
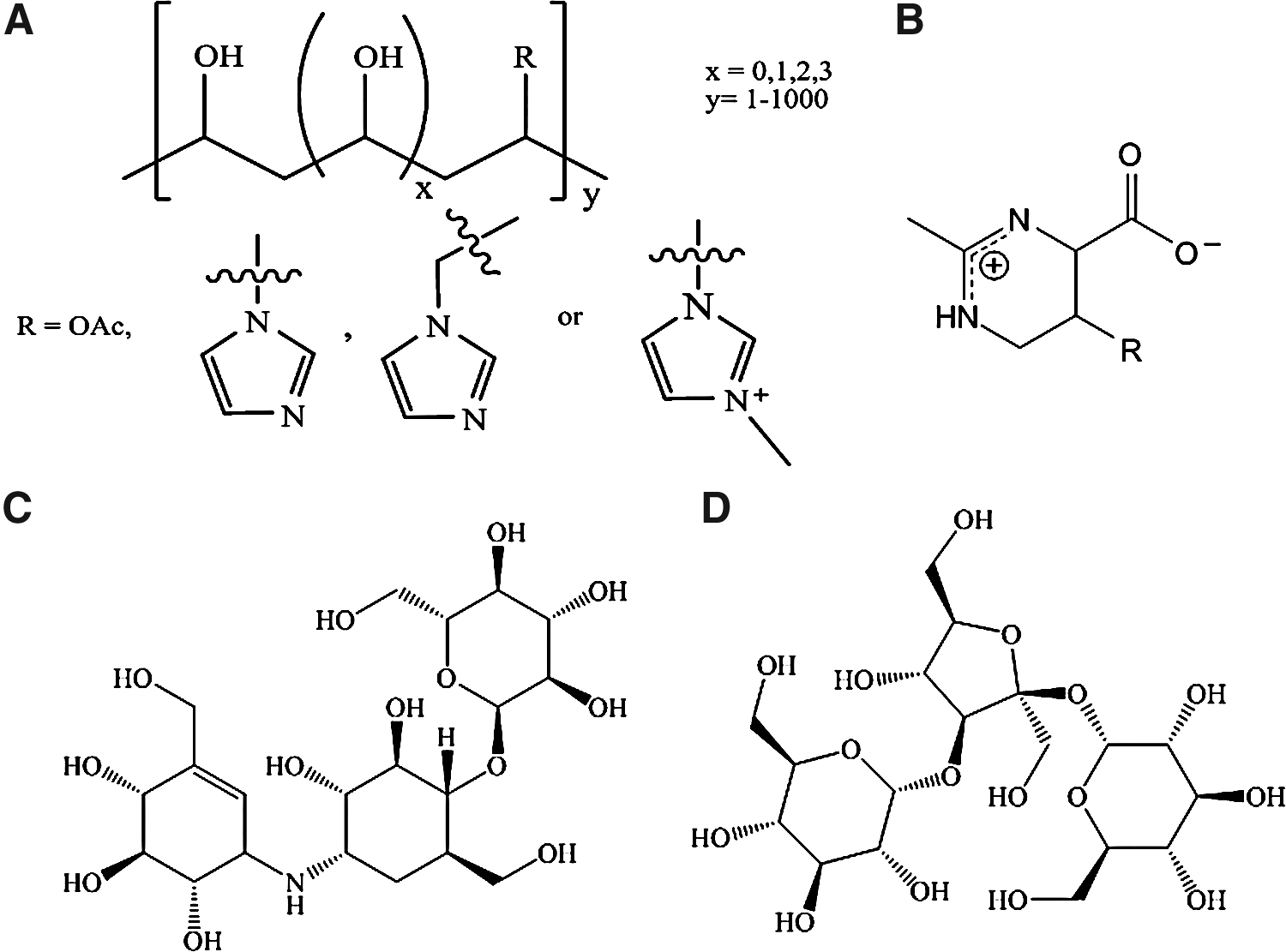
Abstract
We describe here a unique approach to identifying small molecules that are useful in stabilizing biomolecules in the dry and liquid states. Using biostability screens aided by in silico docking experiments and synthetic chemistry, libraries of biostability molecules are analyzed for their ability to protect important biological materials. In the case of DNA stabilization in the dry state, interactions of suitable candidate biostability compounds with DNA are studied in initial screens to identify their ability to form glasses at elevated temperatures. The most promising compounds are then tested for their capacity to preserve DNA during long-term storage. The results have led to a commercial product for storage of DNA that is being adopted for many commercial and experimental applications. Further studies have shown that small molecules for preservation of RNA, proteins, and tissue samples can be developed by following the same progression of screens.
The Need for Stabilization of Biomolecules
Currently, biological materials are usually preserved in the frozen state. The science of cryobiology has advanced over the past 50 years, with “cold-chain” logistics made more reliable through technical improvements in freezers, utilization of tracking systems for recording sample temperature fluctuations, development of sophisticated alarm systems, and improved and standardized biospecimen handling procedures. Nevertheless, even with these improvements, biospecimen quality remains permanently at risk, either through slow, inexorable loss of specimen integrity over time, or by rapid loss of specimens through storage system failures and freeze/thaw degradation. 9 Therefore, “cold chain” systems have both high direct costs associated with their establishment and maintenance, and considerable hidden costs associated with the risk of specimen loss. To address these problems, we have developed a way to stabilize biomolecules at room temperature that provides an effective alternative to freezing, and that effectively addresses the cost associated with cold-chain transport. 10
Biomolecule Stabilization
At the start of this work, the aim was to develop methods for room temperature storage of DNA, which is currently stored in −80°C freezers or in liquid nitrogen, at enormous cost. Clearly, providing a way to obviate the need for freezers to store DNA would be a worthwhile endeavor, and as is shown below, we have been successful in this regard. Over time, we have developed processes for field collection, preservation, transport, analysis, and long-term storage of biomaterials such as tissue samples, blood, urine, sputum, nasopharyngeal swabs, and sweat (Fig. 1). The materials are sufficiently well preserved that they are suitable for genome, transcriptome, and proteome analysis.

