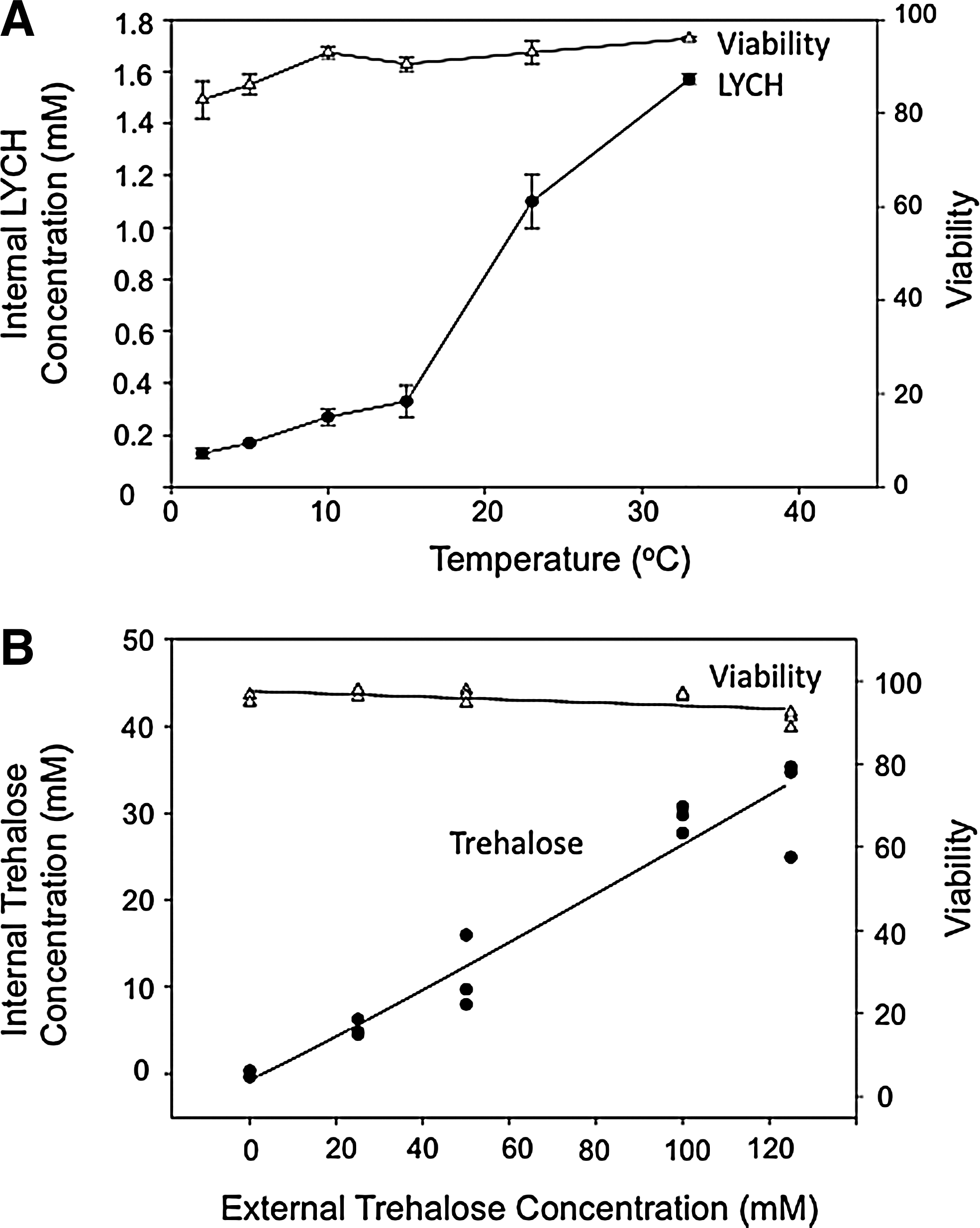
A common theme among investigations that tested for post drying/rehydration viability was the finding that trehalose loading enabled greater survival compared to cells that were not loaded with the sugar (Fig. 2).34,35,40,44–48 Despite the efficient introduction of protective solutes by various methods, however, efforts to achieve viable nucleated cells following drying and rehydration continue to be hampered by low viabilities at low water contents. In fact, regardless of loading technique, there appears to be a lower limit, in the range of 0.2–0.5 g H2O/g dry weight (DW), below which nucleated cells do not usually survive.34,35,44–48 This commonality represents the remaining significant challenge to the dry storage of nucleated cells. An easily accessible method that removes this residual water and enables trehalose vitrification, would increase by orders of magnitude the stability and storage lifetime of dry cell-based products.
