
Abstract
The advent of molecular characterization of tissues has brought an increasing emphasis on the quality of biospecimens, starting with the tissue procurement process. RNA levels are particularly affected by factors in the collection process, but the influence of different pre-analytical factors is not well understood. Here we present the influence of tissue specimen size, as well as the transport and freezing protocols, on RNA quality. Large, medium, and smaller porcine liver samples were stored either dry, on moist gauze, or in salt solution for various times, and then frozen in either liquid nitrogen or in pre-cooled isopentane. Large and small human liver samples were frozen in pre-cooled isopentane either immediately or after one hour at room temperature. The small samples were stored dry, on moist gauze, or in salt solution. RNA was isolated and RIN values were measured. The RNA for six standard reference genes from human liver was analyzed by RT-qPCR, and tissue morphology was assessed for artifacts of freezing. Experiments using porcine liver samples showed that RNA derived from smaller samples was more degraded after one hour of cold ischemia, and that cooled transport is preferable. Human liver samples showed significant RNA degradation after 1 h of cold ischemia, which was more pronounced in smaller samples. RNA integrity was not significantly influenced by the transport or freezing method, but changes in gene expression were observed in samples either transported on gauze or in salt solution. Based on observations in liver samples, smaller samples are more subject to gene expression variability introduced by post-excision sample handling than are larger samples. Small biopsies should be transported on ice and snap frozen as soon as possible after acquisition from the patient.
Introduction
R
While cold ischemia time (CIT) is known to compromise RNA integrity and gene expression,7–9 it is not known if the sample size and the transport method can influence RNA quality. When tissue is procured during routine macroscopic examination of a surgical specimen, it can be desirable to divide larger tissue samples into smaller pieces to avoid repeated handling of the frozen tissue sample. In cases where material is limited, it is not even possible to procure large samples. Tissues contain nucleases that vary in amount and composition with tissue type. 10 Cutting tissue into small pieces might release these damaging enzymes, degrading the RNA. Small samples are also more exposed to air and may sustain damage through oxidation. Evaporation of water during storage and transport can also increase salt concentration which may affect expression as well as degradation.
Slow freezing may lead to variation in RNA quality between the core of a larger sample that freezes later compared to its outer surfaces. Freezing tissue samples directly in liquid nitrogen or in pre-cooled isopentane are considered the best methods to avoid the formation of ice crystals that damage cells and local accumulation of salt that may lyse cells, 11 exposing RNA to extracellular nucleases. Snap freezing is also preferred for conserving tissue morphology. When tissue samples are frozen directly in liquid nitrogen, an insulating layer of nitrogen vapor forms around the sample that slows the freezing process (the Leidenfrost effect). 12 The Leidenfrost effect can be avoided by freezing tissue samples in an empty metal container that floats on the liquid nitrogen. This procedure also avoids the formation of large ice crystals in the tissue. 11 Pre-cooled isopentane can also be used to avoid the Leidenfrost effect and is widely accepted as the optimal way to preserve morphology of snap frozen tissue samples. 11 In a previous study, while the authors acknowledge the preservation effects of isopentane on morphology, it was not found to be better than nitrogen with respect to RNA quality. 13
Small samples such as needle biopsies can suffer air drying artifacts that damage tissue morphology and possibly also compromise RNA. To minimize evaporation, clinicians often send small samples to the laboratory either on moist gauze or submerged in isotonic salt solution. When biopsies are submerged in salt solution, the salt diffuses into blood vessels and other hollow structures in the tissue that can precipitate upon freezing. During the cutting of frozen sections for RNA isolation, excess salt may be carried over to the isolated RNA, compromising the yield and interfering with downstream biochemical processing. Excess salt may also affect tissue morphology.
In summary, while many possible complications have been described, there are little data showing if/how these various factors really affect RNA quality. Here, the effects of sample size in combination with transport and freezing conditions on tissue morphology, RNA integrity, and gene expression are reported. RNA quality is used as an indicator to find the optimal conditions for tissue procurement.