Biospecimen samples are collected in the field, instantly preserved, and specific analytes are stabilized and made ready for identification for analysis.
Principles of biostabilization technology
Our biostability platform technology and product development process focuses on the stabilization of biomolecules such as DNA, RNA, metabolites, proteins, whole cells, tissue samples, tissue sections, and assays at ambient and elevated temperatures (see www.biomatrica.com for further details). The technology is a direct result of studies on the biology of extremophiles, particularly those that exhibit anhydrobiosis (“life without water”). 11 A common feature of such organisms is that they produce large quantities of disaccharides, the most well-known of which is trehalose.12,13 Over the past three decades, numerous groups around the world, particularly the Crowe laboratory, have shown that trehalose has the capacity to stabilize membranes, 14 proteins, 15 and even intact platelets16,17 in the dry state. The consensus on the mechanism by which this stabilization is accomplished is that the sugar forms a glass in the dry state, 13 which inhibits close approach of the biomaterials during storage. 16 However, glass formation is not the entire story; many molecules that form glasses are ineffective in conferring a stable state during dehydration, indicating that something else is required. The bulk of the evidence suggests that direct interaction between the stabilizing molecule and the biomaterial is required. 13 This direct interaction, most likely made by hydrogen bonds, confers on membranes a physical state that resembles that seen when the material is fully hydrated. 17 This phenomenon is known as the “water replacement hypothesis”. 18
In our first entry into this field, we discovered that trehalose stabilizes DNA in the dry state (discussed below). When docking experiments between these two components were conducted, it became apparent that in the dry state, trehalose is most likely situated in the minor groove, where it is probably hydrogen-bonded to polar residues in the backbone, although evidence on this point is still uncertain.
Having identified potential stabilizers in silico, we then examined whether these molecules could stabilize real biological materials (Fig. 2). Using large libraries of stabilization molecules, including chemical modifications, we hypothesized that some molecules would be able to surpass the stabilizing effects of trehalose. We started with a small molecule that forms glasses in the dry state, conducted an in silico screen for interactions with DNA, and then tested its ability as a stabilizer in vitro. This has resulted in a library of custom-engineered stabilizers (Fig. 3), and the advent of the SampleMatrix® Technology. This approach has been remarkably successful, beginning with DNA stabilization, which resulted in our first commercial product, DNAstable®.

Our core technology is based on small molecules synthesized to perform specific interactions with the biological material that has to be stabilized. In the early stages, Autodock, the technology development docking prediction program, has been used to understand the interactions of the stabilization molecules with the surface of the biological material. 37 Trehalose (center blue molecules), the best known natural biostabilization molecule, is following the binding pattern of water molecules (left, blues spheres); the right panel (stabilization molecules in green) demonstrates that synthetic polyol based stabilization molecules follows the same binding pattern as trehalose. Molecular modeling was used to predict interactions of Sample Matrix with nucleic acid molecules. This approach allowed for rapid in silico screening of potential binding molecules and predicts their potential suitability to stabilize biomolecules.
Successful stabilization molecules, next to the well-documented trehalose molecule, have been found during the screening process.
Some of our initial formulation work focused on finding materials with glass transition temperatures (Tg) far above room temperature. However, the work of Ciccerone and Soles 19 has shown that inclusion of low molecular weight plasticizers in formulations actually produces improvement in protein stability greater than 100-fold over the unplasticized glasses. We have found that certain polymers incorporating small molecules in selected molar ranges can yield materials capable of protecting DNA and RNA for long periods of time at ambient and elevated temperature conditions. Further extensions of this work are expected in the immediate future to yield improved stability of proteins, bacteria, viruses, and hopefully intact mammalian cells in the dry state.
Protection of genomic DNA
To investigate the capacity of our biostability compounds to protect DNA, various amounts of human genomic DNA (500 ng, 200 ng, 100 ng, 50 ng, and 4 ng) were mixed with DNAstable in microfuge tubes and allowed to dry in a laminar flow hood. A control group of genomic DNA was treated identically but without addition of DNAstable. Samples were then placed in an Erlenmeyer flask, covered with foil, and subjected to a dry cycle autoclave run. Autoclaving proceeded at 250°F/121°C for 15 min at 15 lb/in 2 and dried for 30 min at 150°F/66°C. Samples were removed from the autoclave and allowed to cool. The DNA was re-hydrated in 10 μL of water for 15 min at room temperature. Samples were used to amplify the human β-actin gene in a polymerase chain reaction (PCR) in a final volume of 30 μL. Cycling parameters were initial denaturation at 94°C for 5 min, followed by 40 cycles of 94°C for 15 sec, 55°C for 30 sec, and 72°C for 30 sec. Ten microliters of the PCR reactions were run on an agarose gel containing ethidium bromide. The results indicate that storage of DNA in SampleMatrix under extreme conditions (e.g., autoclave) maintained DNA integrity as evidenced by the generation of gene-specific PCR amplification products of the expected size. In contrast, unprotected DNA did not yield any amplification product indicating that the DNA was destroyed by the autoclaving (Fig. 4). 20

Protection of DNA in SampleMatrix under extreme conditions. Aliquots of 500 ng gDNA were PCR amplified using human β-actin primers after storage of unprotected DNA in the freezer (U) or DNAstable (SM) protected DNA stored at room temperature (left). Various amounts (500 ng; 200 ng; 100 ng; 50 ng; 4 ng) of gDNA were dried with DNAstable or without (unprotected) and subjected to autoclaving. Hydrated DNA was PCR analyzed using human βactin primers. NC: no template control (right).