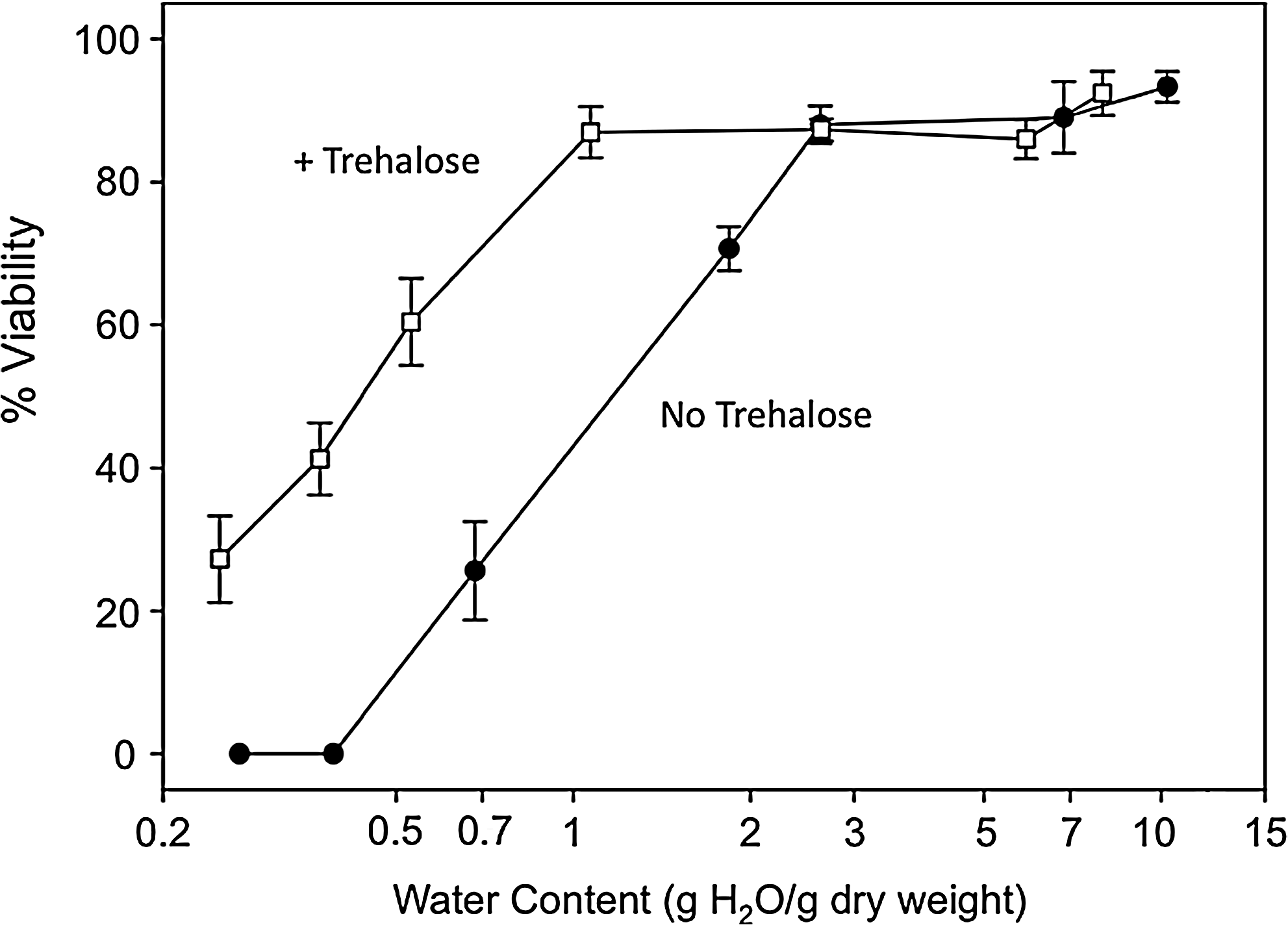
Cell viability as a function of residual water content in vacuum-dried samples, as measured by propidium iodide exclusion. Trehalose-loaded MSCs dried in buffer containing trehalose were compared with unloaded cells dried in buffer lacking trehalose. When dehydrated below 2 g H2O/g dry weight, a dramatic difference in viability emerged between the two types of samples. Data represent means and standard deviations (n=3). (Adapted from Ref. 48 with permission.)