Materials and Methods
Exploratory experiments were performed on porcine liver samples to determine the main aspects of the relationship between sample size, transport and freezing methods, and RNA integrity. Based on these results, parameters were selected for the study of human liver. The quality parameters used to assess the conditions were the RNA Integrity Number (RIN) and RT-qPCR Cq values of the following reporter genes: hypoxanthine phosphoribosyltransferase 1 (HPRT1), hydroxymethylbilane synthase (HMBS), glucuronidase, beta (GUSB), peptidylprolyl isomerase B (PPIB), glyceraldehyde-3-phosphate dehydrogenase (GAPDH), and TATA box binding protein (TBP). The reporter genes selected belong to the group commonly used as reference genes and are occasionally referred to as ‘housekeeping’ genes. These genes are expressed in all cell types at rather constant levels.14,15 Variation in their expression therefore reflects changes introduced by the pre-analytical variation during the tissue procurement process.
Tissue samples
Normal liver (porcine and human) tissue was chosen for its homogeneity in order to minimize variation due to sampling. Porcine liver tissue was procured under approval number EMC 1878 (130-09-07) after study review by the Erasmus University DEC (animal experiment committee). The animals, a mixed breed of Landrace x Great Yorkshire, were acquired from a commercial pig breeder (meat production). They were housed on bedded, heated concrete floors, with unlimited access to water and food (as specified in the ETS No 123 and Dutch regulations). Animals weighing around 65 kilograms were anesthetized (with a cocktail of propofol, isoflurane, suffentanyl, and succinylcholine) and used for endoscopic operation technique training. Within 15 min after sacrifice (intravenous overdose of sodium-pentobarbital) by properly trained and certified veterinarian technicians (as described in article 12 of the Dutch animal experiment law; WOD) at the Erasmus MC Skills Lab, whole livers were removed and transported in less than 15 min to the pathology lab at room temperature.
To investigate the influence of sample size on RNA integrity, 12 large (8×4×4 mm), 12 medium (4×4×4 mm), and 12 small (2×2×2 mm) porcine liver samples were excised from a single liver. The samples were snap frozen on the bottom of an aluminium vial (PA6015, Sanbio, Uden, The Netherlands) floating on liquid nitrogen (“without isopentane”) either immediately (T0), or after 5, 15, 30, or 60 min storage at room temperature (Fig. 1A).
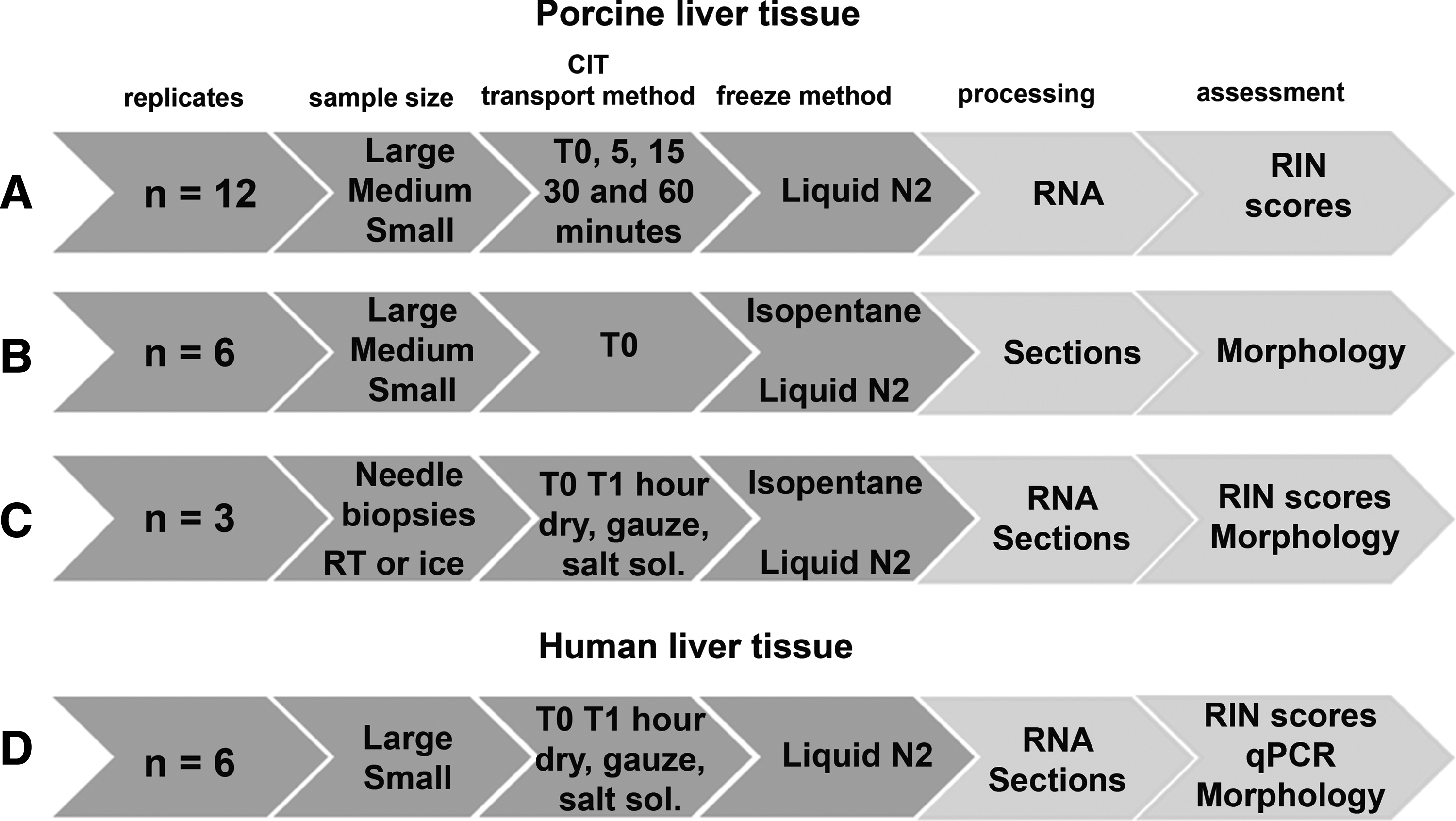
Collection and assessment conditions for porcine and human liver tissue. N=the number of samples tested for each condition. (For example,
