
Abstract
The ability to compact and inject the cat germinal vesicle (GV) into a recipient cytoplast allows exploration of a new fertility preservation strategy that avoids whole oocyte freezing. The objective of the present study was to understand the impact of water loss and storage time on GV DNA integrity. Immature cat oocytes were exposed to 1.5 M trehalose for 10 min before microwave-assisted dehydration for 0, 5, 10, 15, 20, 25, 30, or 40 min. Oocytes then were rehydrated to assess chromatin configuration and the incidence of DNA fragmentation (TUNEL assay). The moisture content progressively decreased (p<0.05) from 1.7 to 0.1 gH2O/gDW over the first 30 min, but did not decrease further (p>0.05) after 40 min. Chromatin configuration was unaffected (p>0.05) over time. The percentage of GVs with DNA fragmentation was unaltered (p>0.05) from 0 to 30 min of treatment (range, 6.1%–12%), but increased (p<0.05) to 32.5% after 40 min. Next, the influence of storage at two different supra-zero temperatures after 30 min of drying was investigated. Oocyte-loaded, microwave-treated filters were individually sealed in Dri-Shield moisture barrier bags and stored at 4°C or ambient temperature for 0 to 8 weeks. Moisture contents gradually decreased (p<0.05) from 0.12 to 0.10 gH2O/gDW after 8 weeks of storage at 4°C or ambient temperature. The percentage of GVs with DNA fragmentation more than doubled (p<0.05) from 0 (14.3%) to 2 days (30.0%–33.0%), but remained stable (p>0.05) thereafter (1 through 4 weeks, 25.0%–35.0%). Collective results demonstrate the feasibility of using microwave processing to dehydrate the mammalian GV to a moisture content that is nonlethal and enables nonfrozen storage, an alternative approach for preserving the maternal genome at cool or ambient temperature.
Introduction
W
More specifically, the GV is the organelle surrounded by a nuclear envelope that is present within all immature oocytes and which contains (1) de-condensed chromatin arrested at prophase I of meiosis, and (2) nucleoplasmic factors that mix with the cytoplasm during meiotic resumption, both of which are essential for subsequent fertilization.7,8 Although still at the initial stage of meiosis, the GV is already a highly specialized nucleus that differs from diploid somatic cell nuclei in that it has a ∼10-fold larger volume and specific epigenetic patterns.9,10 Recovery, preservation, and subsequent transfer of GV into a competent, enucleated oocyte is also the only viable option to rescue the genome from (1) an oocyte that has mitochondrial issues, (2) one that is morphologically-abnormal or poor quality that consistently fails to mature or fertilize in vitro (including from an aging donor), or (3) one that is incompetent due to being derived from a preantral or early antral follicle in a pre-pubertal or adult ovary. 11 The value of GV transfer has been envisioned in many species, including humans. 12
There is now a growing body of evidence on the value of the laboratory cat as a reproductive model for humans, largely due to intriguing morphological and biological similarities between the ovaries and oocytes of the two species, including practical issues such as fertility preservation.4,13 More specifically, substantial knowledge has emerged on the immature cat oocyte, which we have asserted is relevant to enhancing our overall understanding of oocyte competence and GV functionality. 4 Cat GV chromatin is remarkably tolerant to osmotic stresses, artificial chromatin compaction, and freezing temperatures.5,14 This observation underpins our contention that the cat is a germane model for exploring the plasticity and resilience of the GV to different external environments. 10
Interestingly, the ability to artificially compact and inject the cat germinal vesicle (GV) into a recipient cytoplast allows exploration of a new fertility preservation strategy that avoids whole oocyte freezing and could enable drying and storage of GV at supra-zero temperatures. 11 This could avoid the high cost of refrigerated storage. 15 The concept of anhydrous preservation, or preservation in a dry state, is a natural phenomenon that some species use to survive desiccating conditions. These ‘anhydrobiotes’ frequently produce the glass-forming disaccharide trehalose, which serves as both a biomolecular protectant, and a highly viscous matrix that slows molecular motions that lead to degradation.16–18 Previous studies demonstrated that (1) 60% of GVs can survive simple air-drying in the presence of trehalose followed by 4 weeks of storage at 4°C, and (2) 10% of reconstructed oocytes with rehydrated GV can achieve nuclear maturation. 19 However, air-drying is an uncontrolled process that often results in non-uniform distributions of water within and between samples. Compared to existing techniques, microwave-assisted dehydration appears to be a simple and reliable approach to dry biological samples at ambient temperatures,20,21 while reducing the exposure time of biologics to damaging high osmolar liquid solutions. 22
