
Abstract
The aim of this study was to evaluate the effects of Kit Ligand (KL) on the growth of vitrified follicle, oocyte quality, and embryo development in two- and three-dimensional culture systems. Vitrified and nonvitrified mouse whole ovaries were cultured for 1 week, then their isolated preantral follicles were cultured for 12 days in two- or three-dimensional culture systems in the presence and absence of KL. The growth and diameter of follicles, maturation of oocytes, and hormonal level were assessed. Finally, embryo developmental rate and oocytes reactive oxygen species (ROS) levels, adenosine triphosphate (ATP) content, and distribution of mitochondria were examined. The results showed the maturation and survival rates and steroidogenesis of follicles were significantly higher not only in the three-dimensional culture system but also in the presence of KL (p < 0.001). The embryo development, ATP content, and ROS level of collected oocytes were not significantly different in all groups. Mitochondrial distribution within the oocyte was similar in all groups of the study. Our results demonstrated that KL supplementation during follicular culture in two- and three-dimensional culture systems appeared to significantly increase follicular function and development. The ovarian vitrification had no harmful effect on the steroidogenesis, growth, and maturation of follicles.
Introduction
I
Several culture systems have been developed to support preantral follicular growth, including two- and three-dimensional culture systems.7–11 In the two-dimensional culture system, the isolated follicles are attached to the surface of culture dishes and the granulosa cells show outgrowth onto the plate.12,13 In the three-dimensional culture system, follicles do not attach to the surface of the culture dish, and therefore, their native morphology is preserved and could mimic the stromal microenvironment of the ovary, to support the somatic cell and oocyte interactions for optimization of oocyte development. Some extracellular supports such as sodium alginate, agarose, and Matrigel are used for this purpose.14–17
Follicular development is a complex process that is regulated by locally produced factors such as Kit Ligand (KL). KL is known as steel factor, stem cell factor, or mast cell growth factor and is produced by granulosa cells. It is an important regulator of primordial follicle development and it acts on oocytes, theca, and stromal cells. 18 Several studies have demonstrated that KL is involved in the survival and activation of primordial germ cells, proliferation of granulosa cells, differentiation of theca cells, and cytoplasmic maturation of oocytes.19–23 We have recently showed that KL could improve the development of follicles in vitrified and nonvitrified cultured ovaries, and also could act as antiapoptotic factor during ovarian culture. 16
One of the major challenges regarding in vitro follicular growth is the low rate of oocyte maturation and embryo development when compared to in vivo samples. 24 These effects are more prominent in cryopreserved samples.25–28 Moreover, there are controversial reports regarding the effects of vitrification on the developmental potential of ovarian follicles after in vitro maturation.29–32 Haidari et al. observed that the growth and maturation of cultured preantral follicles derived from vitrified–warmed mouse ovarian tissue were similar to that of fresh ovaries. 30 In addition, Segino et al. demonstrated that vitrification of mouse ovarian tissue did not affect the oocyte's ability to undergo meiosis. 31 In contrast, Luvoni et al. reported that oocytes derived from in vitro matured vitrified feline ovarian fragments had a higher proportion of gametes arrested at the germinal vesicle (GV) stage compared to those isolated from fresh tissue. 25 Trapphoff et al. have shown a decrease in the development of vitrified preantral follicles following 12 days of vitro culture. 32
One suggestion regarding the low developmental rate of in vitro matured follicles is that it's due to oocytes mitochondrial dysfunction, which results in low adenosine triphosphate (ATP) production, oxidative damage, and apoptosis.33–36 The reactive oxygen species (ROS) production is an additional factor that may be involved in oocyte meiotic arrest.37,38
According to our knowledge, there is little information regarding the improvement of follicular development derived from vitrified whole ovaries by supplementation of culture media with KL. Moreover, the comparison of oocyte quality derived from isolated follicles culture in two- and three-dimensional culture systems has not been documented. Therefore, the aim of this study was as follows: first, to evaluate the development and function of cultured follicles derived from vitrified neonatal mouse ovaries in the presence and absence of KL by assessment of follicular diameter, hormonal production, antrum formation, and oocyte maturation; second, to evaluate the oocytes quality by studying their fertilization and embryo development, ROS levels, ATP content, and distribution of mitochondria.
Material and Methods
Unless otherwise stated, all the chemicals and reagents used for this study were purchased from Sigma-Aldrich.