Similar experiments have demonstrated the ability of our biostability compounds to protect DNA during long-term storage in liquid form. In these experiments, genomic DNA isolated from blood or saliva was mixed with DNAgard® and placed into storage at either room or elevated temperatures (50°C). 21 An equivalent set of samples that did not contain the biostability compounds were constructed. As positive controls, portions of the genomic DNA were placed into storage at −20°C. After 62 days, the integrity of the genomic DNA was assessed using agarose gel electrophoresis and ethidium bromide staining. As shown in Figure 5, the genomic DNA protected by DNAgard (indicated by DGS over lanes) contained high molecular weight species similar to that present in the control samples stored at −20°C. Samples that did not contain biostability compounds failed to demonstrate any staining with ethidium bromide, indicating that the DNA had become completely degraded during storage. Interestingly, the DNAgard-containing sample was free of the smearing characteristic of low molecular weight species. Not only do the results support the notion that the biostability compounds can protect DNA, but they were more effective at doing so than storage at −20°C, long considered adequate for stabilizing nucleic acids. 22
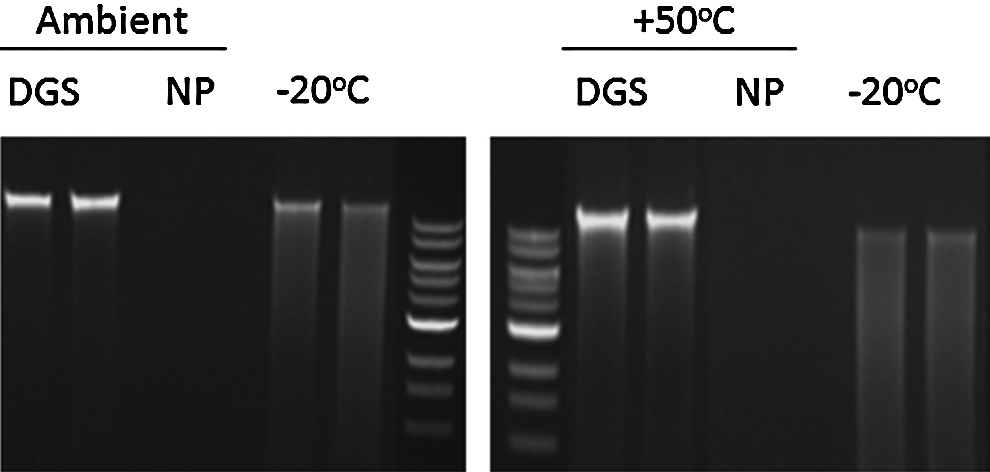
Gel images of genomic DNA isolated from saliva and stored at room temperature or right: at 50°C in DNAgard (DGS) for 62 days prior to DNA isolation. Staining of the DNAgard samples are comparable to the −20°C controls.
The diagnostic implications of storing genomic DNA in the presence of biostability compounds has been evaluated by the Los Angeles County, California Sherriff's Department. 23 Genomic DNA isolated from blood, semen, and buccal samples was mixed together with DNAstable, and stored in the dried state at room temperature without humidity control. Identical sets of genomic DNA were stored in a −20°C freezer. After 45 months, the specimens were rehydrated and tested using quantitative RT-PCR and Short tandem repeat (STR) analysis. For each specimen type, storage in DNAstable at room temperature yielded results that were equivalent to that obtained with DNA that had been stored in the freezer. For two specimen types (buccal and blood), more DNA was recovered from those specimens stored using the Biomatrica SampleMatrix Technology (Fig. 6).
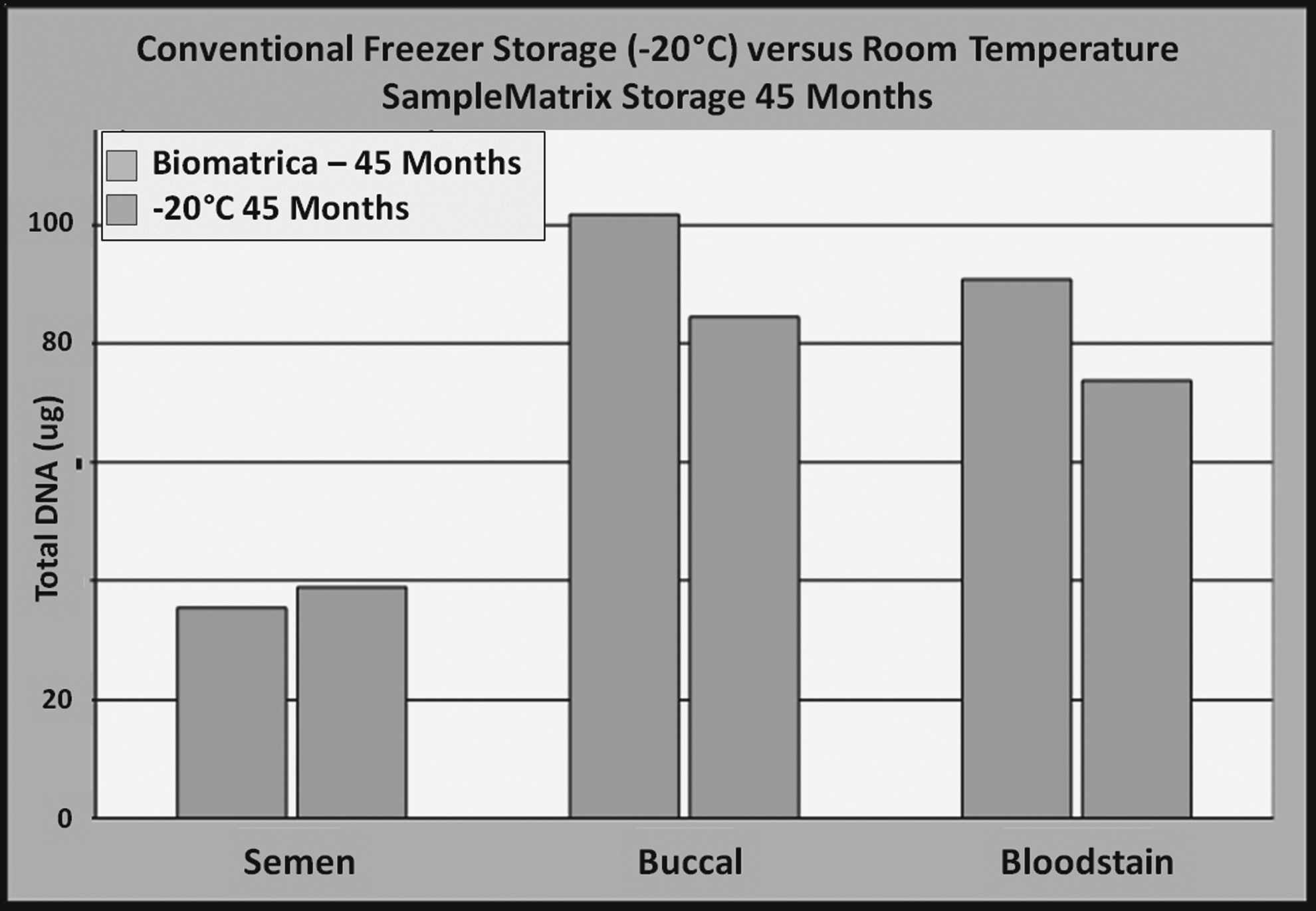
DNA was isolated from semen, blood, and buccal swab samples, divided into freezer controls (−20°C, blue) and stored air-dried in DNAstable® (orange) at room temperature condition in the laboratory (not constant temperature controlled) for 3 years and 9 months. Freezer controls were thawed and dry-stored samples were rehydrated and quantified. No statistically significant difference was detected between room temperature and freezer-stored samples; they were both functional for downstream assays.