Loss of cellular viability can occur at two key stages: during processing into the dry state and during long-term storage. Because a substantial reduction of molecular mobility is necessary to ensure an extended shelf life, depending on the desired storage temperature, it is necessary to dry the sample to different levels, in order to ensure that a glassy state is achieved.23,24 Previous work with macrophages has shown that damage accrues as cells are processed to lower moisture contents, thus it is important to determine the extent of moisture loss that can be tolerated by GVs during processing in order to establish a realistic storage goal.20,22 Furthermore, while the ability of the GV to achieve nuclear maturation is a good indicator of viability, it does not provide insights into the integrity of the DNA, which could later be manifest as defects in a fertilized embryo. 1 The effect of processing conditions on DNA integrity is thus a critical indicator for identifying a suitable end-point for drying. The objectives of the current study therefore were to assess (1) the impact of microwave-assisted drying on GV structural integrity (chromatin configuration and incidence of DNA damage), and (2) the sensitivity of GV to dehydration/storage processing, with the goal of establishing an appropriate moisture level for dry storage. Towards this end, processing parameters were also determined in order to prescribe specific drying end-points.
Materials and Methods
Oocyte collection
Ovaries from adult domestic cats were recovered after routine ovariohysterectomy at local veterinary clinics and transported in phosphate-buffered saline (PBS) at 4°C to the laboratory within 6–12 hours of excision. Cumulus-oocyte-complexes (COCs) were collected from antral follicles by repeatedly slicing the ovaries in Hepes-buffered minimum essential medium (H-MEM; Gibco Laboratories, Grand Island, NY, USA) supplemented with 1 mM pyruvate, 2 mM L-glutamine, 100 IU/mL penicillin, 100 μg/mL streptomycin, and 4 mg/mL bovine serum albumin (Sigma-Aldrich, St. Louis, MO). Each COC from antral follicles was classified according to the standard quality criteria. 25 Only grade 1 (uniformly dark cytoplasm, ≥5 compact layers of cumulus cells) and grade 2 (same as grade 1, but with <5 cell layers) oocytes were collected. The oocytes were then denuded of cumulus cells by exposure to 0.1% hyaluronidase (Sigma-Aldrich) for 15 min at 38°C, followed by vortexing and rinsing with H-MEM.
Drying solution and process
Denuded oocytes were permeabilized with 10 μg/mL hemolysin for 15 min. Because preservation of the GV was the goal of the current experiments, cytoplasmic membranes were permeabilized to enable trehalose loading. Thus management of cell volume changes typical during cryopreservation processing was not a concern for these studies. In order to minimize the cumulative exposure to high concentrations of sugars that could potentially damage the GV, permeabilized oocytes were immersed in 1.5 M trehalose solution in Tris-EDTA buffer (pH 8.00) for 10 min and then immediately dehydrated. The viscosity of this 1.5 M solution was low enough to enable pipetting and sample handling, yet enables the overall drying time to be reduced compared to starting the drying process from an isotonic condition. 22
An inexpensive conjugate release glass fiber matrix (GE Healthcare, Pittsburgh, PA) was selected as the drying surface, which allowed for simple loading and recovery of GVs by micropipette. This filter, already widely used in clinical applications, was cut to a 13 mm size to enable its use in a Swinnex Filter Holder. This filter holder approach can enable sample drying, storage, and rehydration in a single aseptic container. A customized microwave turntable was used in a SAM 255 microwave (CEM, Matthews, NC) to enable batch processing (max. capacity=12 filters) of samples contained in these filter holders. 21 A set of up to 5 oocytes, along with 40 μL of trehalose solution, were transferred onto each conjugate-release filter paper and dehydrated with the SAM 255 microwave. A control filter without oocytes was processed with each batch at each time point to assess the moisture contents immediately after microwaving or storage. The conjugate release membrane allowed for optimal physical recovery of the dried samples, as the GVs easily detached from the glass fiber matrix upon rehydration with 400 μL of H-MEM medium supplemented with pyruvate, L-glutamine, penicillin, and streptomycin. Recovered oocytes were fixed in 4% paraformaldehyde in PBS for the assessment of chromatin configuration (Hoechst staining) and DNA fragmentation (TUNEL assay).
