
Abstract
Insulin-dependent diabetes mellitus is one of the leading causes of death world-wide. Donor-derived pancreas and Islet of Langerhans transplantation are potential cures; however, postmortem ischemia impacts islet quality. The murine βt3 cell line was employed as a model to study cell viability and proliferation after hypothermic storage by comparing Belzer's Machine Perfusion Solution with Unisol™ Solution. The objective was to determine which of these solutions provided the best base line support for βt3 cells and to screen potential cytoprotective additives to the solutions. Initial βt3 cell viability was similar in the two storage solutions; however, better proliferation was observed after storage in Unisol Solution. The caspase inhibitor, Q-VD-OPH, and α-tocopherol improved viability in both storage solutions, suggesting that apoptotic pathways may be responsible for cell death during hypothermic storage of βt3 cells. Analysis of apoptosis markers, caspase activity, and DNA laddering showed a reduction in apoptosis when these additives were included. The effects of Q-VD-OPH and α-tocopherol were also synergistic when employed together during either hypothermic exposure, post-hypothermic physiologic incubation, or combinations of hypothermic exposure and physiologic incubation. These results suggest that both supplements should be included in pancreas hypothermic storage solutions and in islet culture media during post-isolation culture prior to transplantation.
Introduction
In the search for a cure for diabetes, researchers have sought ways to return normal pancreatic function to the body. The methods employed have included whole pancreas transplants, human islet transplants, animal islet transplants, fetal tissue exchange, creation of artificial pancreas or beta cells, and transplantation of genetically-engineered cells.1,2 All of these procedures have both positive and negative attributes. Pancreatic islet transplantation received a strong boost from the introduction of glucocorticoid-free immunosuppressive regimens. As a result, there is now a consensus that islet transplantation may be a viable option for the treatment of insulin-dependent diabetes mellitus. The short-term success of the first glucocorticoid-free protocol3,4 and progress in modification of the protocol for longer-term post-transplant islet function 5 stimulated our search for technologies that may help overcome the shortage of pancreata for islet isolation.
Procurement of live donor pancreata for islet isolation and transplantation is in its infancy. Many pancreata suitable for islet isolation and transplantation are not procured due to concerns about postmortem ischemia. Postmortem ischemia during hypothermic transport on ice results in autolysis of the insulin-producing β-cells in the islets, inadequate islet yields, and poor function. Current practice is to flush and transport the pancreas with University of Wisconsin (UW) Solution on ice. We anticipate that better pancreas preservation may be achieved by perfusing the pancreas during hypothermic storage.6–8 Allogeneic kidneys have been shown to function better after perfusion in a large prospective, randomized, multicenter study. 9 The long-term objective of our studies is development of an optimized pancreas storage solution for hypothermic perfusion of the pancreas, with preservation of the Islets of Langerhans for transplantation.
To this end, we employed a murine β cell line as a model to study cell viability and proliferation after hypothermic storage to compare the lead commercially available organ perfusion solution, Belzer's Machine Perfusion Solution (BMPS), with a new proprietary solution, Unisol™.10,11 The objective was to determine which of these solutions provided the best base line support of βt3 cells and to screen potential additives to the solutions for the ability to improve cell survival during and after hypothermic storage. It is anticipated that these studies will translate to large mammal pancreas models and eventually human pancreas preservation.
Methods
Cell line
A murine cell line, βt3, derived from pancreatic islets was used in these studies. 12 The cells were maintained at 5% carbon dioxide at 37°C and cultured in Dulbecco's Modified Eagles Medium (DMEM) supplemented with 15% horse serum, 4.5 g/l glucose, 2.5% fetal bovine serum, 100 U penicillin, 100 μg streptomycin, and 2 mM glutamine. Cells, 2×104/well, were plated in 96-well microtiter plates and cultured overnight in supplemented DMEM prior to use in experiments.
Storage conditions
After plating the day before, the cells were washed with phosphate-buffered saline (PBS) and then stored in 100 μl/well of either the intracellular formulation of Unisol10,11 or Belzer's Machine Perfusion (BMPS) solution 13 at 4°C for 16–72 h. After cold storage, the cells were washed 3 times with PBS and then incubated in supplemented DMEM for 1 h at 37°C for recovery prior to cell viability and proliferation assessment. 14 Q-VD-OPH (QVD) (MP Biomedical) and α–tocopherol (αT) (Sigma-Aldrich) were added at various concentrations either to the storage solutions during the cold storage period, during a 1 h recovery period in supplemented DMEM at 37°C, or during both periods.