To test how different freezing methods affect tissue morphology, six large, six medium, and six small porcine liver samples were prepared as described above and immediately snap frozen in either liquid nitrogen or pre-cooled isopentane. Tissue sections (4 μm) were cut and stained with hematoxylin and eosin (H&E) to assess the morphology (Fig. 1B).
To study the effect of transport or freezing method on RNA integrity and tissue morphology, needle biopsies (28 gauge, 2 cm) of porcine liver were taken and immediately frozen. Replicate needle biopsies were also stored for 1 h either dry, on moist gauze, or submerged in physiological salt solution (0.9% NaCl) at room temperature or on ice, and then snap frozen. Three biopsies were taken for each condition studied (Fig. 1C).
Human residual liver samples were procured according to the Dutch Code of Conduct legislation concerning the use of residual tissue for research. The Code of Conduct maintains an opt-out consent system and therefore, no written informed consent was required. The study and the consent regulation, as described in the Code of Conduct, 16 was approved by the Erasmus MC Medical Ethical Commission, under number MEC-2008-397. The tissue specimen was obtained after 3.5 h of surgery (warm ischemia) and 30 min of transport at room temperature (cold ischemia). All human liver tissue samples were collected during routine macroscopic examination of the specimen. Small (2×2×4 mm) and large (8×4×4 mm) samples (n=6 each) were obtained and frozen on the bottom of a metal container cooled in liquid nitrogen. The large samples were frozen either immediately, or after being stored dry for 60 min at room temperature. The small samples were frozen either immediately or after 60 min at room temperature either dry, stored on moist gauze, or submerged in isotonic salt solution. Sections (4 μm) from all tissue samples were cut and stained with H&E and submitted for morphological assessment (Fig. 1D).
RNA isolation and RIN measurement
Frozen 10 μm sections were cut and manipulated with pre-cooled tools and immediately placed in 700 μL Qiazol (Qiagen, Hilden, Germany). The amount of tissue used for RNA isolation was the same for all samples, using a smaller number of sections for the larger samples (10 sections from large samples, 20 from medium samples, and 40 from small samples). The time in the Qiazol solution was limited by analyzing no more than six samples at each time. The tissue sections in Qiazol were disrupted by shaking the tubes vigorously for about 5 sec. Total RNA was then isolated according to the manufacturer's protocol using the (mi)RNeasy mini kit (Qiagen, Hilden, Germany). The resulting RNA was placed on ice prior to RIN assessment on an Agilent bioanalyzer (Bioanalyzer 2100, Agilent, The Netherlands).