Beyond Trehalose: Heat Shock Proteins
Although trehalose has greatly improved post-rehydration survival in many cell types, it is clearly not sufficient to defend nucleated cells from all types of damage that occur during drying and rehydration. In fact, anyhdrobiotic organisms use multiple protective strategies in order to survive the extreme stresses of desiccation. 49 For instance, encysted embryos of the brine shrimp Artemia franciscana, which can withstand removal of virtually all cellular water, contain large quantities of the small heat shock protein p26.50–52 This protein acts in concert with trehalose to inhibit oxidative damage in primate COS-1 cells, 53 and performs molecular chaperone functions in vitro and probably in vivo.52,54 Other examples exist as well of heat shock proteins (HSPs) being linked to stress resistance. HSP104 in the yeast Sacchromyces cerevisiae, for example, is linked to thermotoloerance and protection of proper protein folding.55,56
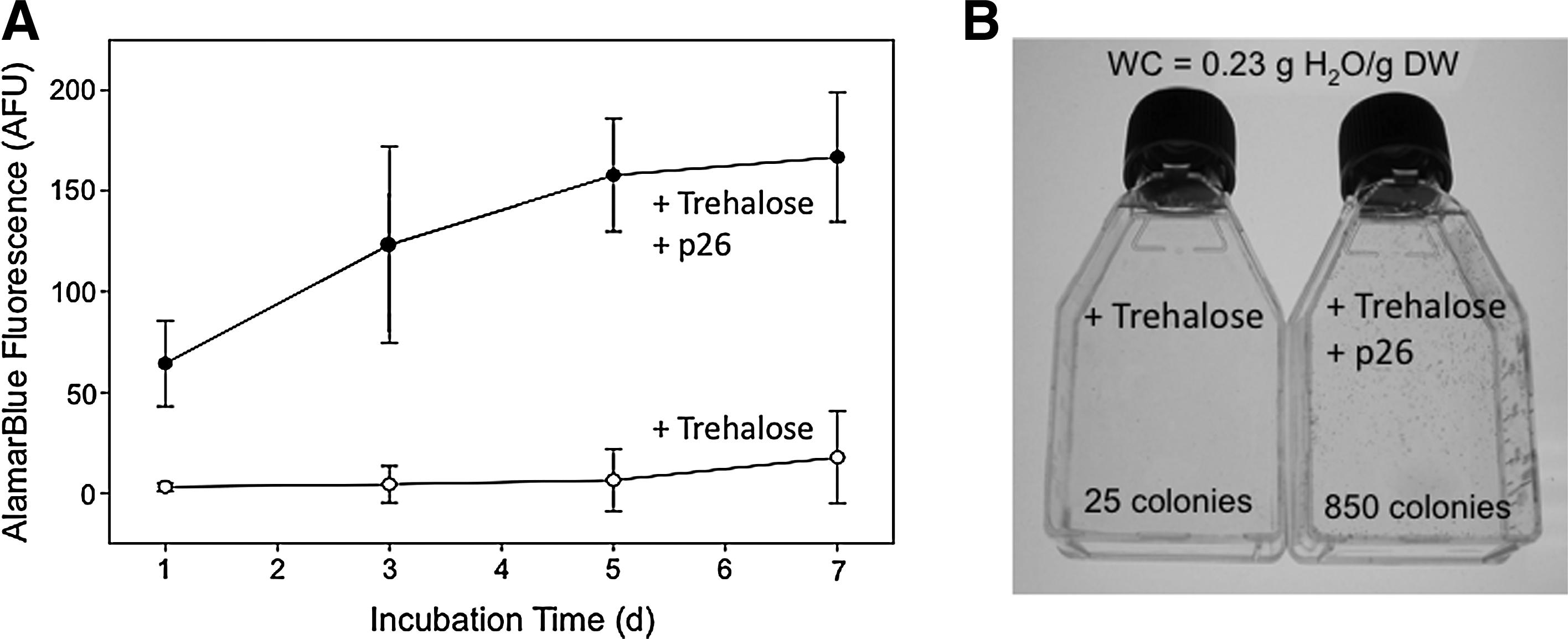
To determine whether p26 could aid cell survival following dehydration, embryonic kidney 239H cells were transfected to express p26. Cells loaded with trehalose and expressing p26 survived to a greater extent than sham-transfected cells loaded with trehalose (about a 2-fold difference at 0.25 g H2O/g DW). 45 However, the more striking finding was that p26-expressing cells recovered better over time following rehydration. At 7 days post-rehydration, the cells expressing p26 and loaded with trehalose were more metabolically active and formed as many as 34-fold more colonies than cells loaded with trehalose alone (Fig. 3). 45 These results indicate that HSPs protect cells beyond the initial rehydration event, and suggest that they may confer a distinct advantage during the recovery phase following rehydration.
Another example of HSPs correlating with protection during drying was seen in HeLa cells. Although all HeLa cells should be genetically identical, significant differences are now seen between strains maintained at different repositories, as the cell lines have diverged over the decades. 57 HeLa cells from the American Type Culture Collection (ATCC) were compared to those from the European Collection of Cell Cultures (ECCC) with regard to their ability to survive dehydration. 