
Abstract
Storage of labile RNA in laboratories is accomplished through ultra-low freezing of the nucleic acids. This however requires expensive freezers, convenient storage, reliable electrical power, and increased shipping costs, thereby making it a less viable option. Biomatrica (San Diego, CA) has created RNAstable®, a stabilization reagent that is used to store RNA in a dehydrated state at room temperature (RT) and protects the RNA from degradation. Our objective was to investigate the sequence integrity and suitability of RNA when stored in RNAstable at extended time periods and at varying temperatures through use of Illumina and Agilent RNA expression microarrays. We observed in Bioanalyzer electropherograms that total RNA extracted from 293 cells stored at RT in RNAstable for 4.5 and 11.5 months is similar in quality to RNA stored at −80°C. Illumina mRNA expression array QC metrics and gene expression patterns from RNAstable-protected RNA, in contrast to RNA stored without RNAstable, correlated well with those of freezer controls. Significantly, when RNA was stored in RNAstable at 45°C for 4.5 months, equivalent to 22 months RT storage, RNA quality, microarray probe signal intensities, probe detection rates, and expression profiles remained similar between RNAstable-protected RNA at RT and the −80°C controls. At 10.5 months, miRNA levels were compared among the storage conditions using miRNA expression arrays. Here too we found strong concordance between miRNA expression patterns when total RNA was stored in RNAstable or at −80°C. Further, Bioanalyzer electrophoresis of RNAstable-protected samples stored at RT for a relative total of 33 months or 50.5 months showed comparable integrity scores to those of −80°C controls. We conclude that use of RNAstable holds promise as an effective stabilization reagent for total RNA and should be useful in situations where shipping and storage options are limited resources.
Introduction
RNA
Several studies have described the effectiveness of commercially available products for protection of RNA prior to use in gene expression assays.1–3 However, these items were generally designed for tissue sample storage prior to nucleic acid extraction, rather than the prevention of degradation of newly isolated RNA. The newest generation of RNA stabilizing reagents, including Gentegra-RNA (Gentegra LLC) and RNAshell (Imagene, Inc), work to preserve extracted RNA at room temperature. Another such reagent, RNAstable® by Biomatrica, is unique in that it was developed based on the principles of anhydrobiosis, a natural biological mechanism utilized by some multicellular organisms that allows their survival in a dehydrated state. 4 Once RNAstable is applied to an aqueous solution of RNA, the sample is desiccated by air-drying or use of a vacuum concentrator. A thermostable barrier is subsequently formed around the RNA, effectively safeguarding RNA integrity for extended time periods. Samples are completely recovered for immediate use by rehydration. RNAstable thus offers an alternative technology for storing RNA at RT, bypassing the needs and concerns associated with cold storage and transport.
Maintenance of RNA integrity is critical in accurately assessing RNA expression in downstream assays. In a previous study, Wan et al. 5 compared skin tissue RNA stored in RNAstable to that of RNA stored frozen at −80°C. After storage for 11 days at RT, yield of RNA and RNA integrity values (RINs) were similar between RNAstable-protected RNA and frozen RNA. Additionally, qPCR assays demonstrated comparable amounts of GAPDH expression between the two storage groups. In a longer term study, purified RNA from HIV-1 was stabilized for 92 days at 45°C as measured by quantitative RT-PCR. 6 Hernandez et al. 7 performed microarray expression analyses on human liver RNA stored in RNAstable at RT or at −80°C for 4 weeks. Microarray quality control metrics showed nearly identical values for RNAstable-protected RNA or frozen controls. Our current study illustrates the suitability of RNAstable-protected RNA for use in microarray expression studies. Here we show data suggesting that RNAstable will preserve total RNA at extended storage times greater than 10-times previously reported for use in both mRNA and miRNA expression arrays.
Materials and Methods
Total RNA Isolation
Human 293 cells were grown to 90% confluence in T-175 flasks in DMEM supplemented with 10% fetal calf serum at 37°C, 5% CO2. Cells were dissociated from the flask by incubation in 0.25% Trypsin-EDTA, cell pellets were collected by centrifugation, and stored at −20°C. Total RNA was extracted from frozen 293 cells using TRIzol® (Life Technologies, Grand Island, NY) following the manufacturer's instructions, and stored at −80°C.