Assessment of mass loss kinetics during microwave drying
In order to prescribe specific drying end-points, a drying curve was established for the 1.5 M trehalose buffer solution at 20% microwave power, with the end moisture content determined for every 5 min interval of processing (n=46; five to six replicates), up to 40 min, at which time sample drying reached its equilibrium point. Drying curves were established in an ambient relative humidity of 23.2%–52.2%. Integral to the development of this curve was the use of Karl Fisher titration (Mettler-Toledo V20), a technique that determines water content of samples accurately to levels as low as 100 ppm. The volumetric titrator enabled a rapid determination of the water content during drying, as the entire filter could be loaded into the reaction vessel, reducing the time and subsequent error associated with gravimetric mass determination using an analytical balance. The moisture content is presented as gram H2O per gram dried weight (gH2O/gDW). While the drying solutions in this study contain 10 mM Tris and 1 mM EDTA, the glass transition temperature (Tg) of the solution is not likely to be altered significantly by such a small mass contribution.
24
The Tg estimates were thus obtained using a Gordon-Taylor fit of trehalose-water data according to:
where k=5.2, Tg of trehalose is 100°C, and Tg of water is −135°C.22,23
Assessment of DNA integrity
DNA fragmentation was detected by terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay using an in situ cell death detection kit (Roche Applied Science, Indianapolis, IN). Manufacturer's assay recommendations were slightly modified as follows. Fixed oocytes were rinsed three times in PBS containing 1 mg/mL polyvinylpyrrolidone (PVP; Sigma-Aldrich) and then permeablized with 0.5% Triton X100 solution for 30 min at room temperature. After three washes in PBS-PVP, oocytes were exposed to the TUNEL solution for 1 h at 38°C. Oocytes then were washed three times in PBS-PVP before staining the chromatin with 10 μg/mL Hoechst 33342 (Sigma-Aldrich) for the assessment of chromatin configuration. Each trial included a positive control (exposed to DNAse for 10 min prior to TUNEL staining) and a negative control (incubation in labeling solution instead of TUNEL solution). All oocytes were mounted on slides with Vectashield (Vector Labs Inc., Burlingame, CA). Each slide was examined under an epifluorescence microscope (Olympus BX41; Olympus