Measurement of cell viability and proliferation
AlamarBlue™ (Trek Diagnostics) was used to assess cell viability by measuring the oxidation/reduction reactions that take place within cells. AlamarBlue was added directly to the plates containing cultured cells in culture medium and incubated for 3 h at 37°C. Upon reduction, alamarBlue changes color, which can be measured and quantified. The culture plates were read using a Gemini EM fluorescent microplate reader (Molecular Dynamics) at an excitation wavelength of 544 nm and an emission wavelength of 590 nm.
In addition to measuring viability immediately after rewarming, the ability of the cells to proliferate was also examined using alamarBlue. Because alamarBlue is nontoxic, it can be used repeatedly without harming the cells. The cells were incubated in alamarBlue for 3 h at 37°C each day for 3–5 consecutive days after hypothermic exposure. Once the plate had been read, the alamarBlue was removed and the cells incubated overnight in regular cell culture medium until the next day. Increased metabolic activity is indicative of proliferation. The values obtained from stored cells were compared with those obtained from untreated control cells.
Apoptosis assays
Using the Guava cell analysis system (Guava PCA-96), three separate assays examining early and late apoptotic events were done. Nexin staining binds to phosphatidylserine that translocates to the outside of the membrane early in apoptosis. Caspase activity is measured using a tagged general caspase inhibitor. The inhibitor will bind to activated caspases and the marker allows the relative caspase activity to be measured. Finally, a modified version of the TUNEL assay is used to measure the DNA ladder. As the DNA fragments expose the 3’-hydroxyl ends, terminal deoxynuceotidyl transferase adds Br-dU to the ends, which can subsequently be labeled using tagged antiserum. Once labeled, the cells can be counted and the percent of the apoptotic cells counted can be determined.
Statistics
All experiments were repeated at least three times with 6–12 replicates in each experiment. Statistical differences were assessed by analysis of variance and post tests were done using Dunnett's multiple comparison test. P values<0.05 were regarded as significant.
Results
Base-line storage conditions
Initial experiments were designed to evaluate the long-term storage of βt3 cells in either Unisol or BMPS without the investigational supplements. Cells were incubated 24–72 h at 4°C. Upon removal, the cells were washed with culture medium and allowed to recover for 1 h at 37°C before measurement of metabolic activity. The βt3 cells demonstrated better viability when stored in Unisol compared with BMPS at all hypothermic incubation times tested (Fig. 1). Bt3 cells stored in both solutions demonstrated similar initial viabilities at Day 0 post treatment. However, cells stored in Unisol recovered faster, demonstrating higher levels of proliferation over time (p<0.05).

Cell viability of βt3 cells after storage at 4°C. βt3 cells were stored for the indicated time periods in either Unisol™ or BMPS. Then cell viability was measured every day for 7 days post-storage. Values represent the mean (±SEM) for 24 replicates. Differences in viability were considered significant by one way ANOVA, p<0.05.
Additives
Two compounds were evaluated for their ability to improve cell viability after storage. Q-VD-OPH is a broad spectrum caspase inhibitor 15 and α-tocopherol is an antioxidant and free radical scavenger.16,17 A range of concentrations of each of these additives was added to Unisol and BMPS during storage at 4°C and evaluated for improvements in cell viability.
QVD was added at several concentrations (0–50 μM). Cells were left in storage for 16–24 h at 4°C, then washed and allowed to recover as described above. Addition of QVD improved cell viability at all concentrations tested in both storage solutions. This improvement was significant at the higher concentrations tested in Unisol, 25 μM (p<0.05) and 50 μM (p<0.01). Improvement in cell viability was also observed using QVD in BMPS but it was not considered significant. Inclusion of QVD produced better viability and proliferation post-hypothermic exposure compared with control cells stored in either storage solution alone (Fig. 2), particularly during longer storage periods.

Cell viability after storage with an apoptotic inhibitor. βt3 cells were stored for 16–24 hours at 4°C in BMPS
The effects of α-tocopherol when added to Unisol or BMPS at concentrations of 0–5 mM were assessed during 16–24 h storage of βt3 cells at 4°C. A modified concentration range 0–1 mM α-tocopherol is presented in Figure 3. Concentrations above 1 mM produced a marked decrease in cell viability (data not shown). The presence of α-tocopherol improved cell viability significantly (p<0.05) immediately after storage, regardless of the concentration used. Increased proliferation was also observed during post-hypothermic exposure culture. The presence of α-tocopherol in either solution produced improvement in viability over either solution alone. The best α–tocopherol concentration was 50–100 μM (p<0.001) using both solutions, Unisol or BMPS. Decreased viability was observed at concentrations>250 μM. Similar results were obtained using the water-soluble α-tocopherol analog, Trolox (data not shown).