Sample preparation and storage
The chronological workflow for RNA extraction, preparation, storage, and testing encompassing over 4 years' time can be found in Figure 1. Specifically, aliquots (3.4 μg) of total RNA were applied to RNAstable® (Biomatrica, San Diego, CA) or left unprotected in a 96-well plate format and allowed to dry overnight in a laminar flow hood (t=0). Dried samples were then stored inside a sealed moisture-barrier bag including a desiccant pack at RT or at 45°C to model accelerated aging.8–10 Aliquots of reference control samples (3.4 μg) were stored at −80°C. After 4.5 months, the −80°C samples were shipped on dry ice for overnight delivery, and the RNAstable and unprotected samples were placed into foil sealed bags with desiccant packets and shipped via ground delivery. Upon arrival, random aliquots of dried RNA were rehydrated in 30 μL nuclease-free water for 10 min and gently resuspended by pipetting. The plate containing unused dehydrated RNA was placed back into the foil bag with desiccant pack and stored in a dry-storage cabinet at RT until additional aliquots were resuspended at additional time points. Freezer control RNAs were also brought to 30 μL in nuclease-free water and stored at −80°C.
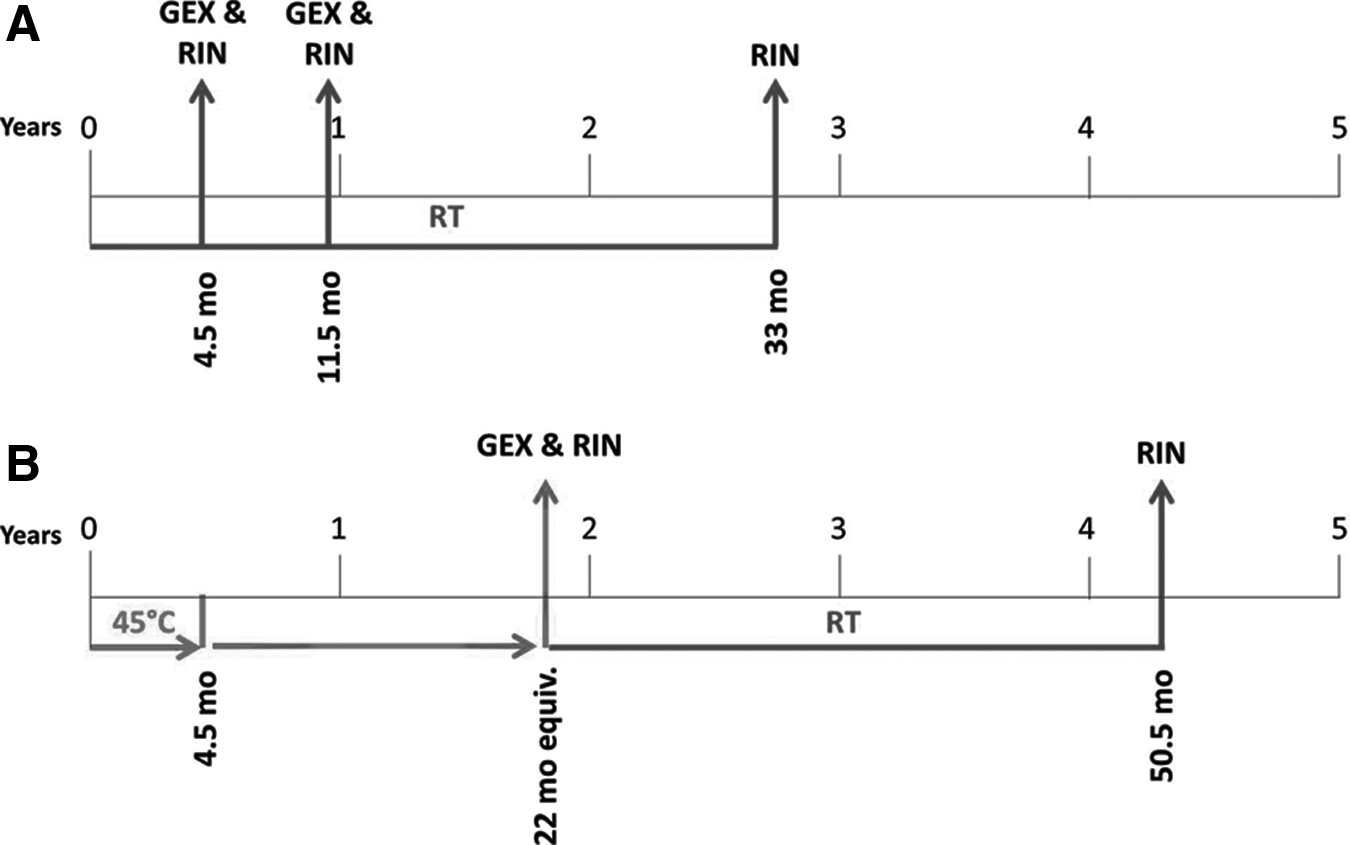
Timeline of sample storage and analysis in RNAstable®. Time=0 is when total RNA were extracted and applied to RNAstable or left unprotected prior to shipping.