Corporation, Melville, NY) using SPOT software 5.0 (Diagnostic Instruments, Inc., Sterling Heights, MI). TUNEL positive/negative oocytes were classified based on the presence/absence of green fluorescence in GVs (Fig. 1). Absence of GV (no Hoechst staining) likely revealed the loss of the GV during the handling or drying process. The proportion of TUNEL positive GV was calculated relative to the number of oocytes with a GV present. The proportion of intact GV was estimated relative to the total number of oocytes (including those with absent GVs).
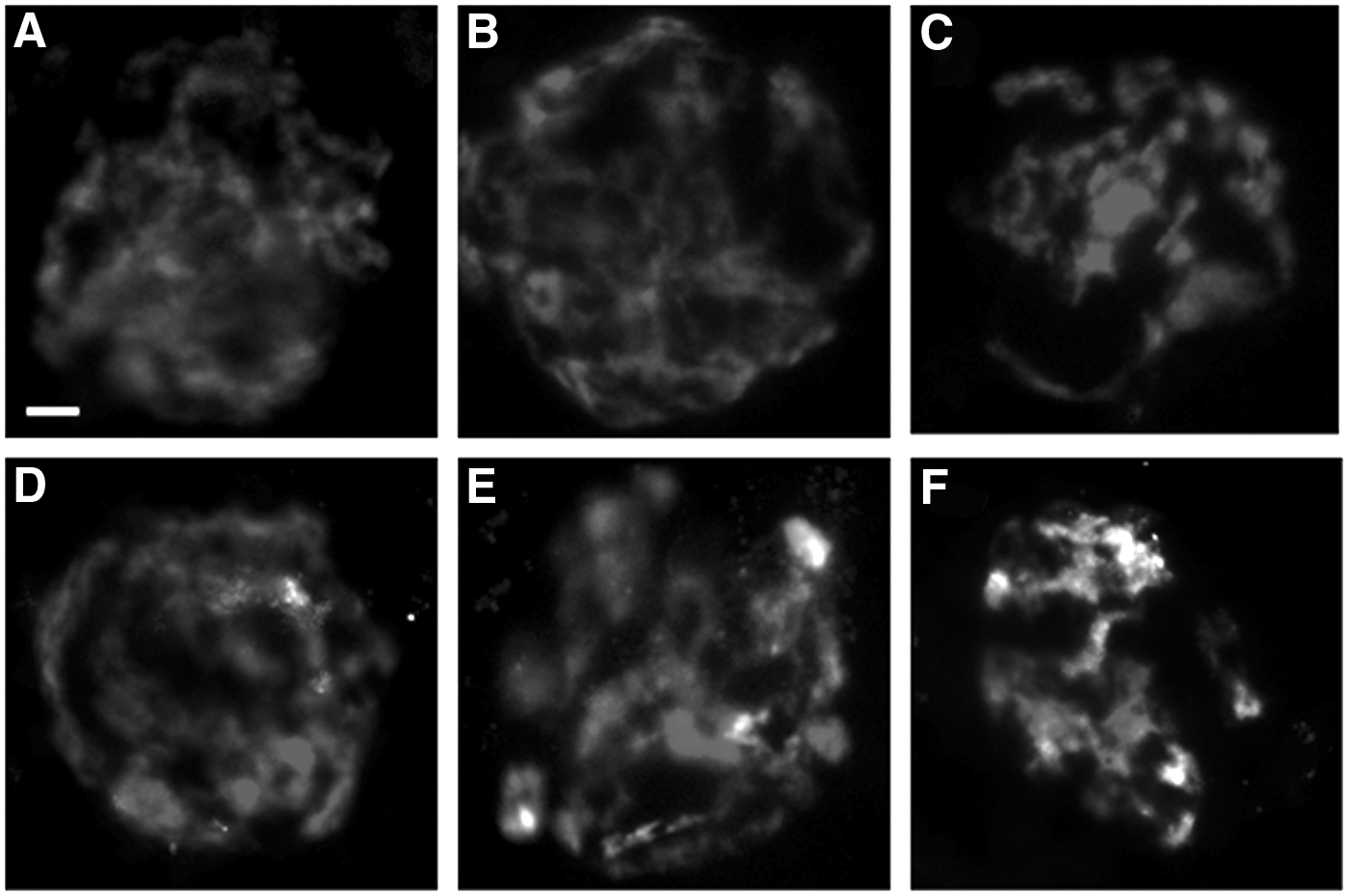
Representative chromatin configuration (Hoechst staining) and DNA integrity (green TUNEL staining) after microwave-assisted drying for 40 min.
Experimental design and statistical analysis
A preliminary experiment was conducted to optimize the concentration of trehalose in the drying solution and the droplet size, allowing efficient drying and ease of oocyte recovery after rehydration. TUNEL assay and Hoechst staining were then carried out to assess the sensitivity of chromatin to processing conditions and the associated chemical stress that accumulates as drying progresses to lower moisture contents. Experiment 1 assessed the DNA damage during the drying process. Oocytes selected from a pool of different ovaries collected on a given day (one replicate) were allocated randomly for different treatments for each experiment. Samples (n=337 total oocytes, 4 to 5 replicates) were dehydrated at 20% power for 0, 5, 10, 15, 20, 25, 30, or 40 min. Moisture contents on replicate samples were measured immediately after microwaving using volumetric Karl Fischer titration. Oocytes were rehydrated to assess chromatin configuration (Hoechst staining) and DNA fragmentation (TUNEL assay). Based on these findings, Experiment 2 (n=160 total oocytes, 2 to 5 replicates) was conducted to examine the influence of storage at two different supra-zero temperatures after 30 min drying. Oocyte-loaded, microwave-treated filters were individually-sealed in Dri-Shield moisture barrier bags and stored at 4°C or ambient temperature (21.4°–23.9°C) for 0 or 2 days or 1, 2, 4, or 8 weeks. Moisture contents and DNA fragmentation were assessed at the end of the storage periods. Data from all replicates were pooled to determine the overall percentage of each treatment group. Comparisons between treatment groups were evaluated by chi-square test. Differences were considered significant at p<0.05.