46 Interestingly, cells from the ATCC, which showed a nearly 10-fold higher level of HSP70 and HSP27 expression than those from the ECCC line, also displayed dramatically higher viabilities and lower levels of intracellular reactive oxygen species (ROS) following drying and rehydration. 46 This correlative evidence provides additional support for the role of heat shock proteins in preventing cellular damage during dehydration.
Several compounds, such as aspirin and salicylic acid, are known to induce the expression of endogenous HSPs in mammalian cells.58–62 Arbutin, a compound found in high concentrations in dry leaves of the resurrection plant Myrothamnus flabellifolia,63,64 has a structure chemically similar to aspirin and salicylic acid (Fig. 4A). Thus, arbutin was evaluated for its ability to induce heat shock proteins in human MSCs. 48 Indeed, Western blot analysis revealed a dose-dependent induction of HSP70 when MCSs were incubated in medium containing arbutin for 24 h. 48 In addition, though survival immediately following rehydration was unaffected by arbutin, metabolic activity and ability to differentiate (Fig. 4B and C) were significantly enhanced (4-fold and 25-fold, respectively) in samples dried to 0.3 g H2O/g DW with arbutin + trehalose, compared to cells that were treated with trehalose alone. 48 This result correlates well with the p26 study, in that the full protective effect was realized during the recovery phase (∼1 week) following rehydration.

The mechanism of protection afforded by arbutin is likely to be complex. In addition to inducing endogenous HSPs, arbutin is also a strong antioxidant, 65 which could help to safeguard the cells against damaging ROS present under dehydrating conditions.66,67 Furthermore, arbutin has varied effects on membrane structure, from depressing the Tm of dry lipid, and inhibiting enzymatic lipid hydrolysis, 68 to stabilizing the lamellar structure in membranes containing nonbilayer forming lipids. 69 Even the presence of HSP70 has varied consequences. Besides the well-known “chaperone” function of maintaining proper protein folding,70,71 HSP70 also inhibits apoptosis.72,73 It is reasonable, therefore, that the increases arbutin allows in cell health following drying and rehydration are likely to be due to a complex set of interrelated activities and involve multiple pathways.