Accelerated stability studies conducted with SampleMatrix indicate that gDNA stored in the presence of this stabilizer exhibits greater than 20 years of stability at room temperature. Large-scale evaluations have been performed in the scientific and forensic community, 24 documenting that the stabilization of DNA under environmental and extreme transport conditions is equivalent to the storage of DNA under the best conditions in Teflon tubes in a −80°C freezer. 25 After years of testing,26–28 the technology is being adopted in many scientific, forensic laboratories, and biobanks. 25 The ambient storage method has become the method of choice especially for low concentration samples from forensic evidence.29–32
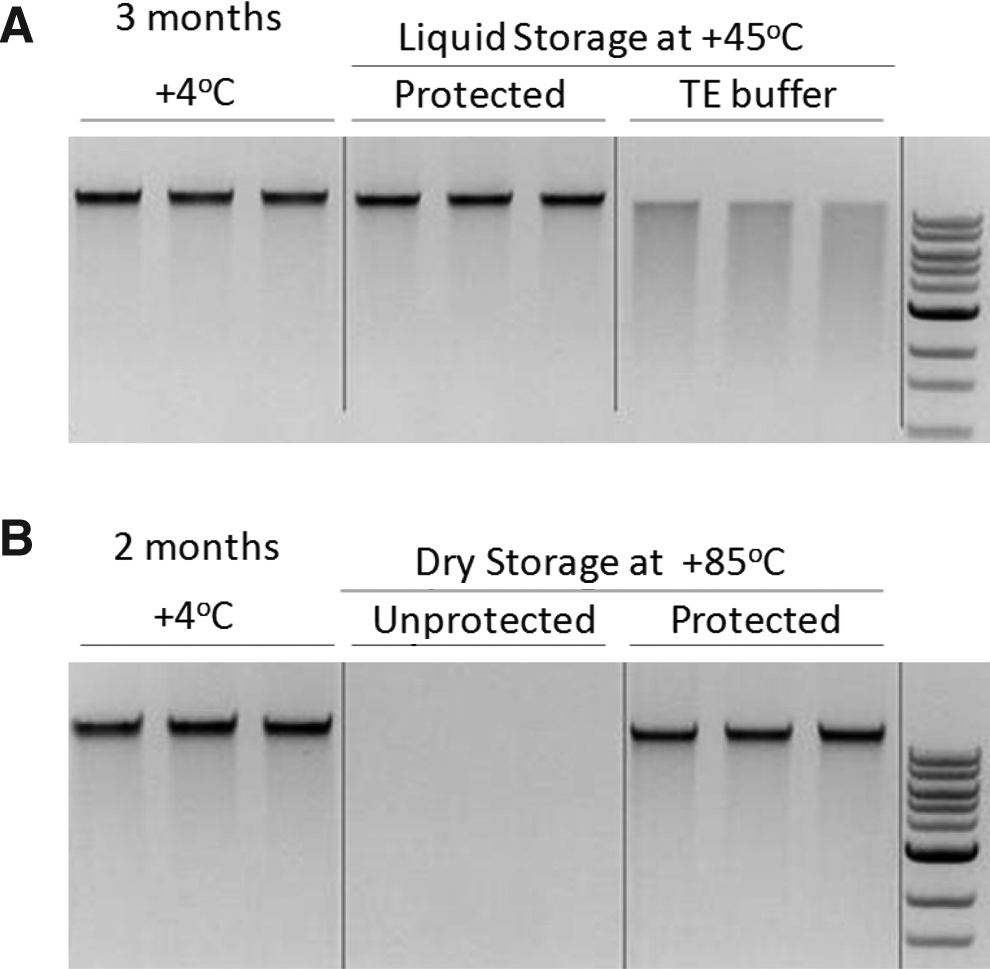
Combining the liquid and dry stabilization approaches, it was possible to develop stabilization matrices that protected DNA in the liquid as well in the dry states (DNAstable Plus®) for more than 12 months at ambient temperature and under harsh environmental conditions (45°C) for more than 3 months, without degradation (Fig. 7A). In addition, DNAstable Plus protects the DNA from constant extreme temperatures up to 85°C (Fig. 7B).