Results
Solution drying kinetics and predicted Tg
A microwave-assisted drying mass loss curve was initially established for the trehalose drying solution (Fig. 2). Relative humidity (RH) fluctuated between 23.2 and 52.2% over the course of the study. The majority of subsequent experiments were also conducted under similar conditions. Moisture contents were observed to progressively decrease (p<0.05) from 1.69 to 0.11 gH2O/gDW over the first 30 min (with a rapid decrease during the first 10 min; Fig. 2) but were not significantly lower (p>0.05) between 30 and 40 min (Fig. 2). The predicted glass transition temperature (Tg; the temperature at which the glass transitions from liquid-like to solid-like behavior) of the binary trehalose solution provided insights into a suitable storage temperature for long-term storage (Fig. 2). The moisture level achieved in the composition at 30 min was predicted to yield a Tg of ∼18°C based on published data for binary trehalose compositions.23,24 It should be noted that this estimate served only as a rough guide, as measurements of Tg vary widely from study to study, and our composition was more complex than those that have been studied to date. Furthermore, measurements of Tg will vary based on experimental conditions, and cannot be used to replace shelf life studies.

Water content in gH2O/gDW (left axis) and predicted Tg of 1.5M trehalose in 1X TE buffer (right axis) as a function of drying time. Drying curves were established at 23%–52% RH. The inset graph provides the moisture content region between 0.1 and 0.16 gH20/gDW at higher resolution.
Impact of the microwave exposure on DNA integrity
Oocytes in 40 μL of 1.5 M trehalose solution were loaded on filters and dried for 0, 5, 10, 15, 20, 25, 30, or 40 min. GV chromatin configuration did not change as a result of exposure to microwave energy of any duration (Fig. 1). Proportions of empty oocytes (range, 2.2%–8.3%) did not change (p>0.05) over time except at 25 min (9.1%; Table 1). The incidence of GVs with DNA fragmentation (TUNEL positive relative to the total number of oocytes with a GV) was stable (p>0.05) from 0 to 30 min of treatment (range, 6.1%–12%) and increased (p<0.05) to 32.5% after 40 min (Table 1). As a result, the proportion of intact oocytes (relative to the total number of oocytes subjected to the treatment) was slightly lower (p<0.05) after 25 min than the other time points and negatively affected after 40 min (p<0.05; Table 1).
Percentage of oocyte without GV relative to total number of oocytes; **percentage of TUNEL positive oocytes relative to the total number of oocytes with GV; £percentage of oocytes with intact DNA relative to the total number of oocytes; Within columns, proportions with different superscripts differ ( p<0.05, Chi-square).
Impact of storage time and temperature on the DNA integrity
Because the glass transition temperature was predicted to be around 18°C at 0.11 gH2O/gDW (achieved in representative samples after 30 min in a RH of 23%–52%) and processing beyond this level led to further DNA damage, additional samples intended for storage were processed for 30 min. During these experiments the RH varied between 20% and 46%. Dried samples were immediately sealed within Dri-shield bags (Fig. 3) together with a small volume of the ambient environment (i.e., the package was not evacuated nor filled with any other gas). Based on Karl-Fisher titration of parallel samples, in these experiments batches were dried to 0.1201±0.0076 gH2O/gDW with microwave-assisted drying (Time 0; Fig. 4). Moderate water loss continued during the 8-week storage period (Fig. 4). Specifically, the average moisture content gradually decreased (p<0.05) from 0.1201±0.0076 to 0.1027±0.0110 and 0.1066±0.0095 gH2O/gDW after 4 and 8 weeks of storage at 4°C, respectively. When stored at ambient temperature, the moisture content decreased (p<0.05) to 0.0677±0.0065 gH2O/gDW after 4 weeks, and to 0.0915±0.0085 gH2O/gDW after 8 weeks (Fig. 4). While the samples stored at ambient temperature appeared to lose moisture by 4 weeks and then regain some moisture at 8 weeks, this is likely an artifact of the experimental design. It is possible that the variation in moisture content that occurred during processing combined with the variation in ambient temperature resulted in the apparent trend in end moisture that was observed. This hypothesis would need validation in future studies.

Heat-sealed samples in Dri-shield bags.
