
Abstract
Human tissue banks are a potential source of cellular material for the nascent cell-based therapy industry; umbilical cord (UC) tissue is increasingly privately banked in such facilities as a source of mesenchymal stem cells for future therapeutic use. However, early handling of UC tissue is relatively uncontrolled due to the clinical demands of the birth environment and subsequent transport logistics. It is therefore necessary to develop extraction methods that are robust to real-world operating conditions, rather than idealized operation. Cell yield, growth, and differentiation potential of UC tissue extracted cells was analyzed from tissue processed by explant and enzymatic digestion. Variability of cell yield extracted with the digestion method was significantly greater than with the explant method. This was primarily due to location within the cord tissue (higher yield from placental end) and time delay before tissue processing (substantially reduced yield with time). In contrast, extraction of cells by explant culture was more robust to these processing variables. All cells isolated showed comparable proliferative and differentiation functionality. In conclusion, given the challenge of tightly controlled operating conditions associated with isolation and shipping of UC tissue to banking facilities, explant extraction of cells offers a more robust and lower-variability extraction method than enzymatic digestion.
Introduction
Mesenchymal stem cells (MSCs) are a type of adult stem cell found in many organs and tissues in the body.1,2 They reside in a specific area of each tissue, the stem cell niche, where they retain stem cell characteristics of self-renewal and differentiation and are involved in the maintenance and repair of tissues and organs throughout the life span of the individual. 3 The multiple potential therapeutic applications of these cells, such as immunomodulation, cartilage, bone or muscle repair, amongst others, have led to a substantial number of MSC-based therapeutic developments. They are therefore a prime candidate-cell type for individual donor banking for future use. The opportunity for personal banking has been enhanced by the identification of MSCs in human umbilical cord (UC) tissue, 4 a tissue previously disposed of as waste at birth, which affords easy and ethically non-contentious access to the cells.5,6 Further, evidence of a small population of cells with wider potency has increased the potential value of UC storage. 7
MSCs have been isolated from different areas of the UC, including blood, umbilical vein sub-endothelium, and Wharton's jelly.8,9 However, it is still inconclusive whether MSCs isolated from these different compartments represent functionally different populations. 6 The primary method of MSC extraction involves nonselective isolation by enzymatic digestion, generally using Type I or Type A collagenase-based solutions. These solutions often have poorly defined and potentially detrimental caseinase, clostripain, and tryptic activities. Recent modifications describe the use of a combination of collagenase with hyaluronidase which facilitates the degradation of matrix ground substance and shortens the time required for the isolation process.10,11 The use of type II collagenase, which has stronger clostripain activity, or collagenase type B, which is relatively more efficient at solubilizing the UC microfibrils has also been successful. 12 The duration of collagenase treatment is critically important, especially if collagenase/hyaluronidase cocktails are used, since there is a risk of degradation of cellular external lamina, a phenomenon preventing cells from adhering to the culture substrate after isolation and potentially causing cellular damage. 8 The time required for tissue digestion ranges from 30 minutes 13 to 16 hours 14 depending on the quantity/concentration of enzyme and duration of treatment with digesting reagents. Additional process steps, such as filtration of the digested material through 70–100 μm pore-sized cell strainers, are sometimes included to facilitate the removal of any unwanted tissue debris.13,15
An alternative, less explored, cell extraction method is explant culture. 16 This entails fine chopping of the Wharton's jelly section of the cord tissue, after excision of the blood vessels, and plating of the fine fragments in sterile culture plates or Petri dishes.7,17
In order to store tissue with consistent clinical potential, methods need to be selected to minimize the variability in the extracted stem cells given the operating restrictions of the cell banking model. Due to the nature of tissue collection in a birthing environment the early period of tissue processing is relatively uncontrolled; the priority is maternal and neonate safety. Further, tissue often needs to be transported from maternity units to distant processing sites. Such factors make imposing tight process controls on early handling challenging. In addition, innate biological variation in the tissue will affect the cell yield.