Inhibition of Apoptosis
As mentioned, one important function of HSPs is the inhibition of apoptosis, or programmed cell death.72,73 Although apoptosis has been well studied with regard to cells under stress from heat, 74 radiation, 75 exposure to ROS and heavy metals,76,78 loss of nutrients, 76 and cryopreservation, 79 it is also becoming apparent that apoptosis is a major damage process in cells undergoing dehydration as well.47,80 A recent study with CANARY cells specifically addressed the inhibition of apoptotic damage, and its resulting effects on viability and functionality of nucleated cells following drying and rehydration. 47
CANARY cells are clonally-derived murine B cells that have been selected to detect a specific pathogen, and were genetically engineered to express cytosolic aequorin. 81 The designation “CANARY” indicates their use for “Cellular Analysis and Notification of Antigen Risks and Yields.” Upon stimulation by antibodies to the appropriate pathogen, intracellular calcium concentration increases, causing the emission of bioluminescence. 81 These cells, which respond in seconds, represent an exciting development in the detection of various infectious pathogens and bioterrorism agents. A stable dry cell product, capable of being stored at room temperature, would vastly increase the impact of this tool, due to improved ability to transport and utilize the cells in remote locations without the necessity for refrigeration or freezing equipment.
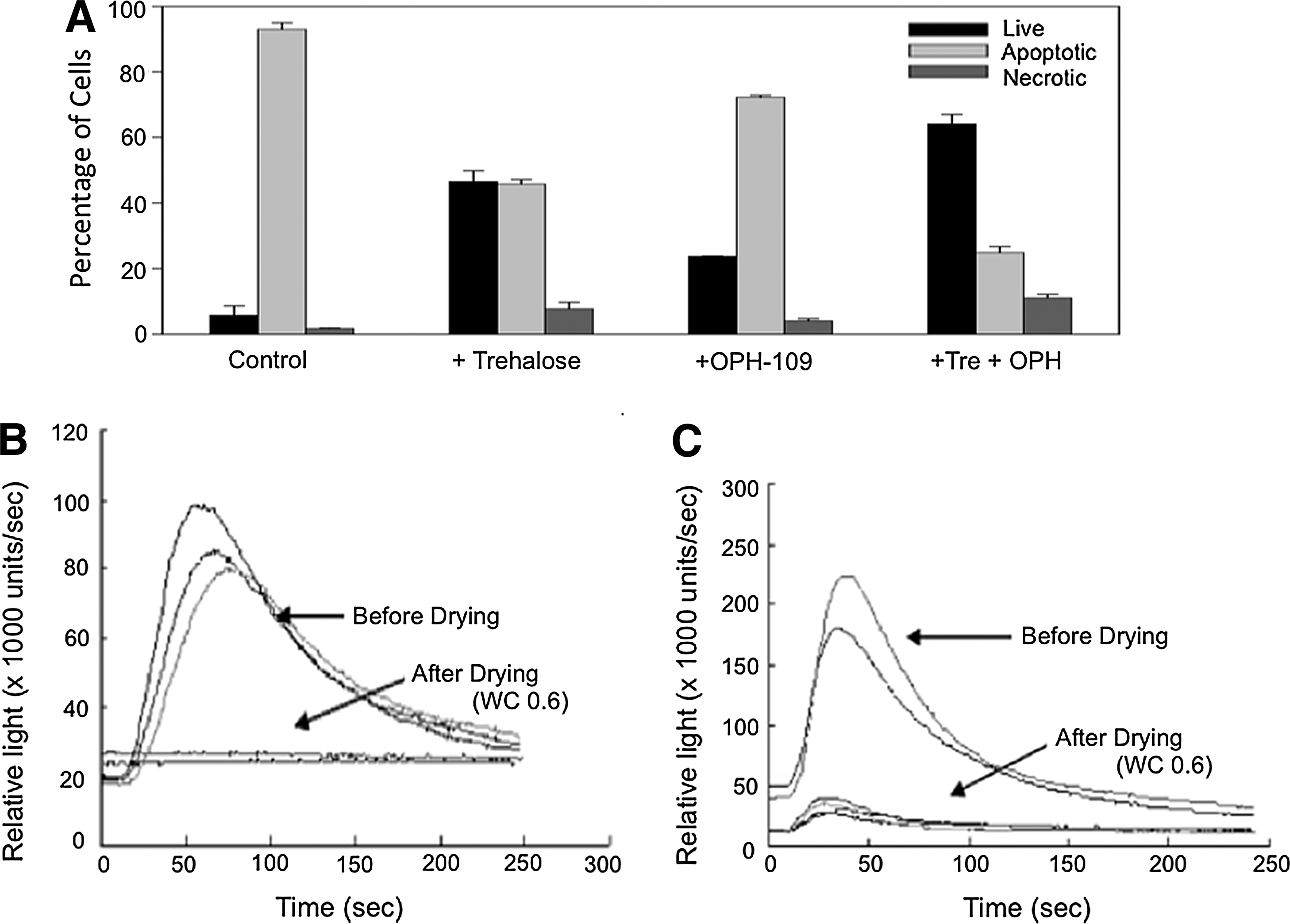
Similar to other cell types, drying CANARY cells without protective solutes resulted in low viabilities even at relatively high water contents, and apoptosis was identified as an important source of damage during this process. 47 Trehalose and a commercially available pan-caspase inhibitor, OPH-109, were tested individually and in combination for their ability to inhibit dehydration-induced apoptosis, and improve post-rehydration viability. Interestingly, trehalose alone provided better protection than OPH-109 alone, but the lowest levels of apoptosis and highest viabilities were obtained when trehalose and OPH-109 were both present, suggesting a synergistic effect (Fig. 5A). 47 The trehalose-protected cells were also competent to emit bioluminescence when appropriately stimulated during the CANARY bioassay (Fig. 5B and 5C), but trehalose did not inhibit apoptosis caused by other types of stress (elevated temperature, camptothecin), 47 indicating that trehalose may offer primarily structural protection, a crucial factor in reducing apoptosis in nucleated cells during drying.