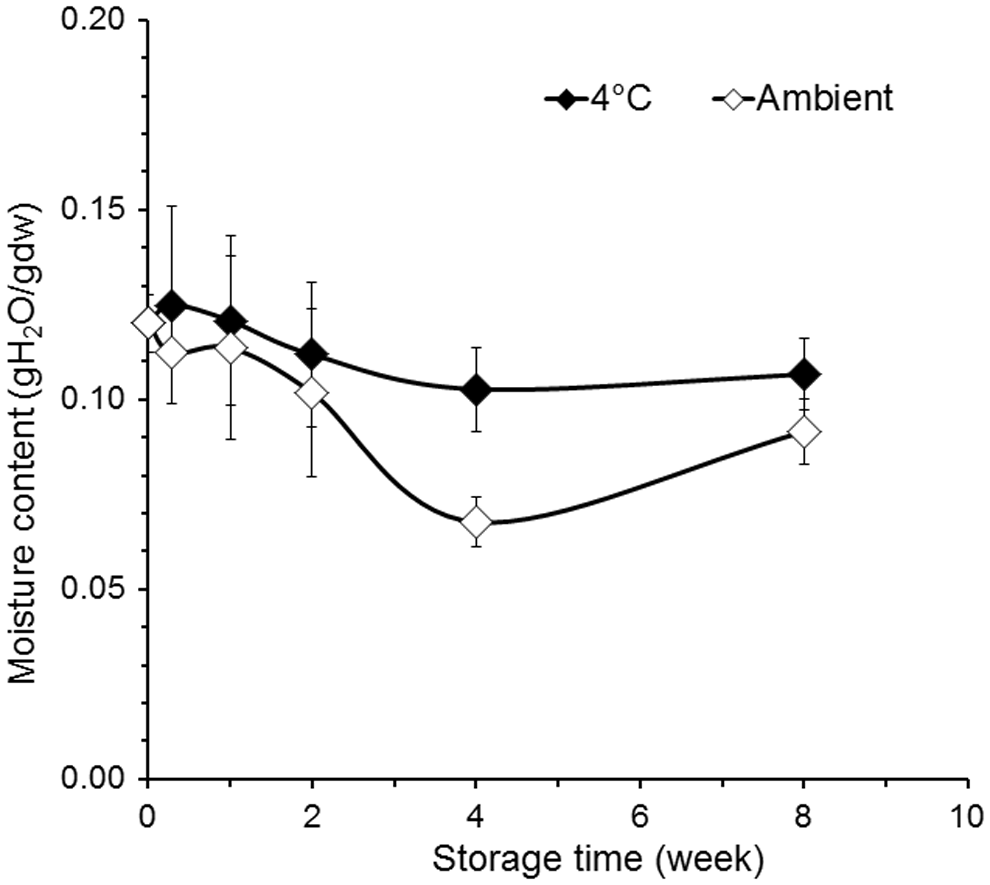
Moisture content as a function of storage time in Dri-shield bags for samples stored at 4°C and room temperature.
The highest proportion of empty oocytes was observed after storage at 8 weeks at ambient temperature, which was not statistically different (p>0.05) from the percentage of empty oocytes at 1 and 2 weeks at this same temperature (Table 2). Interestingly, the incidence of damaged GVs (TUNEL positive relative to the total number of oocytes with a GV) was unchanged (p>0.05) for samples stored at ambient conditions (Table 2). For both storage temperatures, the proportion of GV with DNA fragmentation more than doubled (p<0.05) from 0 to 2 days, then remained stable through 4 weeks (p>0.05; Table 2). The samples stored at ambient conditions for 8 weeks presented with less DNA damage. However, these samples had a higher initial loss of GV, so the overall percentage of intact GVs with no DNA damage was similar to the other storage points.
Percentage of oocyte without GV relative to the total number of oocytes; **percentage of TUNEL positive oocytes relative to the total number of oocytes with GV; £ percentage of oocytes with intact DNA relative to the total number of oocytes. Within columns, proportions with different superscripts differ (p<0.05, Chi-square).
Discussion
This study characterizes, for the first time, the incidence of DNA damage in GVs exposed to microwave-assisted dehydration—an alternative approach for preserving the maternal genome at cool or ambient temperature. In previous work the authors demonstrated the feasibility of using a standard household microwave to enhance drying of cells via the delivery of small pulses of energy. In general, evaporative cooling slows the evaporation rate, thus these pulses serve to increase the temperature of the sample modestly to overcome this slow-down in drying. 26 Modest thermal input can bring the temperature back up to near ambient in order to maintain a more consistent drying rate throughout the whole drying process.