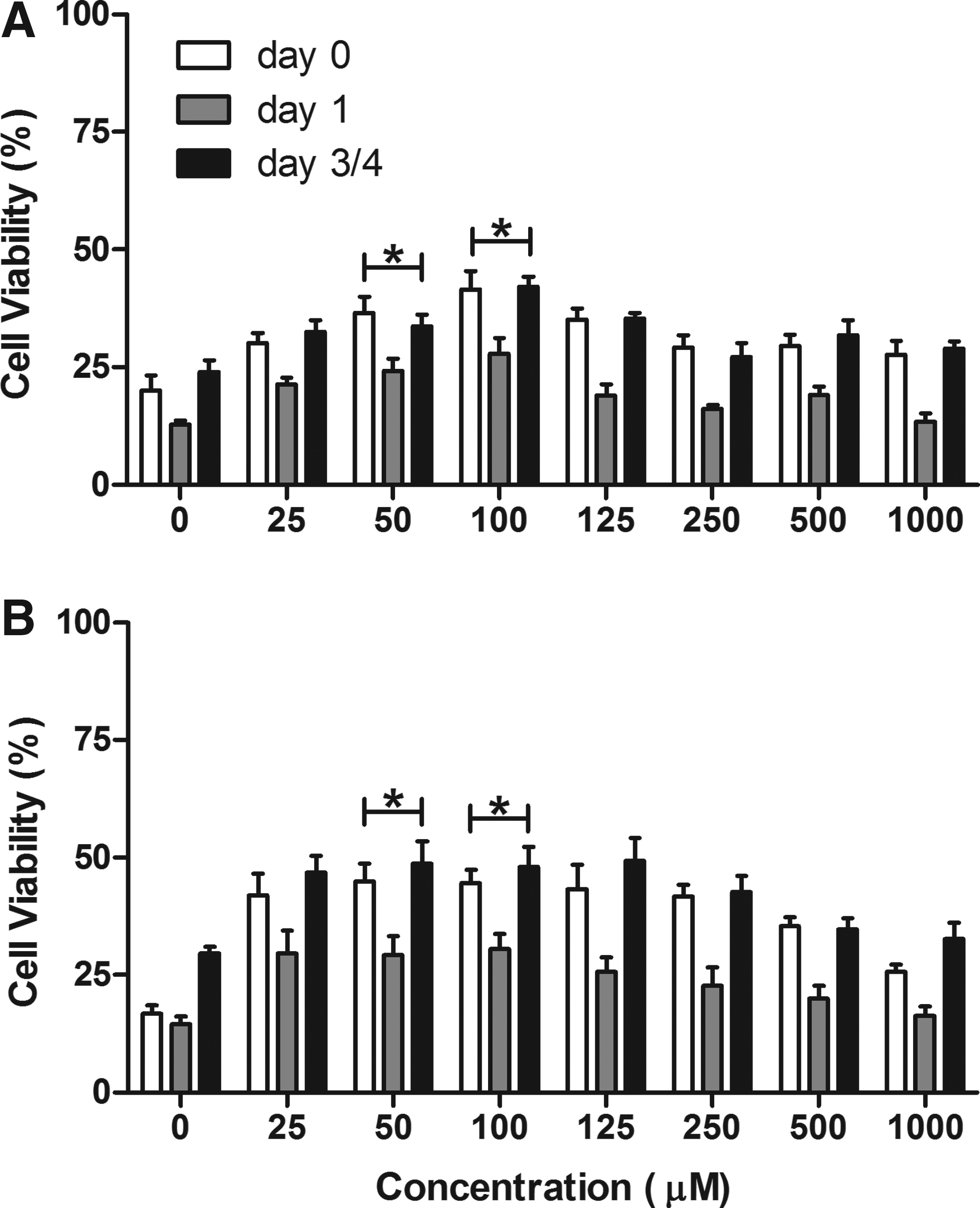
Cell viability after storage with the antioxidant, α-tocopherol (αT). βt3 cells were stored for 16–24 hours at 4°C in BMPS
Combinations of QVD and α-tocopherol
Unisol and BMPS were supplemented with either QVD (10 μM), α-tocopherol (50 μM), both QVD and α-tocopherol, or neither additive. The cells were stored at 4°C for 16–24 h and then viability was assessed immediately after storage and for several days post-storage (Fig. 4). βt3 cells demonstrated benefits when either supplement was included during storage, 26% for QVD and 29% for α-tocopherol. A significant increase in cell viability was observed when a combination of QVD and α-tocopherol was used, (p<0.05) with either Unisol or BMPS. βt3 cells stored in the combination of α-tocopherol and QVD demonstrated higher viability at day 5 post-storage than either additive alone or in only Unisol. Similar results were observed when βt3 cells were stored in BMPS with and without the additives.