Baust et al., also investigated the apoptotic contribution to cell damage during drying. 80 In this study, specific inhibitors to caspases 3, 6, and 9 were included individually during the drying of hepatocarcinoma C3A cells. The findings demonstrated that caspase-3 inhibition resulted in the greatest increase in post-rehydration recovery, but that inhibition of caspases 6 or 9 was not effective. 80 This result makes sense in light of the fact that caspase-3 activates the “execution phase” of apoptosis, serving as a common step for the many pathways of the apoptotic cascade. 82 That is, regardless of which pathway(s) is(are) involved in dehydration-induced apoptosis, blocking caspase-3 will be likely to inhibit cell death by this process.
As the inhibition of apoptosis and the presence of heat shock proteins have each been shown individually to improve the stress tolerance of nucleated cells, a reasonable approach would be to combine these protective effects. Indeed, an investigation into the storage of CANARY cells has revealed that, after 2 weeks' storage time at room temperature (RT), activity in the bioassay was higher for cells that had been transfected with the genes for heat-shock proteins p26 and artemin (from Artemia) as well as the anti-apoptotic gene Bclw, as compared to cells transfected with either p26/artemin or Bclw alone. 83 The physiological effects and interactions between these various proteins are complex, however, as illustrated by the finding that the p26/artemin combination actually seemed to be injurious when the cells were stored at 4°C. 83 Nevertheless, this study shows that the stress response and apoptotic inhibition can be used in concert to prolong storage of nucleated cells, independent of the need for refrigeration. And while this particular study was conducted on hydrated cells, the striking finding that 75% of the RT-stored cells were active in the bioassay after 2 weeks (whereas the standard storage time for cells at RT is only 2 days) 83 suggests that this combination approach could yield potential benefits for dry cells as well.