In initial studies, to control thermal excursions above the desired physiologic range, samples were intermittently heated (manual start/stop) using a 600W conventional microwave. 26 In subsequent work, an analytical grade microwave was used to enable continuous heating at a lower power setting. 21 This system was the preferred choice for the current studies. This analytical grade unit uses the same 2.45 GHz magnetron as a conventional household microwave system, but the instrument is designed to enable more precise control over the power delivered. To the authors' best knowledge, microwave-assisted drying has not been used for drying DNA in the manner described in the current work (i.e., for the purposes of preservation). Extensive studies have been performed to elucidate the cell/DNA damage mechanism involved in the use of microwave energy for biocidal effects,27–29 yet this remains an area of active exploration. It is exceedingly difficult to uncouple thermal microwave injury from athermal microwave injury, as it is difficult to establish a control which has exactly the same thermal history at a local level, but is absent of microwave energy. Thermal histories created by conventional surface heating modes are not directly comparable.
In previous work we have attempted to review some of the literature in this area. 26 Our findings demonstrate the feasibility for successful microwave-assisted dehydration of the mammalian GV to reach end-moisture contents that are nonlethal, yet enable noncryogenic storage. In the current work we demonstrated that the moisture content of trehalose samples with GVs embedded could be reduced to close to 0.1 gH2O/gDW with 30-min microwave treatment without a significant increase in DNA fragmentation compared to samples that had not been dehydrated. At this moisture level, the glass transition temperature is expected to be in the vicinity of 18°C, thus facilitating noncryogenic storage.
Because 30 minutes of processing time enabled maximum water loss without DNA damage, this was chosen as the optimal processing condition in order to prepare samples for extended storage at both 4°C and at ambient temperature. When GVs were stored at these supra-zero temperatures, most DNA disruption occurred within the first 2 days and around two-thirds of GVs retained intact DNA, regardless of further water loss during storage. The degradation that occurred during the initial 2 days can possibly be attributed to the samples being moderately above the ideal moisture content for storage. Samples were sealed into bags at a moisture content of ∼0.12 gH2O/gDW, which was predicted to result in a Tg in the range of ∼10°C, based on a binary best fit curve to multiple data sets.20,21 Given the difficulty in measuring Tg in situ, and a lack of information regarding the optimal storage temperature with respect to Tg, the existence or non-existence of the glassy state in these samples is somewhat speculative. Storage in the Dri-shield bags resulted in moderate additional drying, as the bags themselves were found to absorb some amount of moisture. It is possible that this additional amount of drying may have pushed the samples from the near glassy regime into the glassy state, or possibly deeper into the glassy state (larger Tg-Tstorage), thus providing better protection against degradation pathways that are sensitive to molecular mobility. In recent work using dynamic mechanical analysis, we have shown that aqueous trehalose samples dried to 0.11 gH2O/gDW with microwave-assisted drying (11% RH) on glass fiber matrixes have a Tg≥room temperature, providing support for this hypothesis. 23
Although the samples stored at ambient temperature appeared to dry more than those stored at 4°C, no significant difference was observed on the extent of DNA damage, which would suggest that the samples were indeed in a glassy state at both storage conditions, and after the initial moisture loss during storage, the final moisture contents obtained were suitable for extended storage. In the initial processing studies, extending the microwave drying time beyond 30 minutes did not lead to substantial increases in moisture loss. This was expected because the equilibrium moisture content for amorphous trehalose in a 44% RH environment is ∼0.105 gH20/gDW. 24 In order to dry to lower moisture contents, a lower relative humidity would be necessary. By decreasing the ambient relative humidity to, for example 22%, samples would be expected to achieve the same moisture level that was attained by packaging the samples in the dessicating Dri-shield bags. This could potentially reduce the injury that accrued during the initial days of storage, when the molecular mobility in the sample was likely higher than the optimal level needed to suppress degradation reactions. Interestingly, processing for 40 min led to increased DNA damage (32.5% of GVs presenting with DNA breaks), with only a small amount of further water loss. The source of this damage is not known at this time. Because there is little water left to absorb the microwave energy and cause heating at extended processing times, thermal excursions are not likely to be causative. 18
It is possible that the decrease in water content, although very small, was the source of increased GV injury. It is noteworthy that for the samples that continued to dry during storage, the percentage of GVs with DNA damage was also observed to be as high as 35%, while GV loss also increased. It is possible that there exists a critical moisture level, beyond which some damage is to be expected. Even if this were the case, the fact that a significant fraction of the GVs were intact and had no evidence of DNA damage after nonrefrigerated storage for 8 weeks is a very promising indicator of the viability of this technique.