Testing of RNA quantity and quality
Resuspended RNAs were quantitated using the NanoDrop-1000 (NanoDrop, Wilmington, DE). For the RNAstable-protected samples, wells containing dried RNAstable without RNA were rehydrated in 30 μL nuclease-free water to serve as the spectrophotomeric blank. To assess RNA integrity, each RNA was analyzed on the Agilent Bioanalyzer (Agilent Technologies, Santa Clara, CA) using the Total RNA Nano Protocol or the Small RNA Chip Kit for miRNA.
mRNA and miRNA expression profiling
150 ng total RNA from triplicate or quadruplicate aliquots of each storage condition was labeled and amplified using the Ambion Illumina TotalPrep-96 RNA Amplification Kit (Life Technologies). In this protocol, an oligo d(T) primer reverse transcribes RNA into cDNA starting from the 3′ end. The cDNA undergoes second strand synthesis and subsequent in vitro transcription to generate biotinylated cRNA. At 4.5 months or 11.5 months, the cRNA were hybridized to Illumina Human Ref-8 v3 Expression Bead Chips or the Human HT-12 v4 BeadChips (Illumina, Inc., San Diego, CA), respectively. Bead Chips were scanned using the Illumina Hi-Scan and images were processed using Illumina GenomeStudio v2011.1 software. For miRNA analysis, 200 ng of total RNA of each sample were prepared according to the Agilent miRNA Complete Labeling and Hyb Kit protocol. Labeled miRNAs were hybridized to Agilent Human miRNA v3 microarrays. Slides were scanned using the Agilent High Resolution C Scanner and images processed using Agilent Feature Extraction software.
Statistical analysis of microarray data
Statistical analyses (ANOVA with contrasts, t test and correlation computation, as well as PCA and hierarchical clustering) were performed by using Partek software combined with R and Perl scripts. Multiple-sample test correction (Benjamini and Hochberg) was applied to control for the false discovery rate (adjusted p value <0.05), unless otherwise indicated. A two-fold threshold was applied and only those probes were analyzed.
Results
Effect of RNAstable on RNA recovery and quality