Therefore, cell isolation methods should not be compared under idealized conditions. Methods should be assessed and engineered for robustness to innate biological tissue variation or arising variation due to tissue collection procedures. This is particularly important for tissue stored for autologous use, where a processing facility will not be able to select tissue based on favorable characteristics. Once a method has been established, determining the statistical distribution inherent in the method relative to required cell yield (process capability) will be necessary to allow tissue processing facilities and banks to predict the risk of sub-optimal cell yield from a given cord tissue section and processing method, and thereby to determine the tissue storage requirements and isolation methods with acceptable risk of adequate cell recovery.
We have analyzed the two primary methods of cell extraction, enzymatic digestion and explant culture, to identify the relative variability in cell recovery. We have identified the robustness of each method to changes in ‘hard to control’ process variables to define methods that would be more appropriate to maintain quality under different operational restrictions.
Methods
Umbilical cord collection and transportation
All cords were collected within 5 hours from birth. Using two fingers, blood was pressed out of the cord to neonatal direction. The cord was then cut from the placenta with a sterile scalpel and the outer surface wiped with an alcohol wipe (Cole-Parmer Instrument Co. Ltd., UK). The cord was placed in a sealed sterile flask containing Dulbecco's Phosphate Buffered Saline (D-PBS, Life Technologies, UK) with 1% Antibiotic - Antimycotic (100x Penicillin/Streptomycin/Amphotericin – PSA, Life Technologies, UK) and then shipped via courier to the lab facility in secure sealed boxes.
Tissue preparation
Twenty sections of the cord were cut, from neonatal to placental end, the former referenced as section 1 and the placental end as section 20. Sections were washed with fresh 1% PSA in D-PBS, and then 200–400 mg slices were cut from each section, and placed into 24 well plates (Nunc, Scientific Laboratory Supplies, UK) with sterile 1% PSA in D-PBS, to re-wash, ready for enzymatic or explant processing. Remaining tissue sections with time-delayed processing were placed in sealed 50 ml centrifuge tubes (Fisher Scientific, UK) and stored at ambient temperature.
Isolation by enzymatic digestion
200–400 mg slices were chopped to fine fragments of approximately 1-2 mm3 and then placed in a 15 mL centrifuge tube (Fisher Scientific, UK) containing 3mL of 0.075% Collagenase type I solution (AMS Biotechnology Ltd, UK) in warm Dulbecco's Modified Eagle Medium Low-Glucose with GlutaMAX™ (D-MEM LG - 1X, Life Technologies, UK) and 1% PSA. Cord slice fragments were enzymatically digested for 18 h at 37°C inside an incubated shaker (Mini 4450, ThermoFisher Scientific, UK). The contents of each tube containing digested cord fragments was then diluted with 5 mL of warm D-MEM LG containing GlutaMAX™ with 20% fetal bovine serum (pre-screened FBS for MSCs, Fisher Scientific UK, country of origin USA). The diluted digest solution was added to T25 culture flasks (BD flasks, Scientific Laboratory Supplies, UK) and incubated at 37°C and 5% CO2. Media change was at 48 h and every 3 days thereafter. The process was repeated with stored cord tissue sections after 72 h and 120 h. After 7 days in culture cells were passaged with TrypLE™ Express (1X, Life Technologies, UK) and counted to assess yield (ViaCount Assay on a Guava® System, Merck Millipore, UK).
Isolation by explant culture
200–400 mg cord slices were individually positioned in the center of a dry 6 well plate (Nunc, Scientific Laboratory Supplies, UK) and placed in an incubator for 30–40 min, at 37°C and 5% CO2. 2 mL of media per well was then added without disturbing the tissue by slow dispensing at the side of the well. Medium consisted of D-MEM LG containing GlutaMAX™, 1% PSA and 20% FBS. Media were changed every 3 days. The process was repeated with remaining cord tissue sections after 72 h and 120 h of storage. At 14 days in culture the cell yield was assessed by performing cell counts as previously described.