Cell viability after storage at 4°C with α-tocopherol and QVD. βt3 cells were stored for 16–24 h at 4°C in BMPS
Optimal concentrations of QVD (10 μM), α-tocopherol (50 μM), or both were then added at different points during the storage experiment. Additives were included either at 4°C during hypothermic storage, during the 37°C recovery phase in supplemented DMEM after hypothermic storage, or during hypothermic storage at 4°C and post-storage recovery at 37°C. The rationale for these experiments was two-fold. First, metabolic activity may continue under hypothermic conditions, albeit at a much reduced rate, and the presence of QVD and α-tocopherol may prevent or slow down any induction of cell death pathways. Second, once cells are returned to physiological conditions, biochemical pathways that may have been inhibited at colder temperatures may no longer be constrained, resulting in reperfusion injury. Thus, the presence of QVD and α-tocopherol may inhibit biochemical pathways that ultimately harm and kill cells under both hypothermic storage and physiological recovery conditions.
The βt3 cells showed no significant preference for where either α-tocopherol or QVD was included. Equivalent viability was observed when individual supplements were used in either hypothermic storage solution or during post-storage recovery (Fig. 5). Higher viabilities were measured when the combination of α-tocopherol and QVD was used, but these were not significant except when cells were stored in BMPS with additives during storage and recovery (p<0.05). There did appear to be some benefit for the presence of α-tocopherol and QVD during hypothermic storage versus at 37°C, suggesting that inclusion here may be more advantageous than during post-storage recovery.

Cell viability after storage and recovery with α-tocopherol and QVD. βt3 cells were stored for 16–24 h at 4°C in BMPS
Since cell survival was lower after storage using BMPS, in the last set of experiments, apoptotic activity was evaluated using this solution. Bt3 cells were stored in BMPS alone, with 50 μM α-tocopherol, 10 μM QVD, or both additives for 16–24 h. Cells were harvested and three apoptosis assays were employed using the Guava cell analysis system. The assays measured an early apoptotic event, translocation of phosphatidylserine to the membrane by Nexin staining, the activity of the caspases that control the apoptotic cascade and the formation of the distinctive DNA laddering (TUNEL) that happens much later in the cascade when the cells are committed to death (Fig. 6). The total number of apoptotic cells was significantly higher in cells treated with BMPS alone versus when either additive or the combination of both additives is used. Caspase activity was significantly decreased (p<0.05) for all the groups tested compared with BMPS alone. DNA laddering was also lower in all groups compared with BMPS alone, but only achieved statistical significance with the addition of α-tocopherol (p<0.05). Nexin staining increased in the presence of the additives, achieving statistical significance when both QVD and α-tocopherol were used in combination. Overall, there was a reduction in apoptosis markers in the presence of QVD, α-tocopherol, or both compared to BMPS alone.

Apoptotic activity after storage with α-tocopherol and QVD. βt3 cells were stored for 16–24 h at 4°C in BMPS alone, with 50 μM α-tocopherol, 10 μM QVD, or a combination of both additives. Apoptotic activity was measured by nexin staining, caspase activity, and the detection of DNA laddering (TUNEL). Apoptotic cells are expressed as the percentage of cells positive for apoptosis markers out of 1000 counted cells. Values represent the mean (±SEM) for 12 replicates. Differences in viability were considered significant by one-way ANOVA.
Discussion
Two hypothermic storage solutions, Unisol and BMPS, were evaluated with and without supplements that may promote cell survival during or after hypothermic exposure. Belzer and colleagues designed UW solution for static storage of organs and subsequently BMPS, a modified version of UW for organ perfusion.13,18–23 The design of Unisol10,11,24,25 was based upon the cellular response to cold literature that has emerged over the decades since BMPS was created.24,26–29 Our goal was development of a base formulation that accounted for both biophysical and minimal biochemical requirements for all applications. The base solution for hypothermic exposure (Unisol) would then be used as a vehicle for additive “cocktails” optimized for the diverse biological requirements of different cells, tissues, and organs. The Unisol design contains components: a) to minimize cell and tissue swelling; b) to maintain appropriate ionic balance; c) to prevent a state of acidosis; d) to remove or prevent the formation of free radicals; and e) to provide substrates for the regeneration of high energy compounds and stimulate recovery upon rewarming and reperfusion. Unisol has been evaluated for the preservation of a variety of cell, tissues, organs, and large mammals. 14,24,30–36 In this study, Unisol maintained βt3 cells better than BMPS in the absence of any supplements. Similar numbers of cells survived 24 h of hypothermic exposure, but the βt3 cell proliferation was significantly better, suggesting that there may have been higher levels of delayed cell death in the BMPS stored group.