Technology platform for optimizing biostability compounds
In addition to the molecular modeling that led to development of the stabilizers discussed above, several processes derived from drug discovery research were used to identify biostability compounds that improve on those provided by nature. First, an extensive library of biostabilization molecules that includes natural molecules as well as proprietary synthetic molecules was developed by Biomatrica to optimize the stabilization of different biomolecules, either purified or within complex samples such as blood, saliva and tissue (Fig. 8). 33 This medicinal chemistry approach allows us to query uncharted regions of chemical space for biostability compounds with desired properties in a rational manner. The compounds are subjected to rapid, high-throughput screening to identify those that provide suitable biostability. Some screens are virtual, using in silico-based approaches with custom-developed biostability prediction algorithms and docking solutions, as was used for the original DNA work described above. This is complemented by primary screens consisting of specimen-specific assays using temperature-sensitive biomarkers appropriate to the analyte(s) of interest. Candidate compounds can be evaluated alone or as part of a formulation that includes multiple stability components to ensure stability of diverse analytes. Screening is targeted to the desired output, based on pre-determined performance specifications, such as simple recovery of isolated analytes (e.g., pure DNA) or gene expression analysis of a complex biospecimen such as a tissue slice which is to be stored at ambient temperature prior to use. A simple means of differentiating between compounds that perform similarly is to increase the stringency (e.g., duration of storage yielding complete analyte recovery or temperature maximum yielding complete analyte recovery, or both) of the performance specifications used in screening. As with drug development, multiple iterations of the “synthesis and screen” process may be used to derive optimal lead biostability compounds and formulations with the desired properties. Altogether, the process, which provides a means of rapid product development and optimization, has proven successful for Biomatrica regardless of the specimen type or the analyte(s) being stabilized.
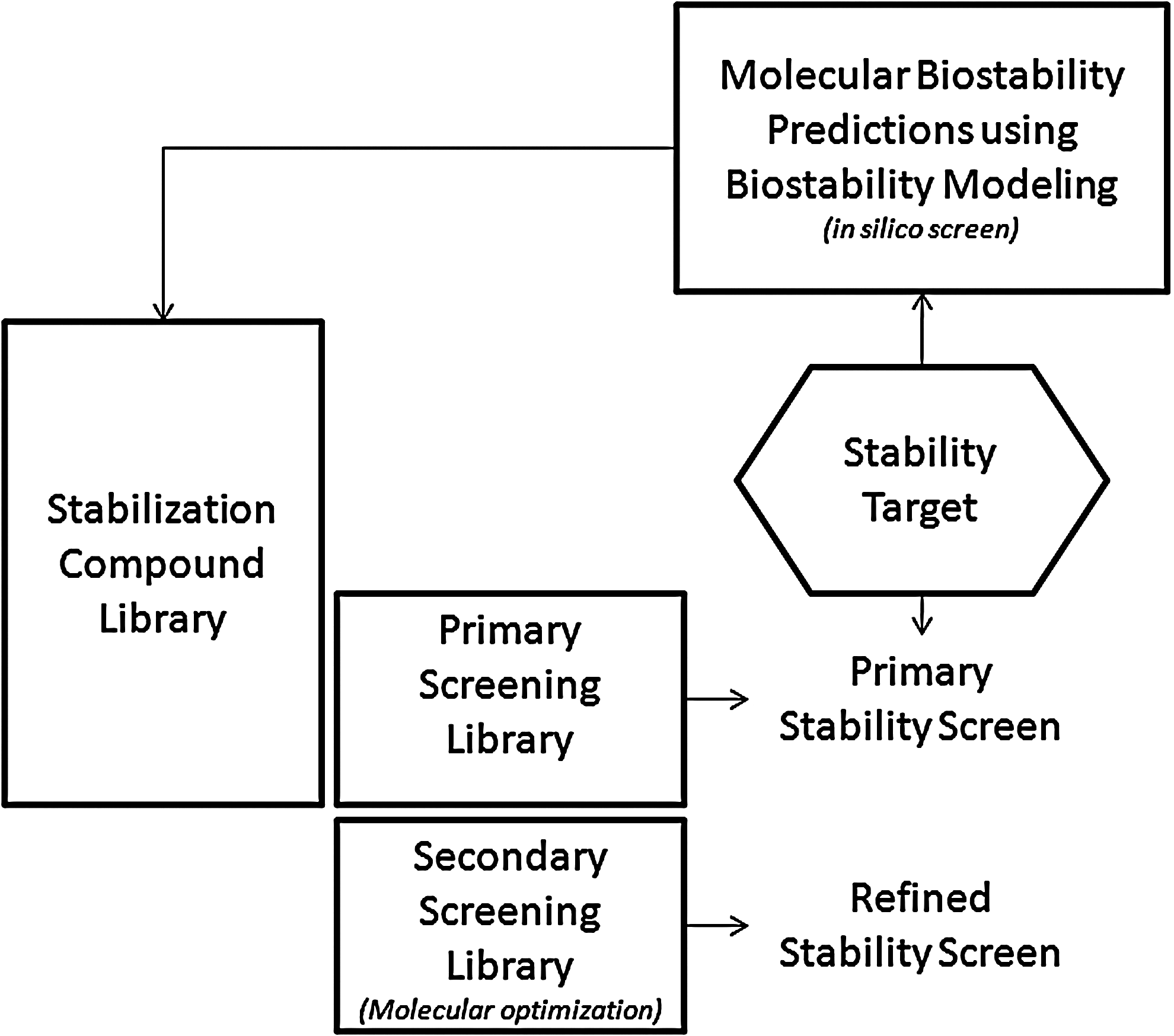
Workflow for the selection of suitable stabilizers from a library of biostability molecules. An example for molecular modeling of water, trehalose, and polyoles into DNA double helix is highlighted in Figure 1. Primary screens are being performed using an existing subset of biostabilization compounds from the library, whereas secondary screens will use structural refined biostability molecules.