Phenotype and functionality
Cells were tested for expression of the following markers: CD90 (Human CD90/Thy1 Phycoerythrin MAb, R&D Systems, UK), CD105 (Human Endoglin/CD105 Phycoerythrin MAb, R&D Systems, UK), CD34 (FITC/581/Human/RUO, BD Biosciences, UK), CD45 (Phycoerythrin MAb, R&D Systems, UK). Flow cytometry analysis was performed on a Guava® System (Merck Millipore, UK). Cells were differentiated towards three standard mesenchymal lineages, adipogenic, chondrogenic and osteogenic. Cells were differentiated with the use of StemPro® Adipogenesis, Chondrogenesis and Osteogenesis Differentiation Kits (R&D Systems, UK) according to manufacturer's instructions. Differentiation was evaluated at 24 days in culture with HCS LipidTOX™ Neutral Lipid Stain Green stain solution (Life Technologies, UK) for adipogenesis, with Alcian Blue stain (Sigma-Aldrich, UK) for chondrogenesis and with Alizarin Red S stain (Sigma-Aldrich, UK) for osteogenesis, according to manufacturer's instructions. Statistical analysis – Data were analyzed using Minitab 15™. Statistical analysis was applied to Box-cox transformed (log10 or square root) data. Summary statistics were transformed back to original scale for presentation. Data were analyzed graphically using box and interval plots to compare means and variability. Outliers in the data sets were not excluded unless attributed to a special cause. Interaction plots were used to show association between factors. Two-sample hypothesis tests were applied to compare mean and standard deviation of the responses between the two isolation methods. A general linear model (GLM) analysis was used to perform an analysis of variance (ANOVA) for the response variable (cell yield) in balanced and unbalanced data sets involving fixed (isolation method, storage time) and random (cord) factors. Post-hoc Bonferroni Simultaneous test was used for comparison of multiple means. For all tests, p≤0.05 was considered significant. One cord (8) was excluded from the analysis because no cells were isolated.
Results and Discussion
Twelve umbilical cords were cut into 20 equal sections. Slices were taken from each section for processing after storage of the cords for 24, 72, and 120 h (to represent variable processing delays associated with logistics of transport and delivery). Slices were then processed by enzymatic digestion or explant culture method. Cell yield data were analyzed for each method to determine the inter- and intra-cord variation and effect of storage time.
Effect of isolation method on cell yield
Baseline process performance was established by evaluating the cell yield from cord slices processed by explant (n=330) or digestion (n=660) method, within a standard process operating range. Digestion yield was assessed after 7 days and explant after 14 days because of significantly slower cell release. The average cell yield isolated from cords using the digestion method was higher (2.3×105) compared to the explant method (1.8×105) (p=0.002), but the cell yields were significantly more variable (p=0.0005) (Table 1).
This suggests that cell yield from the explant method, while slower, may be more robust to input variation (i.e., delay before processing or biological variation) in the operating range selected. An ANOVA (GLM) was conducted to decompose the variation in the cell yield amongst the factors (method, cord and storage time). The analysis showed that cord random effects contributed a significant amount of variation to the observed cell yield. Significant interactions were observed among random (cord) and fixed (method, storage time) factors with evidence that cell yield from cord is dependent on isolation method (p=0.045) and cord storage time (p=0.024), but also that the effect of cord age was dependent on isolation method (p=0.0005) (Fig. 1). To determine how the isolation method influenced the cell yield from the cords at different storage times (24, 72 and 120 h) the data were further stratified and an ANOVA (GLM) applied independently to each isolation method.