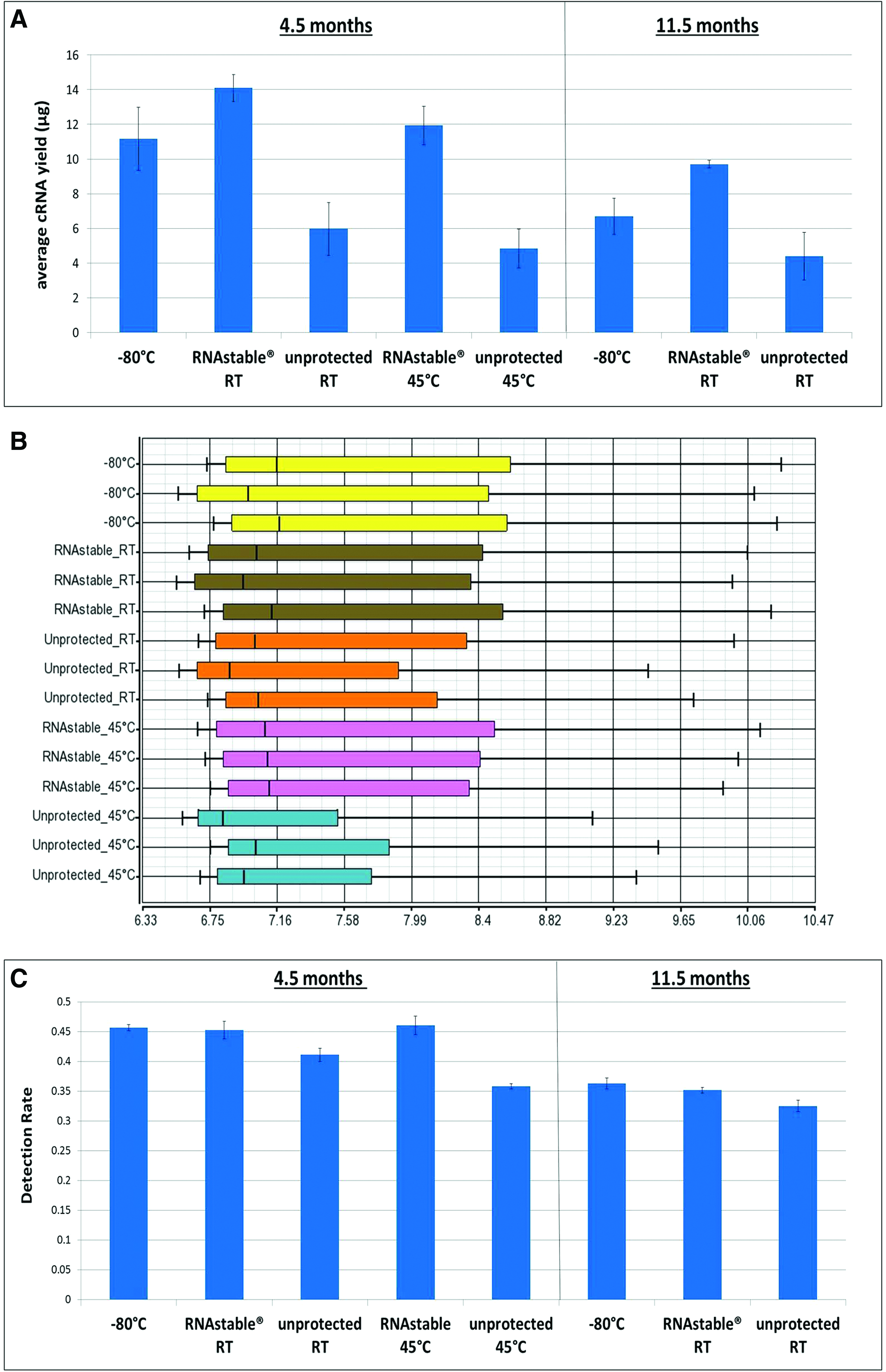
At 4.5 and 11.5 months, RNA replicates stored in the presence or absence of RNAstable® were rehydrated for quantitative and qualitative analyses. The majority of the samples yielded the expected amount of RNA (average=3.65 μg±0.85 at 4.5 months, and 3.80 μg±0.32 at 11.5 months) as measured by NanoDrop. After 4.5 months (n=3 for each condition), Bioanalyzer electropherograms showed that −80°C control RNAs gave an average RIN value of 9.5±0.4 (RIN=10 for highest quality). Similarly, RNA protected in RNAstable at RT resulted in an average RIN of 9.8±0.1. In contrast, unprotected RNA at RT produced an average RIN value of 3.3±0.2. In RNAs stored either at −80°C or in RNAstable at RT, electropherograms showed two distinct sharp peaks representing intact 28S and 18S RNA (Fig. 2A). However, in the unprotected RT RNA, these two peaks were only slightly visible as there was now a broad size range of degraded RNA. At 11.5 months (n=4 for each condition), unprotected RNA at RT produced a slightly decreased average RIN value of 2.5±0.2, consistent with the now almost nonexistent 28S and 18S peaks (Fig. 2C). RNAstable-protected RNA continued to show a high RIN of 9.6±0.0, comparable to the RIN of 9.6±0.3 observed for the frozen controls.

Evaluation of RNA integrity when stored in RNAstable. Replicates of extracted total RNA from 293 cells at the indicated storage condition were analyzed using the Bioanalyzer. Electropherograms show the degradation of a representative sample at 4.5 months at RT
Additional RNA aliquots were stored with or without RNAstable at the accelerated aging temperature of 45°C for 4.5 months (n=3). This is equivalent to 22 months of RT storage based on extrapolation from the Arrhenius equation.8–10 In Bioanalyzer electropherograms, the 45°C unprotected RNA was severely degraded, with only a broad peak of smaller RNA fragments seen and showing an average RIN of 2.3±0.0 (Fig. 2B). However, RNAstable-protected RNA stored at 45°C had an average RIN value of 9.8±0.1, which was similar to frozen controls (average RIN=9.5±0.4).