Further Application of the Technology: Protection of RNA
RNAstable® was designed to optimally protect purified total RNA isolated from biological samples, such as tissue culture cells and plants. Similar to DNAstable, RNAstable is supplied as a dried medium in the bottom of sample storage devices. To evaluate the product's performance, aliquots (500 ng) of total RNA prepared from human embryonic kidney cell line 293T using the TRIzol® reagent were added to RNAstable in 1.5 mL standard microfuge tubes and allowed to dry for 30 min in a SpeedVac® without heat. A control sample (NP) was prepared by drying 500 ng of total RNA in an empty tube under identical conditions. Samples were then stored for 8 months at either room temperature or at 45°C to simulate accelerated aging conditions (equivalent to 4.5 years storage at room temperature). The freezer control samples were stored at −80°C. Aliquots were run on a 1.2% 1×TAE agarose gel to assess integrity of recovered RNA—and on a bioanalyzer to obtain a RNA integrity number (RIN) score. The results indicate that sample integrity is maintained during dry storage in RNAstable at room temperature. 34 Additionally, specimens stored in RNAstable and incubated at high temperatures showed equivalent results to RNA stored at −80°C. As an example, the study of RNA preservation in RNAstable for 29 months at 45°C showed that RNA integrity is preserved at such storage conditions (Fig. 9).
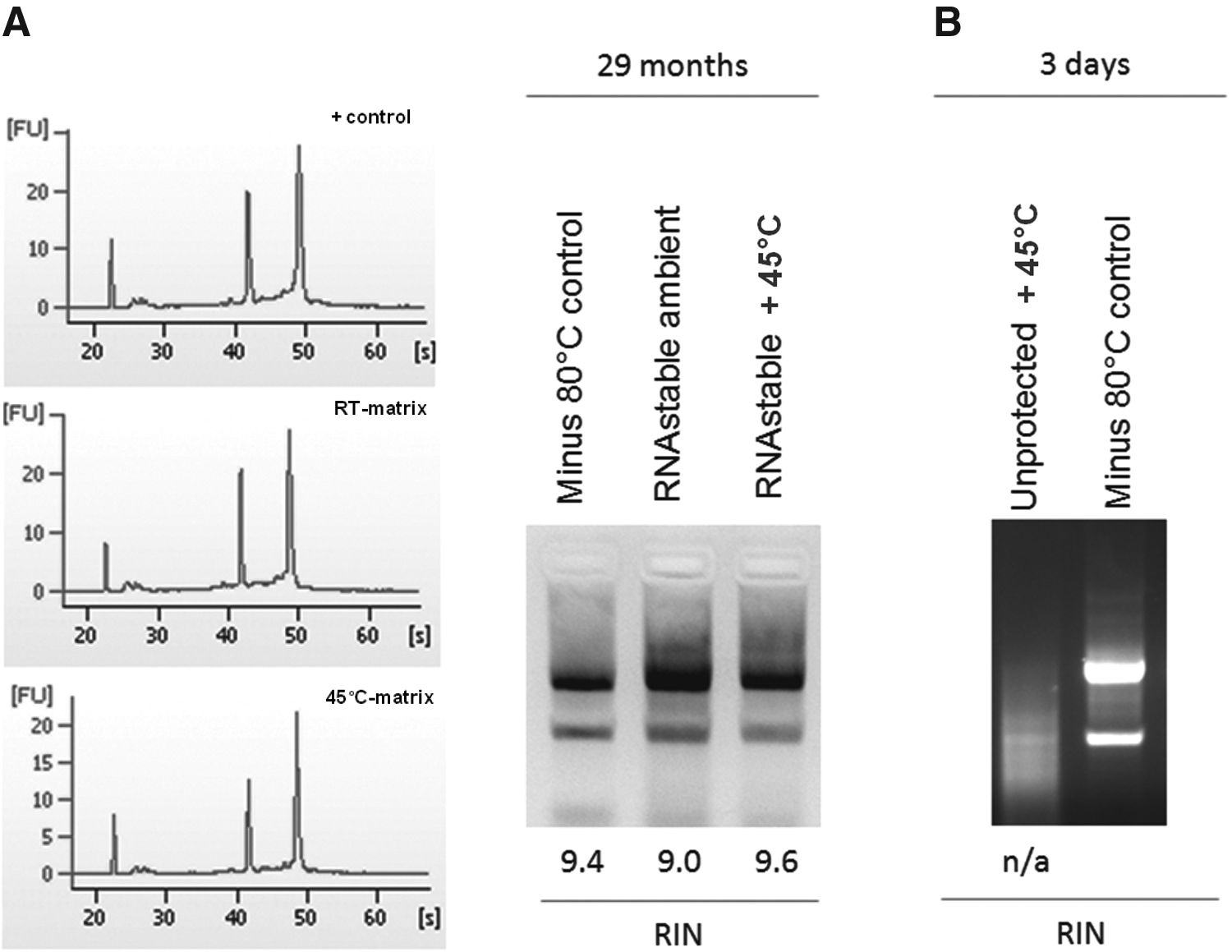
Long-term storage of total RNA at room temperature (RT) and at 45°C under accelerated storage conditions for 29 months. Bioanalyzer results including RIN score analysis demonstrate equivalent RNA quality under room temperature and high temperature storage compared to ultra-low temperature (−80°C) storage. Accelerated aging at 45°C for 29 months represents ∼12 years of RNA preservation (Arrhenius equations).
Protection of Proteins
Protein and assay stabilization is very important for research and diagnostic and drug development. Proteins can be stored at very low temperatures, but freezing induces large stress on protein molecules. 35 Long-term preservation of protein in the solid or dried state can be achieved, 36 as well as short-term stabilization in an aqueous environment. Biostabilization of proteins at ambient temperature for use in assay development and to avoid cold chain requirements is feasible and has been used successfully by Biomatrica and others to protect proteins (Fig. 10).

Storage of Taq polymerase (Promega) in the dry state for 2 years and 6 months at either ambient (room) temperature or at 50°C. The stabilized sample (Protected) and nonstabilized enzyme (Unprotected) indicates the sample stored in the dry state without stabilization matrix. After storage, the protein was used to perform an endpoint PCR assay amplifying a 490 bp DNA fragment (arrow) from a genomic DNA template. Similar performance can be achieved for many enzymes as well as complete enzymatic assays (data not shown).