RT-qPCR on human liver tissue derived RNA samples
Complementary DNA (cDNA) was prepared from 2 μg RNA (measured with nanodrop, Isogen Life Science, The Netherlands) using the Thermo Scientific RevertAid H Minus M-MuLV First Strand cDNA Synthesis Kit, followed by a ribonuclease H step to degrade any remaining RNA as described previously. 17 To prevent amplification by the GAPDH assay of gDNA potentially still present in the RNA preparations, the samples were treated with a heat-labile double-strand specific DNase (HL-dsDNase) as described by the manufacturer (ArcticZymes, Norway) to remove any gDNA. Complete removal of the gDNA was confirmed using ValidPrime (TATAA Biocenter, Sweden).
The samples were diluted 20 times to a final cDNA concentration of 2.38 ng/μL prior to qPCR analysis, which was performed on the Stratagene Mx3000P platform (Agilent, The Netherlands) using 96-well plates. The six reporter transcripts were included in the analysis. Assays for TBP, HPRT1, and HMBS (designed by A. Sieuwerts) purchased from Invitrogen (Invitrogen, The Netherlands), were readily available as SYBR green primer assays (Table 1). The run protocol for the SYBR Green primer assays consisted of: Activation; 95°C 15 min (1 cycle), Amplification; 95°C 15 sec, 62°C 30 sec, 72°C 30 sec, 79°C 30 sec (40 cycles), followed by a melting curve analysis. A final reaction volume of 25 μL containing 11.9 ng cDNA, 330 nM of primers, and 50% (v/v) SYBR qPCR Master Mix with ROX (2-fold concentrated stock) from Abgene/Thermo Scientific was used for SYBR qPCR runs. Assays for GUSB, PPIB and GAPDH (designed by TATAA), purchased from Eurofins (MWG, Germany) were FAM-BHQ1 labeled primer/probe assays (Table 1). The run protocol for the primer/probe assays consisted of: Activation; 95°C 15 min (1 cycle), Amplification; 95°C 30 sec, 60°C 60 sec, (40 cycles). A reaction volume of 20 μL containing 11.9 ng cDNA, 200 nM primers, 50 nM probes, and 25% (v/v) Absolute qPCR Master Mix with ROX (2-fold concentrated stock) from Abgene/Thermo Scientific was used for all FAM-BHQ1 qPCR runs.
According to the Serial Analysis of Gene Expression (SAGE) mammalian gene collection (mgc_p5s) database (http://cgap.nci.nih.gov/SAGE/).
Based on a 4-fold dilution series of pooled, fresh frozen tissue-derived cDNA samples; PCR efficiency of 2.00 is considered perfect.
For gene expression analysis, the difference between the Cq value of the stored sample and the average Cq values of large samples frozen at T0 were calculated by using the formula: ΔCq=Cq size Tx – Cq large T0. A ΔCq value of 1.0 corresponds to a two-fold decrease of template RNA.
Morphological assessment
The morphological quality of all samples of both porcine and human origin was assessed, blinded to the method of transport or freezing. For each of the experiments described in Figure 1, the pathologist created two groups of slides, those not displaying considerable morphological artefacts versus those with morphological artifacts that could compromise diagnosis. If no considerable morphological differences were observed within one experiment, the pre-analytical parameters studied did not adversely influence morphology and were not presented. The Fisher exact test was used to analyze whether there was a significant correlation between aberrant morphology and the transport methods.
Statistical analysis
All RIN scores and RT-qPCR Cq values were collected in IBM SPSS version 21 for statistical analysis. Owing to the small test numbers used for each sample-handling condition, the non-parametrical Mann-Witney U test was performed to assess differences between the experimental groups, and the Fisher's exact test was used to analyze the associations between categorical variables. All tests were two-sided, and unless indicated otherwise, p<0.05 was considered statistically significant. For RT-qPCR data and in consideration of the small number of samples and the relatively high number of measured genes, a Bonferroni-Holm multiple testing corrected cut-off p-value of 0.0083 was considered significant.
