
Abstract
Despite marked developments in the field of cryopreservation of cells and tissues for research and therapeutic applications, post-thaw cell death remains a significant drawback faced by cryobiologists. Post cryopreservation apoptosis and necrosis are normally observed within 6 to 24 h after post-thaw culture. As a result, massive loss of cell viability and cellular function occur due to cryopreservation. However, in this new generation of cryopreservation science, scientists in this field are focusing on incorporation of apoptosis and necrosis inhibitors (zVAD-fmk, p38 MAPK inhibitor, ROCK inhibitor, etc.) to cryopreservation and post-thaw culture media. These inhibitors target and inhibit various proteins such as caspases, proteases, and kinases, involved in the cell death cascade, resulting in reduced intensity of apoptosis and necrosis in the cryopreserved cells and tissues, increased cell viability, and maintenance of cellular function; thus improved overall cryopreservation efficiency is achieved. The present article provides an overview of various cell death pathways, molecules mediating cryopreservation-induced apoptosis and the potential of certain molecules in targeting cryopreservation-induced delayed-onset cell death.
Introduction
C
During the development of preservation protocols and preservation media, scientists have focused on overcoming cryopreservation-induced cell death that occurs mainly because of intracellular ice formation and chemo-osmotic stress leading to morphological and functional damage of cells and subsequent necrosis. To overcome this difficulty, many investigators have used cryoprotective agents such as Me2SO and glycerol to preserve biologics in an extracellular-like carrier solution (i.e., standard cell culture medium).1–5 However, a low survival rate of cryopreserved cells is still observed in spite of using cryoprotectants such as 10% Me2SO. 6 In this context, a molecular-based mode of cell death following cell preservation was first reported by Nagle et al. (1990) under nonfreezing conditions. This was further studied by Baust et al. (1998) under freezing conditions.7,8 Later on, many scientists reported cryopreservation-induced cell death and provided efforts to modify the cryopreservation solution accordingly. Baust et al. 8 used Hypothermosol® (HTS) as an intracellular-like hypothermic maintenance solution and modified it by adding 5% Me2SO and caspase I inhibitor V to HTS for controlling cryopreservation-induced cell death and traumatic necrosis. 8 The modified HTS is known as CryoStor® CS 5N. In their study, the addition of a caspase inhibitor showed remarkable improvement in cryopreservation outcome compared to that of the parent solution without inhibitor. This review spotlights the mechanism of apoptosis and necrosis that occurs during cryopreservation of cells and tissues, and various potential molecules that target cryopreservation-induced delayed onset cell death (CIDOCD), thereby improving cryopreservation.
Modes of Cell Death
Multicellular organisms eliminate unwanted, damaged, or dying cells by two main mechanisms: necrosis and apoptosis. Necrosis, or pathological cell death, is an energy independent process that is characterized by cellular and organelle swelling, formation of blebs and vacuoles, disaggregation and detachment of ribosomes, loss of organelle and cell membrane integrity and rupture, random DNA degradation by endonuclease resulting in cell lysis, and activation of active immune and inflammatory response due to release of cytokines.9–15 Necrosis is triggered by multiple stresses such as osmotic shock, hypothermia