Late Embryogenesis Abundant Proteins
Another class of proteins that have been linked to protection from environmental stresses is the group known as Late Embryogenesis Abundant (LEA) proteins. LEA proteins were originally described in cotton and wheat (for review, see Ref. 84), but have been found in many other plants as well.84–86 In addition, LEA proteins have more recently been described in prokaryotes and animals.87–90 Though their exact function is yet undetermined, they are thought to provide protection during freezing and hydration stresses.84,85,89,91,92 For instance, the LEA protein HVA1 from barley, when expressed in wheat or rice, increases resistance to conditions of water deficit.93.94 In fact, recent work demonstrates that LEA proteins can protect sensitive enzymes from desiccation- or freeze-induced aggregation in vitro, and that the effect is synergistic with that of trehalose. 95
In addition to their effects on proteins, LEA proteins may protect lipid bilayers as well. The molecular interaction between model membranes and the LEA protein LEAM, which accumulates in the matrix of mitochondria from pea (Pisum sativum), were investigated using Fourier transform infrared (FTIR) and fluorescence spectroscopies. 96 FTIR evidence demonstrates a direct interaction between LEAM and the phosphate (P=O) groups in dry phospholipids, leading to higher mobility in the fatty acyl chains. 96 Further, LEAM inhibited leakage of trapped solutes from dry vesicles, an effect that was significantly augmented in the presence of the mitochondrial lipid cardiolipin, suggesting LEAM may function to protect mitochondrial membranes during desiccation stress. 96 In support of this notion, the presence of a mitochondrially targeted LEA protein (AfrLEA3m) has been shown to correlate with resistance to dehydration stress in mitochondria isolated from Artemia franciscana embryos, whereas such resistance was not found in mammalian mitochondria lacking AfrLEA3m. 97
A third protective effect may result from the ability of LEA proteins to participate in vitrification of disaccharides. When a LEA protein isolated from Typha latifolia pollen was dried in combination with sucrose, a dose-dependent increase in the glass transition temperature was seen, an effect that correlated with an increase in the average strength of hydrogen bonding within the glassy matrix. 98 Taken together, these studies point toward an important role for LEA proteins in the ability of nucleated cells to survive desiccation. Moreover, as several of the above mentioned techniques have synergistic effects with each other, the most successful strategy for preserving nucleated cells for dry state storage may involve a combination of protective approaches in a single cellular system.
Beyond Protectants: Modifying the Drying Protocol
While the improvements to post-rehydration viability and function, based on loading or expressing certain biomolecules has been significant, additional advances are likely to be achieved by optimizing the drying protocol itself. For instance, vacuum drying results in higher viabilities than convective (i.e., air) drying, likely due to lengthy drying times, inhomogeneous distribution of residual water, and difficulty controlling the final moisture content in the air-dried samples.35,45,99 In addition, controlling a single important parameter, that of buffer osmolarity, has been shown to strongly affect rehydration outcomes. 100 When 3T3 fibroblasts were dried from an isotonic buffer solution, they demonstrated enhanced survival and growth following drying and rehydration, compared to those dried from a hypertonic buffer, even though the trehalose concentration was the same in both (0.2 M). 100
This clear indication that reducing osmotic stress could improve cellular recoveries prompted other attempts at minimizing the cells' exposure to hypertonic conditions. For instance, using microwave radiation to enhance evaporation rates, drying times were reduced by as much as 8-fold, and the distribution of residual water was more homogeneous, as compared to similar samples dried in a dry box. 44 Even more striking, spin-drying CHO cells from a trehalose solution, over a period of <1 min, resulted in residual water contents in the range of 0.2 g H2O/g DW and viabilities in the range of 95%. 101
It should be noted that under very specialized conditions, nucleated cells have been dried to water contents below 0.2 g H2O/g DW. Mononuclear cells (MNCs) derived from human umbilical cord blood were freeze-dried using a directional freezing apparatus, which operates by passing the sample through a defined thermal gradient. 102 Using a protective combination of trehalose and epigallocatechin gallate (EGCG), post-rehydration viabilities in the range of 90%, which fell to 65% after washing, were found in samples dried to ∼4% residual water. 102 Although such samples would not be in the glassy state at room temperature (Tg was measured at 11.8°C), vitrified storage would be possible at 4°C. 102 This patented technology is currently available through collaboration with the company responsible for its design and manufacture.
Silica-Based Protection
The studies described thus far are based on the common theme of carbohydrate-based protection, with various enhancements afforded by other biomolecules. However, vitrification of other compounds is also a possibility. And just as the use of trehalose, one of nature's best tactics for enabling anhydrobiosis, has become widespread in the effort to preserve sensitive biomolecules in the laboratory, other similar lessons are still available. One possible strategy might be adapted from the diatomaceous algae, which are highly resistant to environmental and physical stresses (Refs. 103, 104 and references therein). Diatoms are unicellular algae encased in a protective cell wall (frustule) composed of silica glass, 105 which among other possible functions imparts mechanical strength. 106 Recently, the idea that encasing sensitive biological materials in silica glass could impart stress resistance has been garnering attention. Because of the thermodynamic stability of the Si-O bond (452 KJ/mol−1), 107 this exciting partnering of organic and inorganic materials might make possible cell- or protein-based devices inaccessible by other means.