Results
RNA integrity and morphology of porcine liver tissue
Sample size
The results of the RNA integrity analysis expressed as RIN scores in relation to sample size (described in Fig. 1A) are shown in Figure 2. Large samples showed no significant decrease in RIN score during the time course (0–60 min). Medium sized samples showed significant RNA degradation after 60 min (T0 vs. T60, p=0.001; T5 vs. T60, p=0.041; T15 vs. T60, p=0.006; T30 vs. T60, p=0.001; Mann-Witney U test). Although small samples seemed to show RNA degradation earlier, this decrease is not significant (p=0.05; Mann-Witney U test) when measured between T0 and T60.

Influence of sample size on porcine liver RNA integrity. Storage time of the samples kept at room temperature until freezing in liquid nitrogen is shown on the X-axis. P values for RNA degradation for medium-sized samples after 60 minutes was as follows: *T0 vs. T60, p=0.001; **T5 vs. T60, p=0.041; ***T15 vs. T60, p=0.006; ****T30 vs. T60, p=0.001; (Mann-Whitney U test).
Transport and freezing methods
The method of transport, simulated by storing porcine liver needle biopsies for 60 min either dry, on a moist gauze, or in salt solution (experiment described in Fig. 1C) did not significantly influence RNA integrity. Keeping the biopsies in a container placed on wet ice for 60 min gave significantly better results (p=0.026; Mann-Witney U test) in contrast to keeping the biopsies at room temperature for the same amount of time.
Frozen section artifacts such as vacuoles in nuclei and ice damage in the cytoplasm were observed in 3 of the 17 sections when needle biopsies were stored dry; in 6 of the 18 sections when stored on moist gauze; and in 9 of the 17 sections when stored in salt solution. These differences were, however, not significant (p=0.07, Fisher's exact test).
When morphology between liver samples snap frozen without isopentane was compared to samples frozen in pre-cooled isopentane (described in Fig. 1B), no considerable differences were observed.
RNA integrity and gene expression of human liver tissue
Sample size
The results of RNA integrity analysis expressed as RIN score in relation to sample size and method of transport (described in Fig.1D) are shown in Figure 3. There was a significant decrease in average RIN score between large samples frozen at T0 and those held for 60 min at room temperature (ΔRIN=0.3; p=0.015; Mann-Witney U test). This same decrease in RNA integrity was more pronounced in small samples (ΔRIN=0.85; p=0.015; Mann-Witney U test) (T0 vs. T60 dry).

Influence of sample size and method of transport on human liver RNA integrity. The x-axis shows, respectively, the large (L) samples at T0 and T60 (60 min), transported dry (D) *p=0.015; the small (S) samples at T0 and T60 transported dry (D) **p=0.015, on moist gauze (G) ***p=0.009, and in salt solution (S) ****P=0.002. The level of RNA degradation in large samples is relatively low (ΔRINLT0-LT60D=0.3) compared to that in small samples (ΔRINST0-ST60D=0.85).
To measure the effect of cold ischemia and sample size on gene expression, six common reference genes were assessed with RT-qPCR. Using the formula ΔCq=Cq large T1 – Cq large T0 (Fig. 4A), our results indicate that 1 h of cold ischemia time did not appear to significantly alter gene expression in large samples; in small samples, using the formula ΔCq=Cq small T1 dry – Cq small T0 (Fig. 4B), the levels of three reference genes were significantly lower. P-values of the genes with significantly altered expression are presented in Table 2.