and cryopreservation, ischemia, ionic disregulation, and exposure to toxic agents. On the other hand, apoptosis or programmed cell death is an energy-dependent process involved in homeostatic maintenance of cell number and tissue size in complex organisms. It is characterized by the activation of intracellular proteases from their inactive zymogens (procaspases to caspases), cellular shrinking, chromatin condensation, nonrandom cleavage of DNA into 180 KDa fragments by exonucleases, maintenance of intact cell membranes, externalization of phosphotidylserine, and formation of membrane blebs (apoptotic bodies). Furthermore, no recruitment of immune and inflammatory responses has been noted. It has also been reported that apoptosis may be initiated by various stresses such as anoxia, nutrient depletion, withdrawal of essential growth factors, radiation, cytotoxic agents, and exposure to extreme temperatures.8,16–23 It is believed in many cases that when a cell commits to cell death, first apoptosis is activated and then continues via the classical apoptotic pathway until stresses experienced by the cell exceed normal limits or the energy reserves become too low for apoptosis to continue. It is at this point the cells switch from an apoptotic to a necrotic pathway of cell death called secondary necrosis.24,25
Apoptosis
Apoptosis may be defined as a physiological cell death that occurs in response to environmental and developmental signals, during which dying cells silently vanish, without traces left behind. In mammals, apoptosis can be triggered mainly by two different pathways (Fig. 1), an extrinsic pathway also known as the death receptor pathway and an intrinsic pathway (mitochondrial pathway). An additional pathway termed the granzyme pathway involves T cell-mediated cytotoxicity and perforin granzyme-dependent killing of cells via either granzyme A or granzyme B. The extrinsic, intrinsic, and granzyme B pathways finally converge to a common execution path of apoptosis that involves proteolytic activation of caspases -3 and/or -7 from their inactive procaspases.26,27 The granzyme A pathway is a caspase-independent cell death pathway that is characterized by single stranded DNA damage. 28
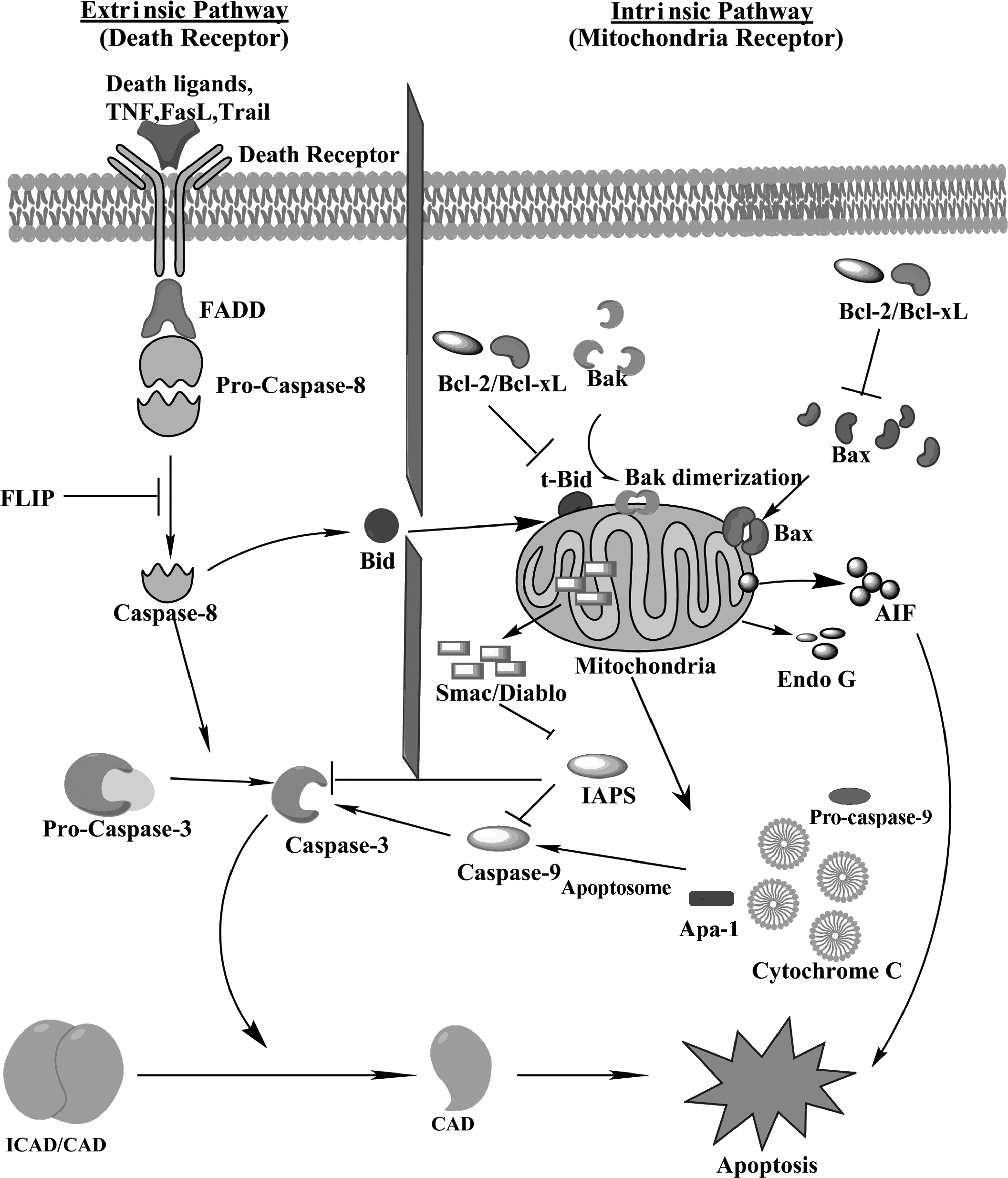
Two distinctive apoptosis pathways have been graphically represented: the death receptor pathway (extrinsic pathway) and the mitochondrial pathway (intrinsic pathway). In the extrinsic pathway in the first pathway death receptors present on the cell membrane ligate due to the binding of their ligands, resulting in the recruitment of adaptor protein Fas-associated protein with death domain (FADD) and procaspase 8. The complex is an “apoptosome” in which the aggregated procaspase transactivates. The active caspase 8 then acts to cleave and activate the downstream caspases. Caspase 8 also activates Bid, which further represents the main link between extrinsic and intrinsic pathway. Truncated Bid (tBid) favors aggregation and permeabilization of Bax and Bak to the outer membrane of mitochondria. In the second pathway, various forms of cellular stress cause mitochondrial membrane permeabilization, which triggers mitochondrial release of cytochrome c, which binds to Apaf1, which in turn self-associates and binds procaspase-9, resulting in an apoptosome. Transactivation of the complexed procaspase-9 to active caspase-9 follows, and the caspase then cleaves and activates downstream caspases. Mitochondria also initiate apoptosis in a caspase-independent manner by releasing pro-apoptotic proteins such as Smac/Diablo, Omi/HtrA2 (caspase dependent), AIF, and Endo G.