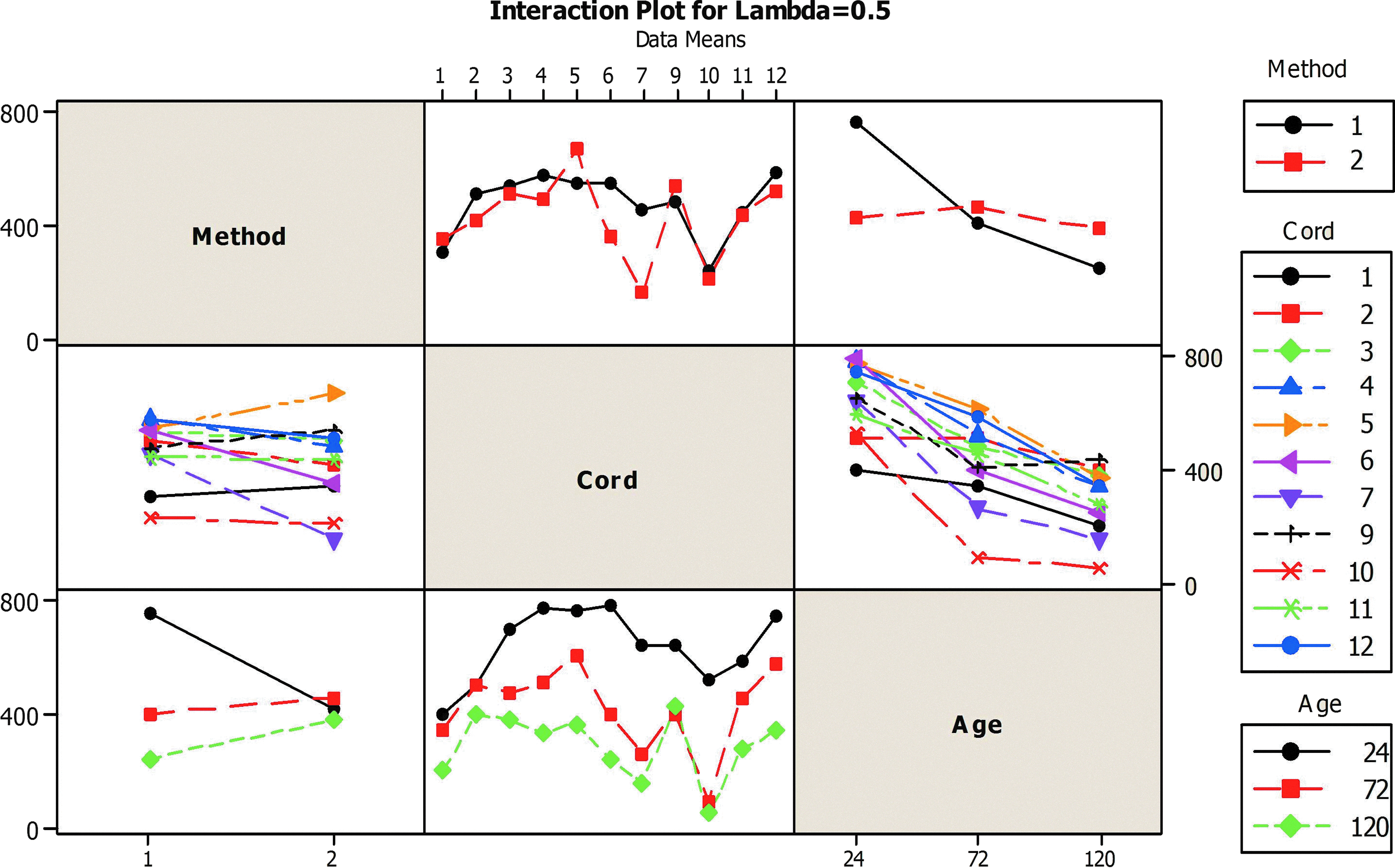
Interaction plot for normalized data (lambda=0.5, sqrt transformation) for digestion (1) and explant (2) isolation methods. Interactions are observed between random factor (cord) and fixed factors (age and isolation methods) as determined by a GLM ANOVA. A Color version of this figure is available in the online article at www.liebertpub.com/bio
Effect of cord storage time on cell yield
The average cell yield isolated from cords using the enzymatic digestion method differed significantly between the storage time points (p=0.0005); the effect, however, was variable, between individual cords (Fig. 2). Post hoc tests revealed that storage of cord significantly reduced mean cell yield after 72 h (1.7×105) and 120 h (0.64×105) compared to the cell yield at 24 h (5.9×105). In contrast, the mean cell yield from the Explant isolation method did not differ significantly between the storage time points (p=0.08), with some suggestion of yield improvement over the first 72 h, although effects could again be seen to vary between individual cords (Fig. 2).

The effect of storage time on cell yield from umbilical cord tissue processed by explant or digestion methods. Boxplots show interquartile range shaded, with median line, whiskers represent 1.5 times the interquartile range, and stars show extreme values beyond this range.