Although an exhaustive account of silica-based protection of biologicals is beyond the scope of this article, excellent reviews are available.107–109 Isolated proteins, including bacteriorhodopsin,110,111 and phycoerythrin 112 have been encapsulated within silica glasses, and retain their optical properties and enzyme function. Even whole cells, including bacteria,113–115 yeast,113,116–118 fibroblasts, 107 and pancreatic islet cells119,120 have been successfully immobilized within a silica matrix. In some cases, the silica-protected cells can be removed from their aqueous environment. For instance, protein expression, as monitored by the appearance of GFP, was detectable in silica-protected yeast cells after 24 h drying time. 113 For bacteria, stress resistance might be further enhanced by delivering “quorum sensing” molecules to the silica matrices. 121 In fact, “quorum sensing” and the associated genetic programming has recently been shown possible in single, isolated S. aureus cells due to silica confinement. 122 Although the cells that have been dried using this method thus far, bacteria and yeast, are naturally quite resistant to drying, current efforts are underway in several laboratories to understand the possible uses for silica-based techniques in the dry state preservation of nucleated cells as well.
Summary
The benefits of achieving successful preservation of nucleated cells in the dry state would be immediate and profound. It would become possible to ship biosensors and cell-based screening devices to remote locations where conditions limit access to bulky refrigeration and freezing equipment. In addition, routine storage of cells and stem cell-based regenerative medicine products could be conducted at room temperature, yielding enormous savings in money, energy, and release of nitrogen (an important greenhouse gas) into the environment.
Most efforts to stabilize nucleated cells for dehydration involve an initial step of loading the cells with a protective disaccharide, particularly trehalose. Several methods have been used successfully to incorporate trehalose into the cytosol, including fluid phase endocytosis, insertion of pores, nanocapsule delivery, and genetic engineering. Although these techniques differ in their levels of invasiveness, all have generated an improvement in cellular viability and function following drying and rehydration.
Additional protective treatments have also been explored. Heat shock proteins, either expressed in transfected cells, or through induction of endogenous pathways, have improved cellular recovery and function following rehydration. Furthermore, apoptosis inhibitors and LEA proteins could prove to be of considerable importance in protecting nucleated cells, including the many varied and complex intracellular structures, from the severe stresses associated with dehydration.
Finally, the drying method itself is a crucial component of the process and will invariably have a large impact on the post-rehydration viability. Recent studies have shown exposure to hypertonic conditions to be damaging. Thus, rapid drying methods, which minimize this type of exposure, are likely to result in improved cellular outcomes. Nevertheless, in most cases thus far, residual water continues to be necessary in order to ensure survival following rehydration. Additional areas that merit consideration include reducing damage incurred through autophagy, necrosis, and the presence of reactive oxygen species, as well as improving the stability of the cytoskeleton, DNA, and intracellular organelles.82,123,124 In some cases, alternate strategies, such as silica-based protection, may prove valuable. It is quite likely that the successful approach will utilize a combination of the various techniques described.
In conclusion, recent progress has enabled nucleated cells to be significantly stabilized using a variety of techniques. Storage lifetimes in the stabilized, unfrozen state can be as long as several weeks, which is a considerable improvement over unprotected cells. The primary remaining challenge will be to consistently achieve high levels of viability and proper physiological function after the removal of residual water, such that vitrification becomes possible at room temperature.
Footnotes
Acknowledgments
The author acknowledges John H. Crowe and Fern Tablin, directors of the UC Davis Center for Biostabilization. Additional thanks to Atul N. Parikh, C. Jeffrey Brinker, and Blaine Butler for stimulating discussions regarding silica-based preservation. AEO also acknowledges current support from the US Department of Energy, Basic Energy Sciences, Division of Materials Science & Engineering under Award No. DE-FG02-04ER46173, through a subcontract from Los Alamos National Laboratory.
Author Disclosure Statement
No competing financial interests exist.