The stabilization of biomarkers in whole blood during long-term storage at ambient temperatures is technically challenging. Proteins in plasma are inherently susceptible to denaturation and/or degradation. In order to determine if the biostability compounds could maintain proteins in a native state compatible with downstream analysis and prevent degradation by proteolytic cleavage or chemical modification, we spiked plasma, either in the presence or absence of biostability compounds, with a collection of 30 hormone-like proteins (cytokines), and stored the resulting samples for 20 days at room temperature. Controls consisted of the same proteins spiked into plasma. Following storage in the liquid phase, samples were evaluated for residual activity using a protein-specific ELISA, and the results expressed relative to the starting signal for each protein at t=0. All 30 of the test cytokines were recovered at 95%–100% of their original input levels. Three of the proteins (INF-g, IP10, and MIP-1a) were selected as representative examples of the stabilization capacity of the biostability compounds in the liquid phase (Fig. 11). By comparison, none of the cytokine samples stored at 4°C without biostability compounds maintained more than 30% of the original input levels, and a majority had less than 10%. These results provide compelling, first-of-its kind data on the stabilization of proteins in plasma. To date, no other technologies have been shown to achieve this level of protection.
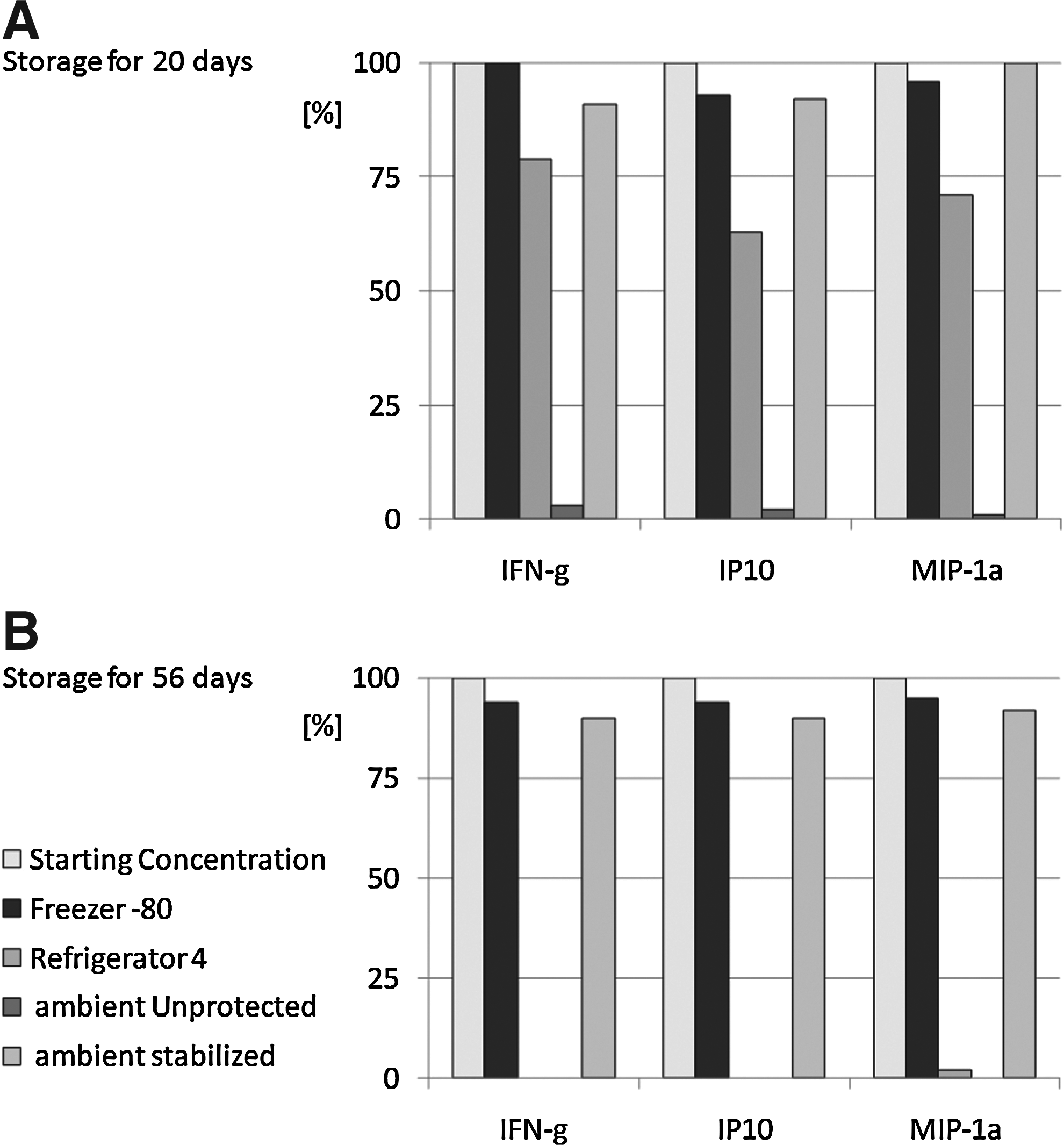
Biomatrica biostability compounds protect cytokines in plasma during room temperature storage. Panomix human cytokine array of plasma spiked with 10 ng/standard after
Parallel Preservation of Molecular Analytes and Morphology in Tissue Sections
Biostability compounds are being developed for the purpose of co-preserving tissue morphology and the analytes contained within the tissues. The concept is that biostabilitity compounds can form the basis of a novel technology for preservation of tissue slices, providing an alternative to the standard methods of formalin-fixed, paraffin-embedded (FFPE) tissue sections or flash freezing. Both of these traditional techniques have drawbacks related to either the retention of morphology or the preservation of analytes within the tissue. Post-processing extraction, amplification, and analysis of cross-linked nucleic acids from FFPE tissues is possible, although it is expensive, yields are poor, and pre-analytical variables are increased. While flash-freezing preserves both nucleic acids and proteins, it impairs morphologic analysis due to varying degrees of histologic distortion, also known as “freeze artifact”.