Caspases play a major role in the final stage of “execution” in apoptosis. The caspases are a highly conserved family of cysteine proteases with specificity for aspartic acid in their substrates. The cleavage of these substrates induces cell death. Caspases are constitutively present in the cytoplasm of most cells in the form of single chain proenzyme (procaspase). These proteins undergo two proteolytic cleavages, the first divides the chain into large and small domains and the second cleaves the N-terminal domain (prodomain), resulting in fully activated and functional caspase. Caspases have been broadly categorized as initiators (caspase-2, -8, -9, -10), effectors or executioners (caspase-3, -6, -7), and inflammatory caspases (caspase-1, -4, -5).29,30 Caspase cascade activation and DNA fragmentation are reported to be the main features of apoptosis. 30 Apoptotic cells have also been reported to exhibit phosphatidyl serine externalization.31–33
Extrinsic and Intrinsic pathways
The receptors for extrinsic pathways located in the plasma membrane are activated by ligands present in extracellular membranes. Some of the well-characterized ligands and corresponding death receptors include FasL/FasR, TNF-α/TNFR1, Apo3L/DR3, Apo2L/DR4, and Apo2L/DR5.34–38 These receptors contain a death domain on their cytosolic side. The binding of FasL to FasR results in the recruitment of adaptor protein FADD (Fas-Associated protein with Death Domain), whereas the attachment of TNF ligand to TNF receptor results in the binding of TRADD (tumor necrosis factor receptor type 1-associated DEATH domain protein) and recruitment of FADD and RIP (receptor interacting protein).39,40 FADD then binds to procaspase-8 via dimerization of their death domain (DD) to DD on the cytoplasmic side of the receptors, 41 resulting in the formation of a death-inducing signaling complex (DISC).35–42 Formation of DISC induces autocatalytic activation of initiator procaspases, 43 which in turn activates downstream effector caspases (caspases-3 or -7) that further carry out the degradation of cellular targets. On the other hand, activated caspase 8 can also cleave BH3 at protein Bid, resulting in the formation of truncated Bid (tBID). tBID then translocates to the mitochondria, changes the permeability of the inner mitochondrial membrane and ultimately induces the release of cytochrome c, leading to the activation of caspases 9 and 3. 44 The extrinsic pathway can be blocked by the expression of c-FLIP (cellular form of FLICE-inhibitory protein), which is physiologically present as a negative caspase-8 that leads to the formation of a signaling nonfunctional DISC.42,45,46
The intrinsic or mitochondrial pathway is triggered by a variety of extracellular and intracellular stresses such as high temperature, oxidative stress (excess of reactive oxygen species), and irradiation. All these stimuli open the mitochondrial permeability transition pore (MPT). This results in the loss of mitochondrial membrane potential, leading to the release of two specialized types of proteins from the intermembrane space into the cytosol. 47 The first group of proteins consists of cytochrome c, serine protease HtrA2/Omi, and Smac/DIABLO.48–51 These proteins further transduce the apoptotic signal via caspase-dependent pathways. Following release into the cytosol, cytochrome c binds to Apaf-1 in conjunction with dATP, leading to the recruitment of pro-caspase-9 to this complex which is termed an apoptosome52–55 containing units of the above molecules. Active caspase-9 further activates caspase-3 and initiates a proteolytic cascade causing cell death. 56 Smac/DIABLO and Omi/HtrA2 activate caspases by neutralizing the inhibitory effects of inhibitors of apoptosis proteins (IAPs).57,58
The second group of proteins released from the intermembrane space of mitochondria include AIF (apoptosis inducing factor). 59 endonuclease G, 60 and CAD (caspase-activated deoxyribonuclease). 61 These proteins are generally released at later stages of apoptosis. AIF and endonuclease G bring about DNA damage and condensation60–62 in a caspase-independent manner. CAD also results in DNA fragmentation and brings about a more advanced level of condensation. 64 Apoptosis also may occur due to p53 protein by its signaling pathway. It has been reported that signals from external stress can cause DNA damage, which can activate p53 protein. 