In a more recent study, we extended storage times of untested desiccated RNA and examined their integrity on the Bioanalyzer. In the first group, following the 11.5 month mark at RT in RNAstable, these aliquots were stored for an additional 21.5 months at RT for a total of 33 months (Fig. 1A). In the second set, untested desiccated aliquots originally stored at 45°C in RNAstable for 4.5 months were transferred to RT for an additional 28.5 months, which was equivalent to 50.5 months total RT storage (Fig. 1B). 8 Upon rehydration, we observed that average RIN values remained similar between RNAstable-protected RNA (average RIN=9.4) and frozen controls (average RIN=9.3) (data not shown). Unprotected RNA was considerably degraded to an average RIN of 2.3.
Effect of RNAstable on mRNA detection analysis
We evaluated the use of RNAstable-protected RNA in gene expression microarrays. Prior to hybridization, biotinylated cRNA were quantitated by NanoDrop. In comparison to the cRNA yield obtained for −80°C control RNA in triplicate at 4.5 months (11.2 μg±1.1), the cRNA yields with RNAstable-protected RNA stored at 45°C were similar (11.9 μg±0.7, p=0.49) and those stored at RT were significantly higher (14.2 μg±1.8, p=0.02). In keeping with RIN scores, unprotected RT and 45°C RNA produced the lowest average amounts (5.9 μg±1.5, p=0.0007, and 4.8 μg±1.1, p=0.00015, respectively) (Fig. 3A). A similar trend was observed at 11.5 months with samples tested in quadruplicate. Here the RNAstable-protected RNA yielded the highest cRNA levels (9.7 μg±1.3, p=0.002) compared to the −80°C control (6.7 μg±1.0), and the unprotected RNA showed the lowest amplification levels (4.4 μg±0.2, p=0.009) (Fig. 3A).
Quality control assessment of gene expression microarrays.
Following hybridization, scanning, and image data processing, a series of quality control metrics were performed to examine if RNAstable-protected RNA performed comparably to −80°C RNA. At 4.5 months, the distribution of microarray probe signal intensities revealed that in relation to frozen controls, there was a slight decrease in the mean intensity for samples stored unprotected at RT (p=0.0313), which was reduced further in the unprotected 45°C samples (Fig. 3B, p=0.0016). No significant difference was observed in mean signal intensities between RNAstable-protected RNAs and frozen controls [p=0.353 (RT) and p=0.578 (45°C)]. Similarly at 11.5 months, the intensities for the unprotected samples were visibly lower compared to the −80°C controls and RNAstable-protected samples (p=0.0004 and p<0.001, respectively, data not shown).
At 4.5 months, analysis of the percent of detectable probes on each array showed that hybridization of −80°C RNA resulted in similar detection rates (0.457±0.005) compared to those of RNAstable-protected RT RNA (0.452±0.014; p=0.646) and RNAstable-protected 45°C RNA (0.460±0.016; p=0.679) (Fig. 3C). A small but significant decrease in detection rates was observed in arrays hybridized with unprotected, RT RNA (0.411±0.011; p=0.0005). In agreement with the lower signal intensities in the unprotected 45°C RNA, we saw a noticeable drop in probe detection rates in these samples to 0.358±0.004 (p=7.36E-07). Seven months later, unprotected RNA continued to show significantly decreased detection rates (0.325±0.009) compared to −80°C controls (0.363±0.009; p=8.62E-05) and RNAstable-protected RNA (0.352±0.004; p<0.001) (Fig. 3C).
Calculation of the correlation coefficient (R) was performed to measure the strength of linear association of probe signal intensities when comparing detection profiles of all targets across each storage condition (Table 1). Strong correlations were observed within replicates of each sample storage condition at each time point (Table 1). At 4.5 months, frozen controls correlated well with RNAstable-treated samples stored at both RT and 45°C, but the level of correlation was reduced when comparing −80°C samples to those without RNAstable at RT (Table 1). The most striking differences were seen when comparing samples without RNAstable stored at 45°C to freezer controls (Table 1). At 11.5 months, freezer control RNA retained their similarity to RNA stored in RNAstable, whereas a slightly lower correlation was observed between either −80°C RNA or RNAstable-protected RNA and unprotected RNA at RT (Table 1).
The values are the mean correlation of Pearson r values.