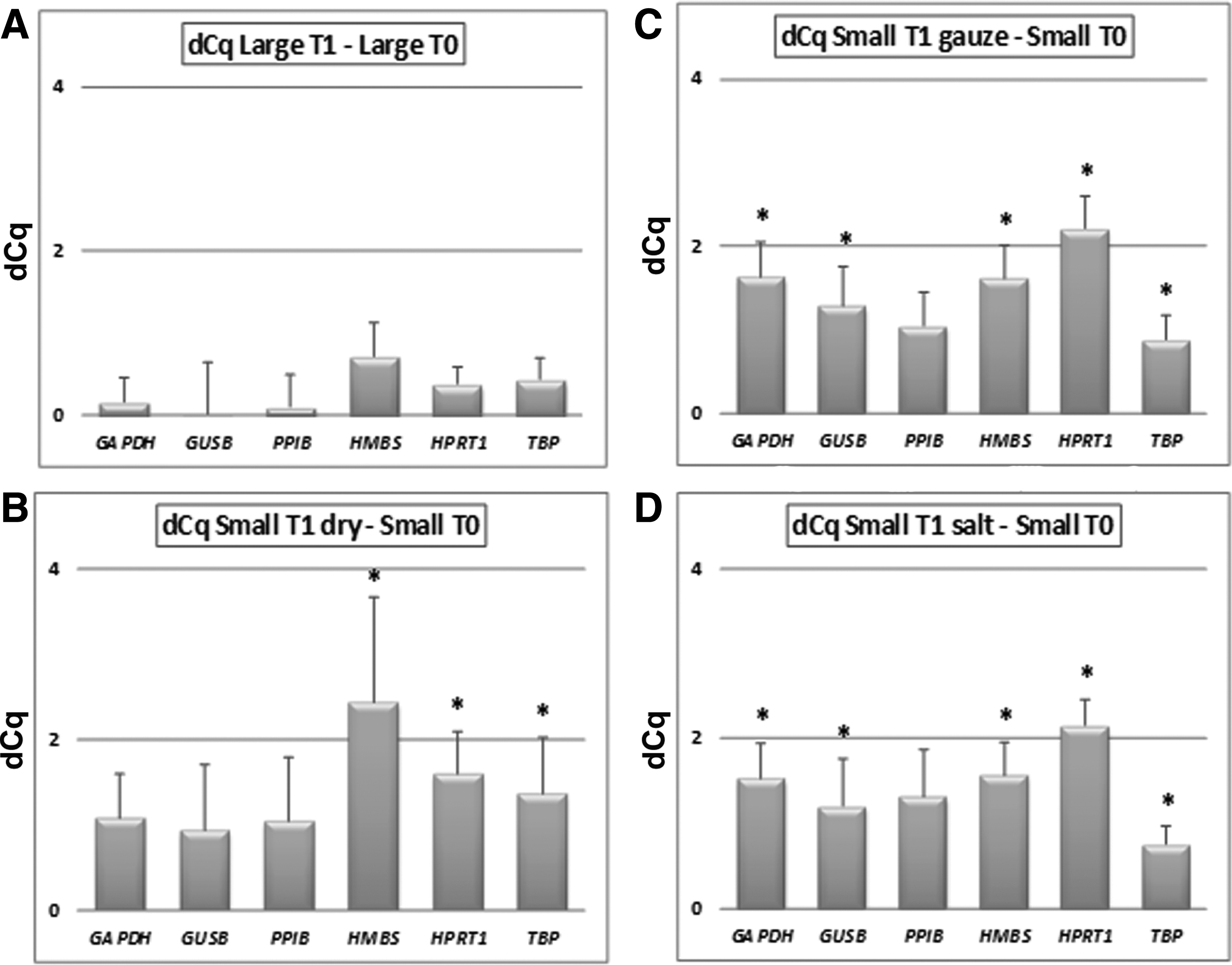
Effect of sample size and method of transport on gene expression in human liver tissue. A positive dCq indicates a decrease in gene expression of the test time point or condition as compared to the control. Significant dCq's after multiple corrections are marked with an asterisk (*). The error bars represent standard deviations for n=6 measurements. One dCq corresponds to a two-fold decrease in gene transcript copy number (original number of mRNA copies per cell).
P-values Mann-Whitney U test; *Significance level RIN score P<0.05 (
GAPDH, glyceraldehyde-3-phosphate dehydrogenase; GUSB, glucuronidase, beta; PIPB, peptidylprolyl isomerase B; HMBS, hydroxymethylbilane synthase; HPRT1, hypoxanthine phosphoribosyltransferase1; TBP, TATA box binding protein.
Method of transport
There was no significant difference in RIN scores among the three transport methods (dry, moist gauze, and salt solution) (Fig. 3), neither were considerable morphological differences observed. When small samples were transported on moist gauze (ΔCq=Cq small T1 gauze – Cq small T0; Fig.4C) or in salt solution (ΔCq=Cq small T1 salt solution – Cq small T0; Fig. 4D), levels of all six reference genes were significantly lower compared to the samples transported dry. P-values of the genes with significantly altered expression are presented in Table 2.