65 High levels of p53 switch on the apoptosis genes and switch off the survival genes. The intrinsic apoptotic pathway is regulated by many p53 regulated genes. These genes contribute to the activation of caspase 9 and ultimately cause apoptosis. 66 p53 protein is normally present in the cytosol at low concentrations that prevent it from entering the nucleus. Various cellular stresses activate p53, leading to apoptosis in either a transcription-dependent or transcription-independent manner. In a transcription-independent manner, p53 acts directly on mitochondria and induces apoptosis.67,68 p53 also enters nuclei and activates various pro-apoptotic genes in response to generation of reactive oxygen species (ROS).
The mitochondrial pathway is basically regulated by two types of proteins, namely pro-apoptotic and anti-apoptotic proteins belonging to the Bcl (B-cell lymphoma 2) family of proteins, 69 which are controlled and regulated by tumor suppressor protein p53. 70 The pro-apoptotic proteins like Bcl-10, Bax, Bak, Bid, Bad, Bim, and Bik insert into the mitochondrial membrane, resulting in release of cytochrome c from the intermembranal space of mitochondria into the cytosol. 71 In contrast, anti-apoptotic Bcl-2 family members, such as Bcl-2, Bcl-x, Bcl-XL, Bcl-XS, Bcl-w, and BAG prevent cytochrome release by binding and inhibiting pro-apoptotic proteins.
Necrosis
Necrosis, also known as pathological cell death, is a term used to define “cellular murder”.9,10 Morphologically, apoptosis and necrosis are very different. However, with the development of techniques that differentiate apoptosis and necrosis, there are many examples where biochemical and morphological characteristics of both apoptosis and necrosis are found in the same cell. This indicates that apoptosis and necrosis are just two extremes of a series of cell death programs that occur in dying cells. Furthermore, apoptosis is considered as a process of cell elimination without disruption of the plasma membrane, while necrosis is the result of rapid efflux of cell constituents into the extracellular space. However, in vitro apoptosis also leads to the disruption of plasma membranes due to absence of phagocytosis.
It is further reported that members of the TNF (tumor necrosis factor) receptor family (TNF, Fas, and TRAIL) not only initiate apoptosis, but also necrosis.72,73 RIP 1 is a death-domain-containing kinase associated with the death receptors that is required for activating necrosis by the death receptor ligands (FasL or TNFα). 72 Another important sensor is massive DNA damage brought about due to excessive ROS production. Excess of DNA damage activates Poly (ADP-ribose) polymerase (PARP), thereby depleting ATP and activating necrosis due to energy depletion.74,75 PARP is a nuclear enzyme activated in response to excessive DNA breaks. Upon activation, it binds to oligomers of ADP-ribose, thus utilizing ATP for synthesis of PARP substrate NAD+. Ca2+ plays an important role as an inducer and secondary messenger involved in receptor-mediated necrosis. 76
Mitochondria play a crucial role in determining cell fate under stress conditions. They are a source of death promoting factors as they can initiate and amplify the caspase-dependent death pathway by effluxing cytochrome c or initiate cell death in a caspase-independent manner by secreting AIF or Endo G. They also produce ROS that may trigger necrosis.77,78 Being the factory for ATP production, mitochondria choose between ATP dependent and independent processes depending upon their availability for bringing about cell death under stress conditions. The anti-apoptotic members of the Bcl-2 family (Bcl-2, Bcl-XL) can inhibit both apoptosis and necrosis.79–82 A protein belonging to the Bcl-2 family named BNIP3 (BCL2/adenovirus E1B 19 kDa protein-interacting protein 3) possesses pronecrotic activity. 83 Stress kinases like JNK (c-Jun N-terminal kinases) and p38 activated by the ligation of death receptors, oxidative stress, and DNA damage have the ability to initiate both apoptotic and necrotic programs. Inhibition and suppression of these stress kinases have resulted in decreased necrotic death in many cells.84,85 On the other hand, stress-induced apoptosis protected by AKT kinase and MAP kinase ERK can also provide protection against necrotic cell death.86–89 This indicates that proapoptotic (JNK and p38) and antiapoptotic (AKT and ERK) kinases function in similar manners in necrosis.