Principal component analysis (PCA) was performed to show the level of variance in gene expression patterns among the different storage conditions. Consistent with the above data, PCA demonstrated that at both 4.5 months (Fig. 4A) and 11.5 months (Fig. 4B), frozen controls and RNAstable®-protected RNAs (RT and 45°C) clustered together. Unprotected RNA was separated from both −80°C and RNAstable-protected samples, indicating a higher degree of variance and decrease in similarity.
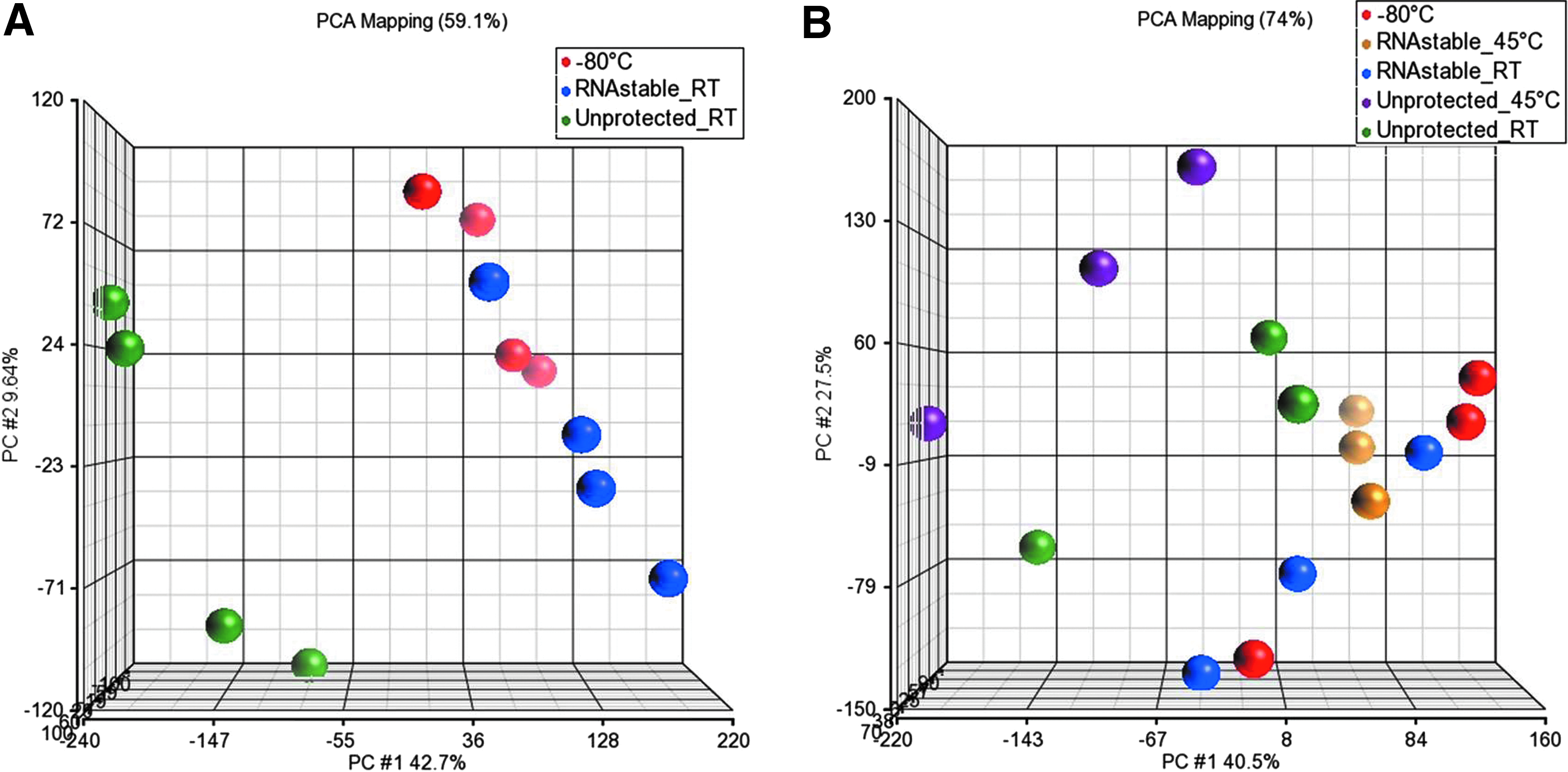
PCA plots based on all 24K probes
We compared array probes that demonstrated either enhanced or reduced detection in the presence of RNAstable. We first determined the number of probes altered between the various storage conditions, using an adjusted p value cutoff of p<0.05 and the requirement that the probe show ≥two-fold change in detection (Table 2). At 4.5 and 11.5 months, <0.1% of RNA targets stored in RNAstable had differential detection at either RT or 45°C in comparison to −80°C storage. However at 4.5 months, 3.6% and 15.3% of targets from unprotected RNA stored at RT or 45°C, respectively, exhibited differential detection compared to the same targets from frozen controls. At 11.5 months, RNA stored unprotected at RT showed 2.7% change in target detection compared to frozen controls and 3.8% change in relation to RNAstable-protected RNA. The majority of targets stored unprotected showed decreased detection as compared to RNA kept at −80°C or in RNAstable at both temperatures and time points (Table 2).