Discussion
As previously shown, cold ischemia influences RNA integrity (RIN values) and gene expression (Cq values).18–20 The effect of cold ischemia temperature (i.e., room temperature vs. 4°C) has also been studied. 21 However, additional factors that affect RNA integrity during the cold ischemic phase have not been previously analyzed. Therefore, we evaluated RNA integrity, gene expression, and tissue morphology in human and porcine liver samples of various sizes, subjected to different transport and freezing conditions, during cold ischemia.
The differences in RNA integrity and/or Cq values were used as indicators to identify the optimal tissue acquisition procedure. Our results indicate that RNA integrity and gene expression in smaller size samples are more distorted by 1 h of cold ischemia than in larger size samples. Although the differences in RNA integrity and gene expression seem minor in a practical sense, the preferred acquisition method should be the one that results in the best quality RNA.
Liver was chosen for these experiments because of tissue homogeneity that kept sampling errors to a minimum; the effects of sample size and method of transport in other tissue types may be different or non-existent, and needs further investigation.
When comparing different methods of transport of human liver samples, no significant differences in RNA integrity (RIN scores) were observed. At the gene transcript level, however, it was found that transport on moist gauze and in salt solution resulted in lower gene transcript levels compared to dry transport. This also shows that measuring gene expression is a more sensitive way to monitor RNA quality than is determining RIN score.
The reason more variation was observed in the gene expression of the three reference genes determined using the SYBR Green primer assays is likely the consequence of the differences in the sizes of the detected transcripts: 94, 139, and 192 bp for the SYBR-based assays versus 87, 91, and 92 for the probe-based assays. The more degraded the material, the more difficult it is to detect larger transcripts.
Since the RNA integrity of several tissue types is not influenced by the freezing method 13 and morphology is not harmed, tissue banks may consider terminating the use of isopentane after further internal validation. However, for diagnostic purposes such as enzyme histochemistry on frozen sections of muscle biopsies, where optimal morphology is of crucial importance, it is still necessary to use isopentane for optimal results. 22
The differences between RNA integrity in human (small samples) and porcine tissue (needle biopsies) in more or less comparable experimental settings might be explained by the fact that the liver was taken from the animals only minutes after the animals were euthanized. These livers were subjected to only a short period (<15 min) of post-mortem warm ischemia, and as samples were taken shortly after the organs were transported to the pathology department, cold ischemia time (15 min) and damage to RNA was probably minimized. The human tissues, taken from a surgically removed liver segment, were exposed to relatively extensive surgical warm ischemia times (report is inconclusive, but at least 3.5 h) and cold ischemia times (30 min consisting of transport and the wait for macroscopic examination), which may have been responsible for the more extensive RNA degradation seen compared to that in porcine tissues.
For the studies involving human liver samples, the dCq as defined in the Materials and Methods section was between 1.5 and 2 Cq (Fig., 4) which reflects a three- to four-fold variation in gene transcript copy number. It is difficult to identify which cellular stress mechanisms, in addition to the decrease of available intact RNA molecules, might have driven the variable transcript levels of the standard reference genes, as observed between tissue samples exposed to different handling conditions. It was surprising to observe that sample size was a greater driver of differential gene transcript level than cold ischemia time. Moreover, when the biopsies were placed on moist gauze or in salt solution, the effect seemed more pronounced for some genes, suggesting that additional regulation besides poor RNA quality due to the hour of cold ischemia likely played a role.
In conclusion, based on observations in liver tissue (other tissue types may behave differently), it seems best not to procure small samples or divide large samples into smaller pieces when the possibility to procure larger samples exists. Using RT-qPCR rather than the relatively crude RIN assessment, we were able to conclude that when transporting small biopsies to the pathology department, it is best to keep them in a dry environment with the container cooled on ice to avoid loss of morphological integrity and to minimize gene expression variability. To prevent air drying artifacts, it is advisable to use small containers that are tightly closed. To avoid post-surgical RNA degradation and gene expression variation, it is of high importance to freeze small biopsies as soon as possible after they have been procured from the patient.
Footnotes
Acknowledgments
This research received funding from the European Union Seventh Framework Programme [FP7/2007–2013] under grant agreement nr 222916. AMS was supported by Cancer Genomics Netherlands funded by the Netherlands Organization for Scientific Research (NWO). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. The authors would like to thank Lynne Rainen for her native English editing, and colleagues at the Erasmus MC Department of Medical Oncology and the Erasmus MC Skills Lab for their hospitality and support.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.