Finally, the terminal step of necrosis is the activation of proteases. Caspases may or may not be involved in the execution stage of necrosis, as it has been observed in many cases that inhibition of caspases during stress conditions may trigger necrosis. Calpains, cathepsins and serine proteases are the main proteases that are involved in necrosis. 90
Autophagy
Besides apoptosis and necrosis, autophagy also has an important role in cell death machinery. Autophagy is a degradative process where cyoplasmic organelles are hydrolyzed to provide metabolic precursors to fulfil the energy demand of cells that are under stress conditions such as starvation. Though, initially, it was believed that autophagy plays an important role in cell survival by elimination of damaged organelles, recently it has been reported that autophagy also kills the cells. 91 This autophagy cell death mechanism is also called type II cell death mechanism. It works as a secondary death mechanism when apoptosis death pathways are not available. Though apoptosis and autophagy have distinct mechanisms of action, both are regulated by one another. 92
LC3B-II protein is the indicator of autophagy. 93 The unprocessed form of LC3 (pro LC3) is proteolytically cleaved by Atg4 protease, resulting in formation of LC3-I (carboxy terminal with exposed glycine). Upon activation of autophagy the LC3-I is conjugated with a phosphatidylethanolamine (PE) moiety in the presence of Atg7, Atg3, Atg12, Atg5, and Atg16L multimers to generate LC3-II. LC3-II is the only well-characterized protein that is specifically localized in the phagophore (double membrane which separates components during macro-autophagy) to lysosomal degradation. 93
Cryopreservation-Induced Cell Death
The various stress factors encountered by a cell during hypothermic or freezing conditions are metabolic uncoupling/coupling, ionic imbalances, energy deprivation, protease (caspase) activation, membrane phase transitions and alterations, free radical production and accumulation, cellular acidosis, cytoskeleton disassembly, protein denaturation, water solidification, cell volume excursions, hyperosmolarity, and protein denaturation. 94 These stress factors are also known initiators of apoptotic cell death and necrosis.95,96 Post-thaw apoptosis has been reported as significant in a wide variety of cell types, including fibroblasts, hepatocytes, cord blood, peripheral blood derived mononuclear cells, renal cells, spermatozoa, oocytes, and ovarian tissue.16,97–102 These results focus on molecular-based cell death following cryopreservation. Manifestation of cryopreservation-induced cell death may take a few hours to days, and an increase in cell death occurs up to the first 6 hours of a post-thaw period. However, apoptotic and necrotic activity are predominant at 24 hours post thaw, followed by a fall in the percentage of cell death at 48 hours, representing regrowth of the culture. The timing of cell death following cryopreservation termed as Cryopreservation-Induced, Delayed-Onset Cell Death (CIDOCD) has been studied in terms of cell viability 103 and post-thaw caspase activity. 16
Recent studies have shown that apoptotic pathways are activated after thawing, which culminates in the activation of caspases 3, 8, and 9.100,101 As mentioned earlier, apoptosis can be mediated via extrinsic and intrinsic pathways. Previously, it was observed that cryoinjury-induced cell death is initiated by the intrinsic or the mitochondrial apoptotic pathways. However, recently it has been reported that both caspases 8 and 9 are activated through the extrinsic and intrinsic pathways post cryopreservation.102,105 This suggests the possibility of controlling both membrane- and mitochondrial-mediated apoptosis pathways to improve the outcome of cryopreservation procedures.
In response to various stimuli, mammalian cells produce various reactive oxygen species (ROS), including superoxide anion, hydroxyl radical and single oxygen, which play important roles in inducing apoptosis through the mitochondrial pathway. Under normal conditions, the antioxidant machinery in cells regulates the production and removal of ROS. 109 However, under stressful conditions, if ROS exceed the normal level, irreversible cell damage occurs. A correlation has also been established between intracellular ROS levels and apoptosis. 110 Many scientists have reported that cryopreservation results in elevation of ROS species inside cellular cytoplasm.108,111,112 Recently, Xia et al. have reported a rise in ROS species such as superoxide anion and hydrogen peroxide as a result of the procedure of freezing during cryopreservation. 108 Therefore, as stated earlier, it can be concluded that cryopreservation leads to cellular oxidative stress. As superoxide anion induces the release of cytochrome c from the intermembrane of mitochondria into the cytosol, 108 it in turn activates the multiprotein complex, including Apaf-1, cytochrome c, and caspase 9, and thus eventually leads to apoptosis. 50 Therefore, ROS suppression using strong antioxidants can be an effective weapon for improving cell recovery after cryopreservation.