One-way ANOVA with paired contrasts were conducted and adjusted p values less than 0.05 (Benjamini and Hochberg multiple sample comparison) were applied to the final gene list. The table only included genes that have at least two-fold expression level changes.
Effect of RNAstable on microRNA detection analysis
At 10.5 months, additional replicates of RNA stored at RT were rehydrated and compared to frozen control RNAs (n=3 for all storage conditions). At all three storage conditions, all samples yielded similar amounts of RNA (data not shown). Small RNA integrity was assessed using the Bioanalyzer. In general, electropherograms showed that small RNA profiles were similar between those generated from −80°C and those stored in RNAstable; as per Agilent's documentation, all expected small RNA peaks were visible and clearly defined (Fig. 5A). The Bioanalyzer software calculated the averaged miRNA levels for the −80°C and RNAstable samples at 6.6% and 5% of all small RNAs, respectively. In contrast, unprotected RNA showed visibly more degradation (Fig. 5A) and therefore artifactually higher average estimated levels of miRNA at 9.3%.

Effect of RNAstable on miRNA expression.
All RNA replicates from each storage condition were assayed using Agilent miRNA microarrays and quality control parameters were examined. Similar fluorescent intensity values were observed across all samples with no visible difference observed between the freezer controls and unprotected or RNAstable-protected miRNAs (data not shown). Examination of miRNA profiles demonstrated that probe detection levels observed in RNAstable-protected samples strongly correlated with −80°C controls (Table 3). However, when compared to miRNA detection patterns derived from unprotected RNA, a lower correlation was seen with both −80°C controls and RNAstable-protected RNA (Table 3). Hierarchical clustering was performed to illustrate the level of similarity in miRNA detection patterns among the different storage conditions. Frozen controls and RNAstable-protected miRNA clustered close together (Fig. 5B). Similar to what we observed with the mRNA gene expression data, unprotected RNA show increased variance in relation to both −800C and RNAstable-protected samples and clustered separately.
The values are the mean Pearson correlation r values.
Discussion
Collectively these data demonstrate that total RNA stored in RNAstable for extended time periods, whether at RT or at 45°C, is not dissimilar in quality to that of RNA stored in ultra-low temperatures (-80°C). As evidenced by the comparable RIN values between RNAstable-protected RNA and frozen controls, RNAstable prevented loss of RNA integrity at RT after 11.5 months of storage. Maintenance of RNA quality was observed at 4.5 months storage at the accelerating temperature of 45°C, suggesting that RNAstable can protect RNA at RT for up to at least 22 months.8–10 When we lengthened RNA storage to 33 months for samples protected by RNAstable at RT, or to 50.5 months for samples originally in RNAstable at 45°C (for 4.5 months, then at RT), RIN values remained similar to frozen controls (RIN≥9.3; data not shown). These findings implied that the behavior of RNA stored in RNAstable would be unaltered in microarray hybridization experiments as compared to RNA kept at the standard temperature of −80°C. Indeed, multiple quality control metrics including cRNA labeling efficiency, hybridization probe signal intensities, and correlation analyses all suggest that RNAstable-protected RNA is suitable for use in gene expression microarray analyses.