This analysis indicates that cord storage time causes significant reduction in mean cell yield (in a population of cords) when cells are isolated using a digestion method but not when isolated using the explant method. It is hypothesized that this could be due to cells becoming more susceptible to stress of single cell isolation after prolonged storage, rendering older cord cells more susceptible to damage during digestion relative to explant. It could also be associated with cell-mediated extracellular matrix degradation during storage, allowing easier migration of cells out of the tissue during explant culture, but exposing them to more enzyme activity during digestion. An alternative explanation is that the explant culture limits the number of cells that can migrate from the cord; in this scenario the reduced cell content with storage time is still present in the explant cultures, but not apparent due to the limitations of the method in releasing the cells. If the latter were the case we would expect to see reduced growth over time; this does not occur and is discussed below.
The variation of cell yield (%CV) increases for both methods with increased tissue storage time (24 h=31%, 72 h=42%, 120 h=52% explant; 24 h=37%, 72 h=67%, 120 h=76% digestion). This strongly implies a non-uniform degradation of cell yield potential from fresh tissue. A non-uniform degradation of tissue yield suggests a change in the relative performance of cords over storage time. This is apparent in Figure 3, which shows that the relative difference between mean cord cell yields from digestion reduces with increased storage time. A weak inverse correlation (R2=55%) between higher initial cell yield and the cells recovered after 72 h of tissue storage as a proportion of initial yield, suggests that this non-uniform degradation may be due to higher yielding tissue degrading faster. It is possible that more metabolically active tissue, with higher cell content, is more sensitive to storage. An alternative explanation is that the cell population in the cords is heterogeneous in sensitivity to a processing delay—this is supported by the fact that approximately 50% of potential cell yield is lost with each additional 48 h of storage/delayed processing. This implies more rapid cell loss in absolute numbers in the early period of storage, and would lead to smaller relative differences between cords after prolonged storage.

Interval plots showing the mean yield and 95% confidence interval of the mean stratified by individual cord after different storage times.
Cord to cord variability and the influence of sampling location on cell yield
In order to determine if the sampling location (section) within the cord influences the cell yield, the mean yield from each cord section was plotted against the sequential position from neonatal to placental end of the cord (section 1=neonate end and section 20=placental end), for both isolation methods (Fig. 4). A discernible effect of tissue location on mean cell yield was not observed for the explant method, but a trend of increasing mean cell yield at the placental end of the cord was observed when the isolation was conducted by enzymatic digestion. This implies that the explant method may initially restrict the number, or growth, of cells recovered from the tissue, thereby potentially hiding a difference in tissue cell content. This effect was also not seen in all cords.
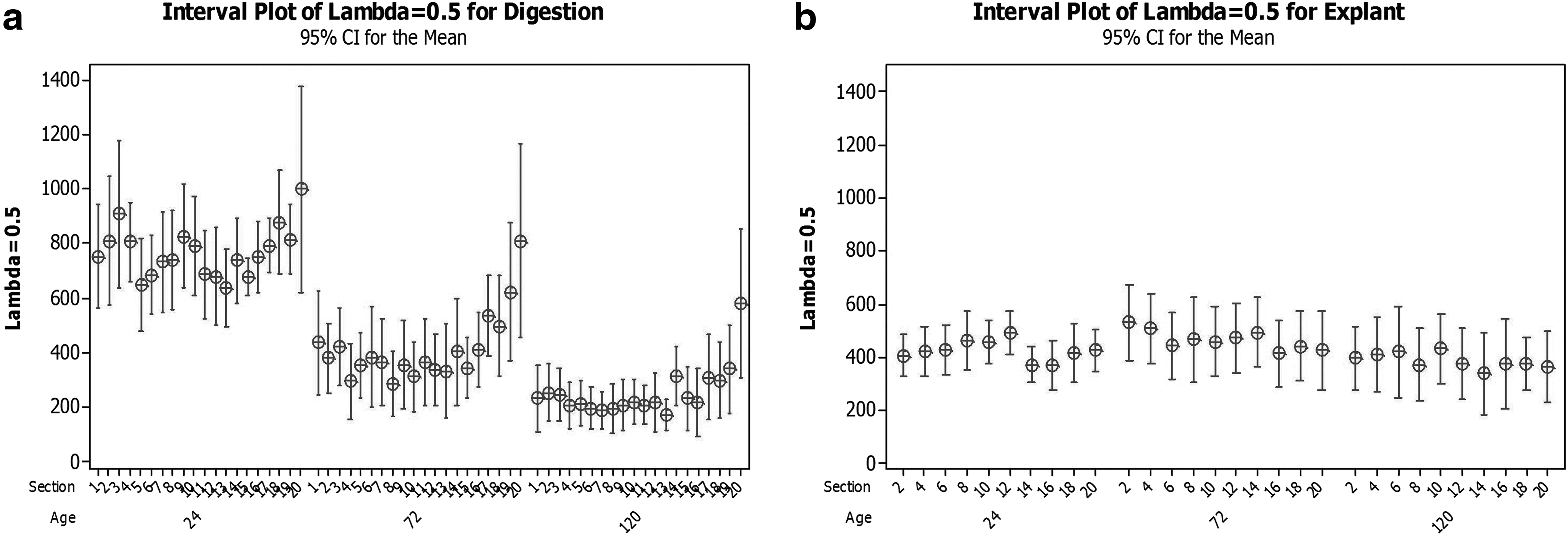
The effect of location in cord on cell yield from umbilical cord tissue processed by explant or digestion methods (section 1=neonate end and section 20=placental end). The 95% statistical confidence interval for the population mean is shown for each position after each storage time.