As a first step in developing biostability compounds for co-preservation, we explored whether these compounds would stabilize RNA and DNA from tissue culture cells stored for extended durations. Cells from human embryonic kidney cell line 293T were either frozen and stored at −80°C, or suspended in solutions containing candidate biostability compounds and stored at room temperature for 14 days. The nucleic acids were extracted and run on an agarose gel containing ethidium bromide. The results demonstrate equivalent recovery of genomic DNA and ribosomal RNA between the two groups of cells (results not shown). Experiments were expanded to mouse tissue (Fig. 12).
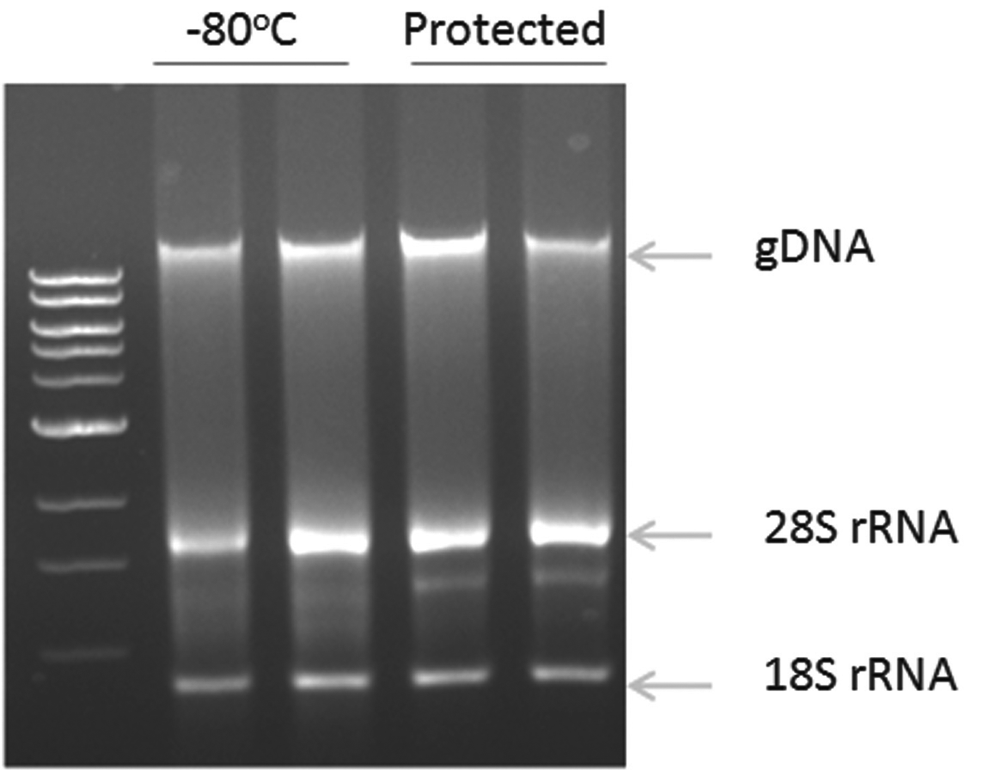
Further validation studies were conducted with two of these biostability compounds in which their capacity to stabilize RNA in tissue was evaluated. In this case, specimens of mouse brain were either quick frozen and stored at −80°C or were suspended in solutions containing the candidate biostability compounds and stored at room temperature for 13 days. At the end of the storage, the RNA was extracted and run on a bioanalyzer to obtain a RIN score. The biostability compounds yielded equivalent RIN scores to the specimens stored at −80°C. In contrast, other (condition) showed changes in the banding pattern and indeterminate RIN indicative of loss of RNA integrity. These results are consistent with the idea that biostability compounds can be used to stabilize nucleic acids contained within tissue specimens prior to extraction.
To address the feasibility of preserving tissue morphology with biostability compounds, a collaborative study that included a central processing site at Biomatrica and multiple clinical specimen collection sites in Asia (China, Japan, Taiwan, and Korea), was conducted. Tissue samples from glioma patients were collected and divided into two equivalent portions. One portion was flash frozen at −80°C and placed on dry ice, while the other was placed into a Biomatrica liquid preservative, DNAgard Tissue®. The specimens were then shipped over 7 days at ambient temperature to the collection site. A portion of each specimen was removed for nucleic acid extraction and the remainder used to produce histology slides. The flash-frozen samples were embedded in OCT (optimal cutting temperature) media before tissue sectioning and slide preparation. The slides from the flash-frozen samples were dried for 60 minutes at ambient temperature; they were not subjected to fixation. The tissue specimens preserved in DNAgard Tissue were processed in an automated tissue processor with a modified initial step in order to avoid tissue exposure to 10% NBF (neutral buffered formalin). As shown in Figure 13, the DNAgard Tissue product preserved tissue landmarks and cellular detail, and tissues showed uniform structural integrity. While the DNAgard Tissue product was not designed for preservation of tissue morphology, these studies give further proof to the concept that biostability compounds can be developed which co-stabilize both the morphology of tissues and the endogenous analytes contained within them. Further studies are in progress to identify optimal biostability compounds for this purpose.

Human glioma tissue, collected fresh in DNAgard Tissue® solution, ambient temperature transport, processed 5 days post collection, paraffin embedded, and sectioned at 4 micron section, H&E stained. H&E stained section shows uniform morphology (left) with retained cellular landmarks (right).
Conclusion
Traditional cryobiology has met with enormous success over the past 50 years, and we have benefited enormously from the wealth of knowledge that is in the literature. However, traditional cryobiology has depended largely on naturally occurring protectants (e.g., glycerol) or on those that were discovered quite by accident. We believe we are charting a new path towards custom design of protectant molecules and technologies that will impact the future of the field.
Footnotes
Acknowledgments
Thanks to Galen Hostetter, M.D., Translational Genomics Research Institute (TGen); Van Andel Research Institute for the histopathology slides; David Goodsell, Ph.D., The Scripps Research Institute (TSRI), for the initial molecular modeling experiments using the Autodock program; Greg Hadinoto, Los Angeles Police Department (LAPD), for the long-term DNA stabilization results.
Part of the research was supported through an NCI SBIR contract HHSN-261201000124C
Author Disclosure Statement
The four authors are shareholders of Biomatrica, Inc.