On the other hand, necrosis has been observed to be associated with many cases of cryopreservation failure.8,109–111 However, Heng et al. have demonstrated that the low recovery of cryopreserved human embryonic stem cells (hES cells) is largely caused by apoptosis instead of cellular necrosis. 6 In spite of the reports stating the involvement of apoptosis as responsible for the low viability of cryopreserved cells, the mechanism of apoptosis or the apoptotic pathways involved are still not clear.
New Generation Cryopreservation and Post-Thaw Culture Medium
New generation cryopreservation solutions aim to target cryopreservation-induced delayed-onset cell death (CIDOCD). Targeting and inhibiting various molecules including proteases, signaling molecules, and other proteins bringing about apoptosis and necrosis down the cell death cascades is the prime goal of researchers in this field (Table 1, Fig. 2).
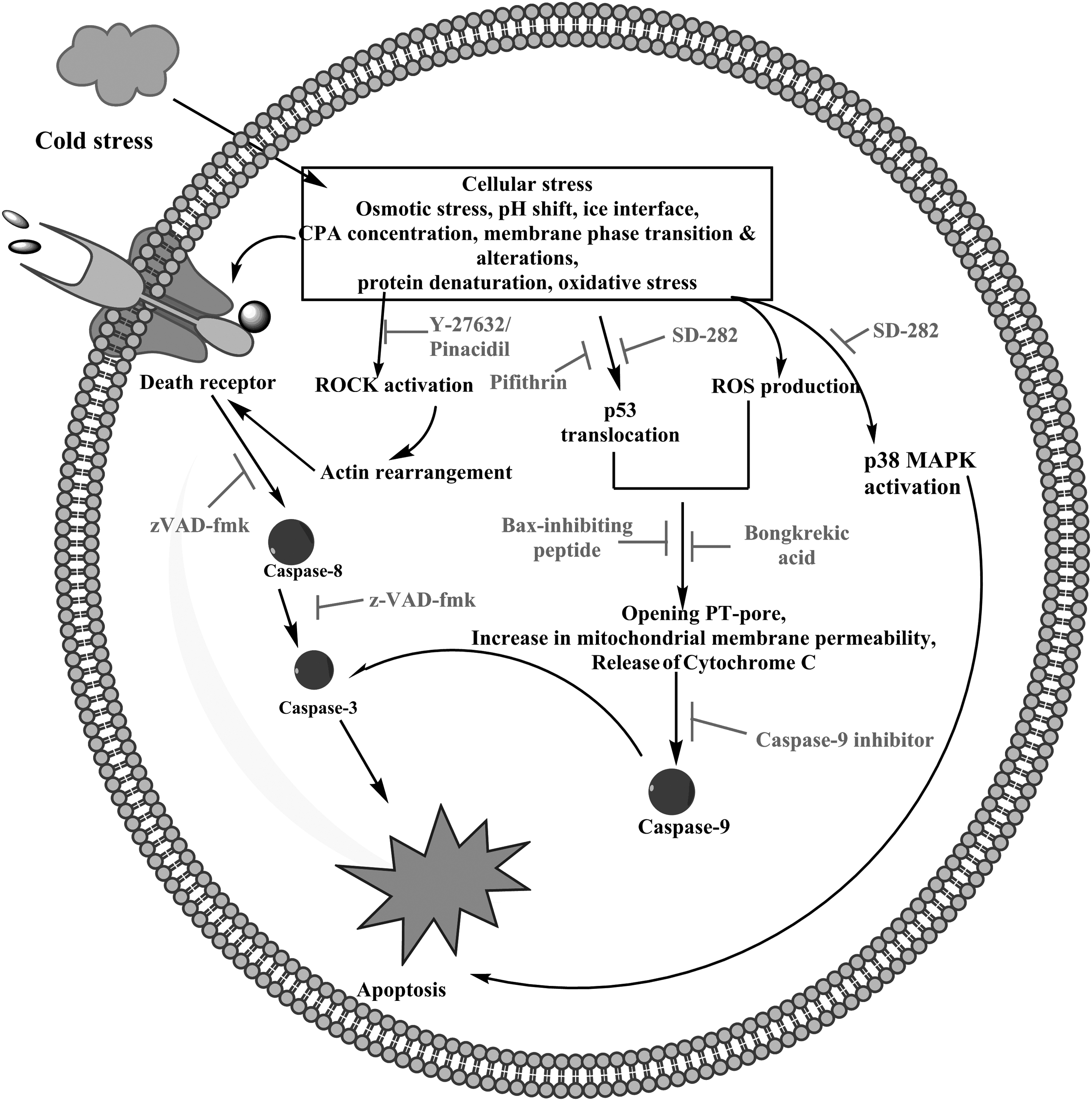
Mediators of apoptosis during cryopreservation and the effective apoptosis inhibitors.