Concordant with RIN scores, higher cRNA yields were generated from RNAstable-protected RNA in comparison to RNA stored in the absence of RNAstable. Additionally, due to the observed increase in degradation, overall hybridization signal intensities, and probe detection rates showed a decreasing trend in unprotected RNA relative to frozen controls and RNAstable-stored RNA. In the protocol used, biotin-labeled cRNA is generated from the 3′ end. It is therefore expected that as we see increased RNA degradation, fewer full-length cRNAs would be generated, and therefore we should observe decreased detection of probes that hybridize to targets more towards the 5′ end.2,11,12 Since the original unprotected RNA were substantially degraded, the lower signal intensities and detection rates are most likely caused by a lack of amplification of full-length RNA and a decrease in hybridization events of labeled cRNA due to missing cRNA sequence. In previous reports, Opitz et al. 11 and Thompson et al. 12 used microarray gene expression analyses to evaluate the mRNA location of probes that showed decreased detection in degraded RNAs. Probes with reduced signal intensities were located closer to the 5′ end of the mRNA, whereas those unaffected by degradation were near the 3′ region.
We also observed increased cRNA yields produced from RNAs stored in RNAstable at RT relative to frozen controls, even though the RIN values and hybridization signal intensities were similar. This indicates either that storage in RNAstable is conducive to more efficient cRNA amplification, or the freeze-thawing process of frozen RNA may affect mRNA stability that leads to lower levels of amplification. However, even though our overall results demonstrate that RNAstable efficiently prevents RNA degradation, 25 differentially detected probes showed ≥two-fold decrease in signal intensity in RNAstable-protected RNA at RT as compared to those stored at −80°C (Table 2). This suggests that a specific few mRNA sequences stored in RNAstable may show reduced protection over time. For example, we observed that BPNT1 is consistently less detected at 4.5 and 11.5 months; why this is requires further investigation. Since our study involved assaying four replicates per storage condition in only a single cell line, testing of a larger number of samples and use of confirmatory assays will be needed to further address these observations. We also noted that cRNA yields and probe detection rates were generally reduced across all storage conditions upon microarray testing at 11.5 months compared to at 4.5 months. This observation could be due to technical variability in the assay, storage time of reagents used in cRNA amplification, or use of an updated array platform.
In miRNA expression studies, we observed a decreased correlation in miRNA levels in samples stored unprotected at RT relative to frozen controls or RNAstable-protected RNA, whereas profiles of RNA stored in RNAstable or at −80°C were intermixed. We associated this finding with the increased levels of RNA degradation identified using the Bioanalyzer. Of note, unprotected RNA showed less defined small RNA peaks, a more broad trace, and had a higher estimated miRNA fraction than RNA stored at −80°C or in RNAstable. One explanation is that since the total RNA is more degraded, there is a larger proportion of fragmented mRNAs that contribute inaccurately to the miRNA percentage by congregating in the miRNA area (20–40 nucleotides). Due to their small size, it is assumed in the presence of conditions that degrade mRNA, miRNA remain intact. Recent literature gives conflicting reports as to whether mRNA degradation conditions impact miRNA stability. For example, Ibberson et al. 13 performed qPCR and microarray miRNA expression profiling studies in RNA from frozen tissues harvested into identical slices that were each induced to undergo varying degrees of degradation. RNA with lower RIN values showed an increased number of miRNA showing aberrant signal intensities. In contrast, Jung et al. 14 observed by qPCR that miRNA expression levels remained stable even in RNA with lower RIN values. Importantly, our results suggest that storing RNA in an unprotected state adversely affects miRNA expression to some greater extent over that of protected RNA either in RNAstable or −80°C. However, RNAstable-protected miRNA is more closely related to frozen miRNA.
In conclusion, utilization of both mRNA and miRNA expression assays demonstrated that RNAstable-protected RNA stored for up to 22 months at RT generated results that do not differ significantly from RNA stored frozen. Additionally, we hypothesize that even up to 50.5 months the excellent quality of the RNAstable-protected RNA as determined by Bioanalyzer analysis would produce microarray results not different from freshly extracted RNA. Importantly, these findings illustrate that stabilization of RNA in RNAstable is an alternative to storage of RNA at −80°C.
Footnotes
Acknowledgments
We acknowledge the contributions of Sharron Ohgi in preparation of the RNA from cultured cells and the initial RNAstable treatments as well as helpful discussions. We thank Illumina, Inc. for the gift of Illumina Human Ref-8 v3 Expression or Human HT-12 v4 BeadChips.
Author Disclosure Statement
No potential conflicts of interest are reported by NCI personnel. Rolf Muller is the founder, president, and CSO of Biomatrica, Inc. This research was supported in part by the Intramural Research Program of the NIH, National Cancer Institute, and internal resources of Biomatrica, Inc.