Verification of cell quality
The quality of cells extracted via the different isolation methods was assessed to verify their functional capacity. Cells from selected cord slices processed by enzymatic digestion or explant method after variable storage times were cultured over a prolonged period of 16 passages and tested for their capacity to differentiate to adipogenic, osteogenic and chondrogenic lineages at passage 1, 8, and 16. Similar to reports by others,18,19 a qualitative reduction in differentiation potential from P1 to P8 was observed as illustrated by representative histochemistry micrographs (Fig. 5). At P16 the cells no longer differentiated effectively. However, no discernible qualitative differences in differentiation outcomes were observed between isolation methods or storage time points. Similarly, the surface marker profile (CD90, CD105) showed some reduction over time in culture, but there was no significant effect of cell isolation method or storage. The proliferative rate (within the time period tested) of the cells isolated by the two methods also did not differ nor was it affected by cord storage time. A small percentage (2.5%) of the individual tissue slice cultures stopped proliferating before P8; all of these culture failures were isolated by digestion method, further supporting explant as a more robust extraction method. Overall the evidence suggests that the explant method is a slow early release, rather than a significantly restricted release, isolation method. This is an important distinction. If the explant was releasing a very small proportion of potential cells, we would anticipate a lower proliferative capacity and more rapid reduction in differentiation potential compared to digestion.
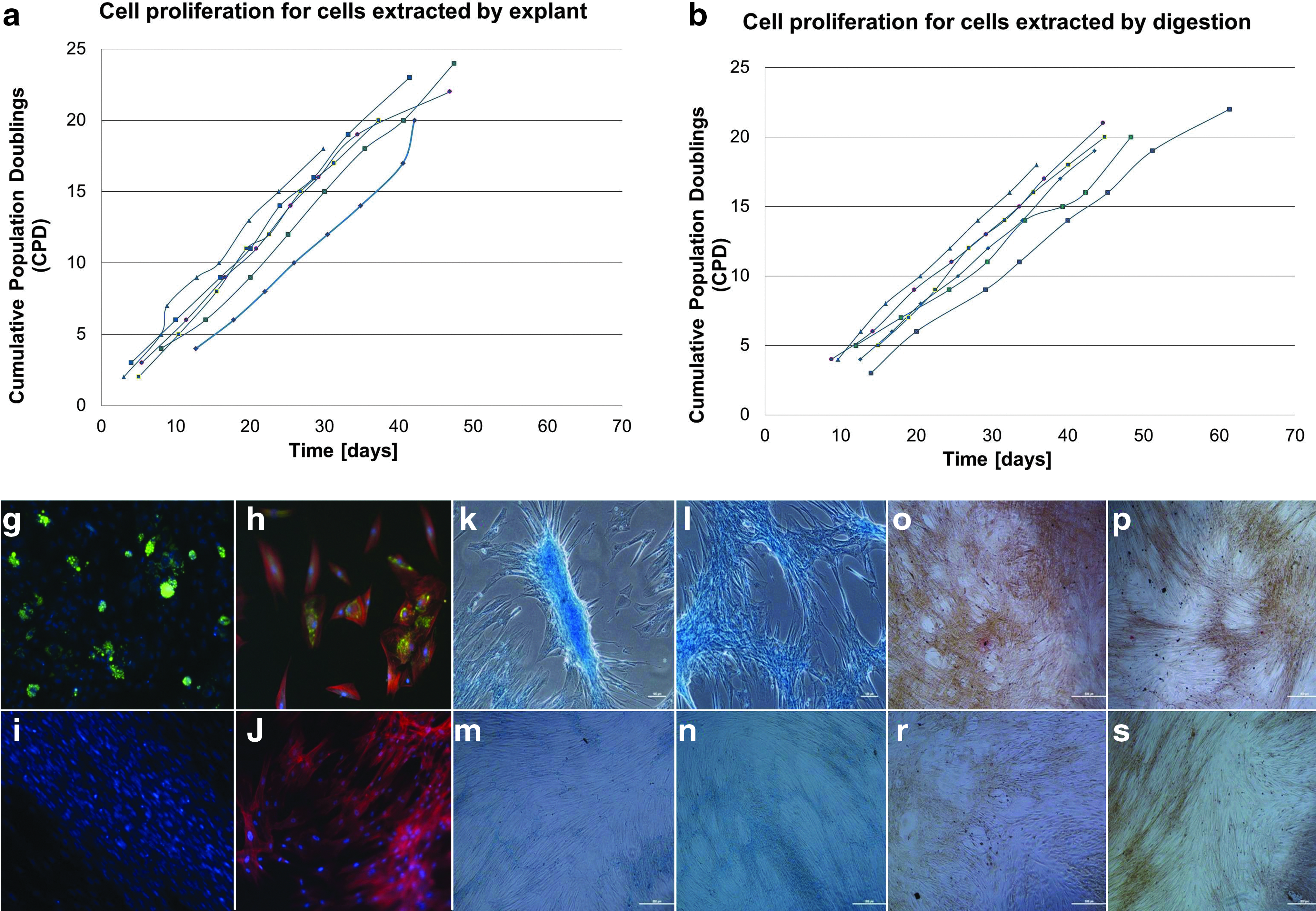
Growth and differentiation properties of cells extracted from two cords by explant or digestion after different storage times.
Operational significance
Given an operating environment where control of rapid tissue processing and cord location selection is not feasible the explant method offers logistical and quality benefits over the digestion method. This applies to many birth environments and subsequent banking and transport logistics.