Incorporation of specific apoptotic protease inhibitors in preservation media has been reported to improve the survival of a variety of cells and tissues markedly.37–51 With respect to the inhibition of caspases, a variety of compounds have been shown to be effective for mammalian cells in culture. For example, P35 confers irreversible inhibition to a large number of caspases, but we chose here to focus on another popular choice in mammalian tissue culture applications, namely, Z-VAD.fmk (or its latest broad-spectrum counterpart, Q-VD-OPH) since this compound has the ability to inhibit both the intrinsic and extrinsic pathways. 15 Since at this point, we have no clear indication as to which pathway is the most dominant following hypothermic stress, it seemed prudent to investigate compounds that have the potential to intervene in either pathway.
There is an extensive literature on the cytoprotective effects of α-tocopherol (Vitamin E) as a ubiquitous cytoprotective agent acting as both an antioxidant and anti-apoptotic agent. Moreover, the addition of α-tocopherol, or Trolox™, a water-soluble analog, to preservation media has been shown to provide additional protection in a variety of systems during low temperature storage.16,17
The results in this study indicate that supplementation of BMPS or Unisol with either or both the broad spectrum caspase inhibitor, Q-VD-OPH (QVD), and α–tocopherol promoted βt3 cell survival. The supplements were effective both during and after hypothermic exposure. The best results were obtained when both supplements were present throughout the hypothermic exposure and post-exposure recovery periods. These results suggest that apoptosis may play a significant role in βt3 cell death after hypothermic storage at 4°C and return to physiologic culture conditions. Cell death occurs over time post-storage because some cell death may not be immediately manifested upon rewarming. Previous studies have shown that supplementation of preservation solutions with apoptosis inhibitors improves cell survival.37,38,47,49,50 Our results also showed improvement in cell survival post-storage as observed by the measurement of viability immediately after thawing and the increase in cell viability over several days post-storage, indicative of cell proliferation. Oxygen-derived free radicals can also cause tissue damage and the production of free radicals can be enhanced during cold storage.52–54 Consequently, this cell damage may not occur entirely during cold storage but actually occurs after rewarming and reintroduction of oxygen under physiological conditions. Prior studies using antioxidants under hypothermic and normothermic reperfusion conditions to circumvent and/or alleviate damage caused by oxidative stress and reperfusion injury have shown that vitamin E, α-tocopherol, or its water-soluble analog, Trolox, improved survival in a number of cell systems.16,17,55–58 There is also evidence that α-tocopherol may be able to act synergistically with other cytoprotective agents. 16
Analysis of the apoptotic markers representing different stages of the cell death cascade revealed that later stage markers, caspase activity, and DNA laddering, were significantly reduced with the addition of one or both supplements. In contrast, there was an increase in nexin staining. This stain identifies an early apoptotic event, the translocation of phosphatidylserine, before cells have been fully committed to apoptosis. This observation suggests that the βT3 cells may have started down the apoptotic pathway but they did not continue and ultimately more remained viable and proliferated. The addition of QVD or α-tocopherol protected βT3 cells during hypothermic storage so that fewer cells were committed to apoptosis and cell death.
In conclusion, these studies demonstrate that addition of compounds to organ perfusion storage solutions, which target specific cell death pathways, can improve βt3 cell viability. The caspase inhibitor, QVD, and the antioxidant, α–tocopherol, demonstrated statistically significant improvements when included in storage solutions during hypothermic exposure and subsequent recovery in culture medium. Unisol consistently yielded higher βt3 cell viability than BMPS with and without supplementation. The aim of this study was to assess the survival of βt3 cells after hypothermic storage in organ preservation media in the presence or absence of key cytoprotective supplements. The latter were selected to target storage-induced apoptosis and oxidative stress based upon recent developments in the field of cell protection against ischemia, cold storage, and reperfusion injury. The salutary effects of these supplements demonstrating improved survival based upon metabolic activity, apoptosis markers and cell proliferation now calls for additional studies to evaluate the effects of solution supplementation during pancreas storage upon Islets of Langerhans.
Footnotes
Author Disclosure Statement
This work was supported by Grant # DK076326-01 from the National Institute of Diabetes and Digestive and Kidney Diseases of the National Institutes of Health. All the authors are employees of and Kelvin G.M. Brockbank is also an owner of Cell & Tissue System, Inc.