The timing of tissue availability for these studies coincided with adverse environmental conditions, thus the environmental control during the experimental phase of this work was constrained and non-ideal (23%–52% RH). Although the nominal drying rates within this environmental humidity range were similar, the end moisture contents were not as precisely prescribed as would have been possible in more stable relative humidity conditions. Because the Tg increases rapidly as the moisture content decreases in this low moisture content regime, variability in the data is possibly associated with these modest differences in end moisture content due to non-stable humidity. Despite these challenges, the moisture content achieved at the end of microwave-assisted drying was close to the desired end-point and statistically-based conclusions could be made. In future studies, improved humidity control would enable the end moisture content to be prescribed more precisely and the processing conditions further tuned to enable samples to be dried to an optimal moisture level for the desired storage goal.
In the current study, a very basic solution composition was used, without any attempt to optimize the components. Trehalose was chosen as the glass-former because it is known to provide protection to biomolecules while yielding a glassy state at high temperatures (relative to ambient). A 1.5 M trehalose solution was used to enable a reduction in the overall processing time. While some investigators prefer to use isotonic solutions, 25 in other work we have shown that starting from a higher concentration can be beneficial as it reduces the overall osmotic stress. 19 It is recognized that this approach may or may not be beneficial in the current case because the membrane is permeabilized to enable trehalose loading into the oocyte. Volume changes due to osmotic excursions are thus not expected to be a contributor to injury.
Effects of chemical excursions on the GV are currently unknown. In the absence of such data, the shortest route to a dried specimen was chosen to evaluate the feasibility of anhydrous preservation of the GV. 1X TE buffer was used in the formulation, which contains 1 mM EDTA and 10 mM Tris. EDTA is a very common additive in DNA handling and preservation media, providing DNA protection by chelating free metal ions (such as magnesium) that are required by DNAases.26,27 However, the existence of a beneficial effect was not evaluated in the current study. There exists considerable room for formulation optimization going forward.
The microwave power level used in this study (20% power, nominally 120 W) was chosen because it was unlikely to cause thermal excursions much beyond ambient conditions. 18 It is possible that by increasing the power it may be possible to further reduce the processing time to achieve a glassy state within a shorter window. This could potentially lead to a reduction in the chemical stress and an overall improvement in outcome. Better thermal feedback during microwave processing would be necessary to enable such processing developments. Because of the small sample size, high resolution in situ methods of thermal sensing would be necessary to enable true sample temperatures to be resolved.
This study characterized, for the first time, the metrics and kinetics of moisture removal, and the DNA damage related to GV desiccation as an alternative approach for preserving the maternal genome at cool or ambient temperature. Because of the high level of preserved DNA integrity, we do not expect issues with functionality after injection into a recipient cytoplast (currently evaluated in ongoing studies). The impact of microwave-assisted drying on the epigenetic patterns of GVs will have to be measured in order to validate the approach. Fortunately, preliminary studies in the cat GVs reveal that the epigenome is highly stable regardless of the nonphysiological conditions (data not shown). Ambient preservation provides an easier and more cost-effective approach for biobanking of genetic resources. Given the generous space for optimization of the process and composition, these initial studies successfully demonstrated the potential for dry storage of GVs. With recent advances in our ability to measure Tg in situ, 28 and with better control of local humidity, we expect to be able to refine these processes, while maximizing recovery of viable GVs.
Footnotes
Acknowledgments
This work was supported in part by National Center for Research Resources (R01 RR026064), a component of the National Institutes of Health (NIH), and currently supported by the Office of Research Infrastructure Programs/Office of the Director (R01 OD010948), as well as by the National Institute of General Medical Sciences (R01 GM101796).
Author Disclosure Statement
No competing financial interests exist. Gloria Elliott is the inventor on a patent for microwave drying of biologics.