Animals
National Medical Research Institute (NMRI) outbred mice were used in this study and housed in the animal house of the university with free access to food and water in a temperature and light controlled environment. Approval for this study was obtained from the Animal Research Ethical Committee of the Tarbiat Modares University (Ref No. 52/99424). The ovaries were obtained from 1-week-old female mice (n = 50). The sperms were prepared from the cauda epididymis of 10–12-week-old NMRI male mice (n = 10).
Experimental design
The ovaries of 1-week-old female mice were collected and pooled (n = 50), then they were divided into vitrified and nonvitrified groups. The whole ovaries from both groups were cultured for 7 days. Then, their preantral follicles were mechanically isolated and cultured for 2 weeks more in both two- and three-dimensional culture systems, in the presence and absence of KL. The isolated follicles were cultured in the following groups: nonvitrified KL− 2D, nonvitrified KL+ 2D, nonvitrified KL− 3D, nonvitrified KL+ 3D, vitrified KL− 2D, vitrified KL+ 2D, vitrified KL− 3D, and vitrified KL+ 3D. The diameter and survival of follicles were recorded, and hormonal analyses were performed.
The released metaphase II (MII) oocytes in each subgroup were collected and randomly considered for ATP content analysis, mitochondrial distribution, ROS assay, and the study of the embryo development to the blastocyst stage.
Vitrification and warming
The whole ovaries were vitrified and warmed based on the previously described method 39 with a modification in the carrier system. Vitrification solution was made of PBI medium containing 30% (w/v) Ficoll 70, 0.5 M sucrose, and 40% (v/v) ethylene glycol. The ovaries were transferred to a vitrification solution for 5 minutes at room temperature. After dehydration, they were individually put on to the inner surface of the Cryolock with a minimal volume of the vitrification solution, quickly immersed into liquid nitrogen, and stored for 1 week. For warming, the Cryolocks containing the ovaries were sequentially dipped into 1 mL of 1, 0.5, and 0.25 M sucrose solutions for 5 minutes in each step at room temperature. The recovered ovaries were transferred and incubated in α-MEM medium supplemented with 10% fetal bovine serum (FBS) under mineral oil at 37°C in a humidified 5% CO2 in air for 5 minutes.
Whole ovary organ culture
Nonvitrified and vitrified whole ovaries (n = 50 for each group) were cultured individually on 24-well plates with tissue culture well inserts (Millicell-CM, 0.4-μm pore size; Millipore Corp.). The culture media were α-MEM medium supplemented with 5% FBS, 1% insulin, transferrin, and selenium (ITS; Gibco), 100 mIU/mL recombinant follicle stimulating hormone (rFSH or Gonal-f; Serono), and 100 ng/mL KL. Approximately 400 μL of culture media was added to the compartment below the membrane insert. The ovaries were cultured at 37°C, 5% CO2, for 7 days. Every other day, 200 μL of media was replaced with fresh culture media. 16
Follicle isolation
After 1 week of whole ovarian culture, the preantral follicles with 140–150 μm size were mechanically isolated from both vitrified and nonvitrified ovaries using 29-gauge needles attached to 1-mL syringes under a stereomicroscope. The follicles containing two or three layers of granulosa cells, with centrally located normal oocytes, and a thin layer of theca were selected and considered for two- and three-dimensional in vitro follicular culture for two additional weeks.
Two- and three-dimensional in vitro culture of isolated preantral follicles
The isolated preantral follicles derived from both vitrified (n = 2722) and nonvitrified (n = 2566) cultured ovaries were cultured in two- and three-dimensional systems for 2 weeks. Uncapsulated isolated preantral follicles were individually cultured in 35 μL drop of culture media under mineral oil in a two-dimensional culture system.
The preantral follicle was encapsulated in sodium alginate using the method described previously in the three-dimensional culture system. 40 Briefly, sodium alginate containing 55%–65% glucuronic acid was dissolved in deionized water at a concentration of 1% (w/v) and mixed with activated charcoal (0.5 g of charcoal per gram of sodium alginate). Before each encapsulation, the prepared sodium alginate solution was diluted with sterile phosphate buffer saline (PBS) at a concentration of 0.5% (w/v) at room temperature. The isolated follicles were individually transferred inside droplets of sodium alginate (5 μL), then they were slowly released into a cross-linking solution (50 mM CaCl2 and 140 mM NaCl) for 2 minutes, then alginate beads were removed and rinsed in α-MEM media.