The impact of the observed variation on operational performance can be illustrated by comparing the predicted population distributions for cell yield obtained from both isolation methods. Figure 6 shows that despite the lower average cell yield, the explant isolation method is more robust than the digestion method to the effects of cord storage on cell yield. This “capability snapshot” indicates that if the lower limit for acceptable cell yield was set at 50,000 cells (as shown in Fig. 6), fewer cords would fail to provide sufficient cells if they were processed using the explant method (14% out of specification) compared to the digestion method (27% out of specification).

Population distribution plots of cell yield for enzymatic
Conclusions
Biological variation in patients, or biological material introduced into samples due to isolation and handling will have a major effect on the safety and efficacy of clinical application. It is critical that processes are engineered for robustness, and with an understanding of critical sensitivities, rather than simply for optimal yield under highly controlled conditions. It is necessary to map the operating environment and assess risk factors before empirically determining the effect on the process. This will be particularly critical for processes using primary tissue or cell sources where the biological variation at input is likely to be high; it will also be labor intensive, requiring large data sets, such as those presented here, due to inherently high variability.
The study indicates that the cell yield obtained from an explant method, while lower in a given timeframe, is more robust to common process/biological input variables relative to the digestion method. Verification of expansion capacity and differentiation potential indicates no loss of potential due to this slower initial yield. The data are important as they indicate that careful design of processing can reduce problems associated with variable biological input material. Regulated therapeutic products will require characterized and risk-assessed manufacturing processes. This fits the philosophy of quality by design (QbD) 20 ; an approach to understanding process operating space and risks of associated variables. The type of study conducted here is an example of the type of data and analysis that will be required to inform a QbD type approach for cell product development and manufacture. It will help tissue processing facilities and banks to predict the probability of cell yields from tissue section given different operating ranges, and inform the experimental approach of others.