Various caspase inhibitors provide protection against apoptosis.112–114,156 The most widely studied is the synthetic broad-spectrum irreversible inhibitor of caspase enzymes, zVAD-fmk.112,113 It functions as a caspase inhibitor and shows beneficial effects on cell survival during low temperature storage.103,113–115 However, incorporation of only zVAD-fmk in the freezing medium does not improve post-thaw viability remarkably, whereas an increase in cell viability is observed when it is added to post-thaw culture media and incorporated in both freezing and post-thaw culture media.121 In another study, zVAD-fmk not only inhibited caspase activity in cryopreserved cells, but also reduced mitochondrial injury by caspase inhibition through increasing the mitochondrial membrane potential in cryopreserved porcine hepatocytes. 117 As discussed earlier, cryopreservation-induced stress activates a mitochondrial death pathway, resulting in an increase in mitochondrial membrane permeability and reduction in mitochondrial membrane potential. Besides zVAD-fmk, many other potential caspase inhibitors have been reported. Recently Xia et al. studied caspase-9 inhibitor and zVAD-fmk and found that they significantly inhibit caspase-8 and caspase-9 activity without enhancing post-thaw cell viability. 108 This proves that inhibition of caspase activity does not protect the cells. 118 Thus, it may be concluded that caspase 8 and caspase 9 act as downstream effectors in cryopreservation-induced cell death; following inhibition of their activity using specific caspase inhibitors does not improve cell recovery. However, the study of Xia et al. 108 contradicts that of Heng et al. 121 reporting an improvement in cell survival rate of cryopreserved hES cells following the use of zVAD-fmk. Therefore, further study is required to provide a detailed role of caspases in the cryopreservation-induced apoptosis pathway.
Ferrusola et al. 124 reported that the addition of bongkrekic acid (BA), a specific inhibitor of mitochondrial permeability transition pore (PT–pore), during cryopreservation of stallion sperm, has resulted in a significant reduction in caspase activity after thawing, reduction in increase of membrane permeability, and increased mitochondrial membrane potential of thawed sperm cells. 124 It has also been reported that the addition of broad spectrum protease inhibitors to the cryopreservation medium significantly reduces the cleavage of apoptosis proteins such as caspase 3 and 8. 100 The p38 MAPK is one of the MAPK signal transduction pathways that can be activated in response to various environmental stimuli and induces apoptosis. It is activated during freezing-thawing and cryopreservation processes120–122 as well. Inclusion of p38 MAPK inhibitor SD-282 inhibits the phosphorylation of p38 MAPK substrate, HSP27, which reduces cell damage/apoptosis and thus results in improvement in viability of cryopreserved cells. 125
The actin cytoskeleton is reported to play a vital role in various cellular functions like cell motility, shape and regulation of complex signaling systems. 129 It has been observed that alteration of the equilibrium between G-actin and F-actin in response to environmental stimuli can induce apoptosis in mammalian cells. 130 The cytoskeleton may be affected by hypertonic conditions and may bring about remodeling of actin. 131 It is known that during cryopreservation, cells are exposed to anisotonic conditions, which result in elevation of F-actin levels within the cytoplasm. 132 This elevation in F-actin levels itself may be the reason for low recovery after cryopreservation, as alteration in cellular F-actin levels may cause apoptosis. 133 Rho GTPase is reported to regulate cytoskeleton dynamics. Rho GTPase targets Rho-associated kinase I (ROCK-I) protein, which is cleaved by caspases during apoptosis. Activated ROCK-I has been implicated in the regulation of apoptosis134,135 and brings about bleb formation in apoptotic cells, 130 which is one of the essential characteristics of apoptosis. In particular, ROCK triggers a signaling pathway that leads to actin–myosin coupling to the plasma membrane, causing the contraction of actin–myosin and plasma membrane blebbing.136–138 The ROCK pathway can be activated by a variety of external stimuli including osmotic stress. 139 Pizzolla et al. has reported that ROCK leads to phosphorylation of ezrin, resulting in clustering of FAS and, ultimately, apoptosis via the extrinsic pathway. 140 A potent ROCK inhibitor, Y-27632, inhibits apoptosis and is effective in improving the post-thaw recovery and survival of many cryopreserved cells such as human embryonic cells hESCs), induced pluripotent stem cells (iPS cells), human mesenchymal cells, and frozen human embryos141–151 except for cord blood-derived CD34+ hematopoietic progenitor cells. 151 Xia et al. has reported that inhibition of ROCK by Y-27632 following cryopreservation reduces caspase 8 activity. 103 Thus, ROCK is involved in the regulation of activation of caspase 8 during cryopreservation, which may be through ezrin phosphorylation; it thereby induces apoptosis via the extrinsic pathway. However, it is still not clear whether caspase 8 induces apoptosis directly or via the mitochondrial pathway. Thus the mechanism of cryopreservation-induced ROCK pathway and the relation between F-actin and ROCK need to be further studied. Recently, Barbaric et al. found an alternative to the potent ROCK inhibitor Y-27632. They found, through a high-throughput screening assay, an FDA approved drug, pinacidil, with similar properties to those of Y-27632. Like Y-27632, the compound is reported to promote attachment of the post-thawed hESCs, to reduce apoptosis and enhance cell survival when added to the post-thaw culture medium. 152 The advantage of pinacidil over Y-27632 is that it has been used over many years as a vasodilator drug for treating hypertension; therefore it is available and much cheaper than Y-27632, potentially making development of a cryopreservation and post-thaw culture medium economical.
As discussed earlier, in response to various stimuli p53 plays an important role in regulation of apoptosis. 123 At low levels of ROS, p53 acts as a survival factor, whereas at high concentrations of ROS, p53 induces the expression of pro-oxidant genes. 152 Xu et al. reported that Bax-peptide inhibitor promotes cell recovery to some extent and also demonstrated and correlated the high level of intracellular ROS and p53 activation that are caused by the procedure of freezing in cryopreservation. They have also reported a reduction in caspase 9, but not caspase 8, activity in the presence of the p53 inhibitor, pifithrin μ.108,153 This suggests that apoptosis following cryopreservation may be induced by activation and accumulation of p53 in the cytosol through the intrinsic pathway.
Furthermore, there is an increased level of LC3-II (phagophore protein to lysosomal degradation) during storage of stallion spermatozoa. 154 Autophagy is also detected after cold storage of fatty livers. 155 However, further study is essential to understand the exact relationship between autophagy and cell survival due to cryopreservation. The full term of all abbreviations used in this article is presented in Table 2.
Conclusion
Various fields like regenerative medicine, tissue engineering, and bio-banking rely on cryopreservation of cells. The main intention is to maintain the viability and functionality of the cryopreserved cells to be applied in these and other fields. In order to achieve this, scientists are making serious efforts to improve the process of cryopreservation. Traditionally, improvement of cryopreservation has been focused on preserving the structural components of the cells by using penetrating cryoprotectants to mitigate chemo-osmotic and ice rupture-related stresses during cryopreservation. A new generation approach focuses on cell death arising due to cryopreservation protocols. Inhibiting the cell death-associated molecular responses in apoptosis and necrosis has undoubtedly improved the cellular response to cryopreservation.
The cryopreservation process activates various molecules, which in turn trigger an apoptosis pathway. However, the exact mechanism of this apoptotic pathway activated by cryopreservation remains to be studied. Furthermore, the apoptotic pathways activated in one type of cell may not be activated in other cell types as cells may differ in their molecular response to cryopreservation-induced stress. Therefore, a detailed study of the possible pathways and mechanisms leading to apoptosis following cryopreservation procedures is of paramount importance. It is, therefore, suggested that further studies should focus on the screening of other apoptosis pathways and incorporation of necrosis inhibitors in cryopreservation solutions and post-thaw culture media either individually or in combination, to improve the post-thaw survival of cryopreserved cells and tissues without significantly affecting their structure and function.
Footnotes
Acknowledgment
The authors are thankful to the Department of Biotechnology (DBT), Government of India, for providing resources to carry out this work.
Author Disclosure Statement
No competing financial interests exist.