The uncapsulated and encapsulated follicles were cultured in α-MEM medium supplemented with 5% FBS, 100 mIU/mL recombinant FSH, 1% ITS, 20 ng/mL murine recombinant epidermal growth factor (rEGF), 100 mg/mL penicillin, and 50 ng/mL streptomycin in the presence and absence (n = 120 for each subgroups) of 50 ng/mL of KL under mineral oil at 37°C with 5% CO2 for 12 days. Every other day, half of the media were replaced with fresh culture media and the collected media were separately frozen at −80°C until hormonal assays were performed.
In vitro ovulation induction
For recovery of follicles in beads, 5 mM sterile ethylene glycol tetra acetic acid was added directly to the medium and incubated at 37°C for 5 minutes, until the beads dissolved. The ovulation was induced by adding 1.5 IU/mL human chorionic gonadotropin (Organon, Griekenweg) to the culture media 12 days after follicular culturing. The released oocytes were scored as GV, GV breakdown (GVBD), and MII.
Assessment of follicular morphology and diameter
The follicular morphology was checked under an inverted microscope every 48 hours during the culturing period. The follicles were considered as degenerated when the oocyte and the granulosa cells had become dark and fragmented or the diameter of the follicle was decreased.
Measurement of follicular diameter was done during cultivation time (n = 15 for each subgroup) using a precalibrated ocular micrometer at 100X magnification under an inverted microscope.
Hormonal assays
The levels of 17-β estradiol (E2), progesterone (P4), and dehydroepiandrosterone (DHEA) were measured in the collected media from all subgroups (n = 5 for each subgroup) at the end of the culture period. The levels of E2 were measured by a Microplate Enzyme Immunoassay kit (Monobind, sensitivity = 6.5 pg/mL), P4 by an enzyme-linked immunosorbent assay kit (DIAPLUS, sensitivity = 0.1 ng/mL), and DHEA by a Microplate Enzyme Immunoassay kit (Monobind, sensitivity = 0.04 μg/mL).
In vitro fertilization and embryo culture
The MII oocytes were collected from each subgroup and subjected to in vitro fertilization. The cauda epididymis was dissected from 7- to 8-week-old male NMRI mice (n = 10) and placed into a 500-μL drop global® medium (global total®; LifeGlobal) supplemented with 5 mg/mL bovine serum albumin (BSA) under mineral oil at 37°C in a 5% CO2 incubator for 1.5 hours. The MII oocytes were transferred to insemination media, which consisted of global medium supplemented with 15 mg/mL BSA, with capacitated spermatozoa, for 4–6 hours. Then, the oocytes were transferred to 20 μL drops of global medium with 5 mg/mL BSA under mineral oil. The rates of fertilization, for 2, 4, 8 cell, morula, and hatching blastocyst embryos, were assessed 10, 24, 48, 72, 96, and 120 hours later, respectively.
ROS assay
To measure ROS levels in MII oocytes from all in vitro culture groups (n = 60 for each subgroup for three repeats), 20 MII oocytes were pooled and washed three times with PBS. Then, they were incubated in 40 mmol/L of Tris–HCl buffer (pH = 7.0) containing 5 μmol/L 2′, 7′ dichlorodihydrofluorescein diacetate (Merck) at 37°C for 30 minutes. They were washed with PBS, sonicated at 50 W for 2 minutes, centrifuged at 4°C and 10,000 g for 20 minutes, and the supernatant monitored using a spectrofluorometer at 488 nm excitation and at 525 nm emissions. Data were expressed as μM H2O2 and the mean dichlorofluorescein fluorescence intensity (mean ± standard deviation [SD]). These experiments were repeated at least three times. 8
Measurement of oocyte ATP content
The ATP content in the cytoplasm of each MII oocyte (n = 10 in each subgroup) was measured using methods described previously. 36 Briefly, the MII oocytes were individually frozen at −80°C in 20 μL of ultrapure water. ATP levels were quantified by measuring the luminescence (Berthold LB 9501 luminometer) generated in an ATP-dependent luciferin–luciferase bioluminescence assay (Bioluminescence Somatic Cell Assay System; Sigma). A standard curve containing different ATP concentrations was analyzed for each series of analyses. The ATP content was calculated from the formula for the standard curve (linear regression).
Staining of mitochondria
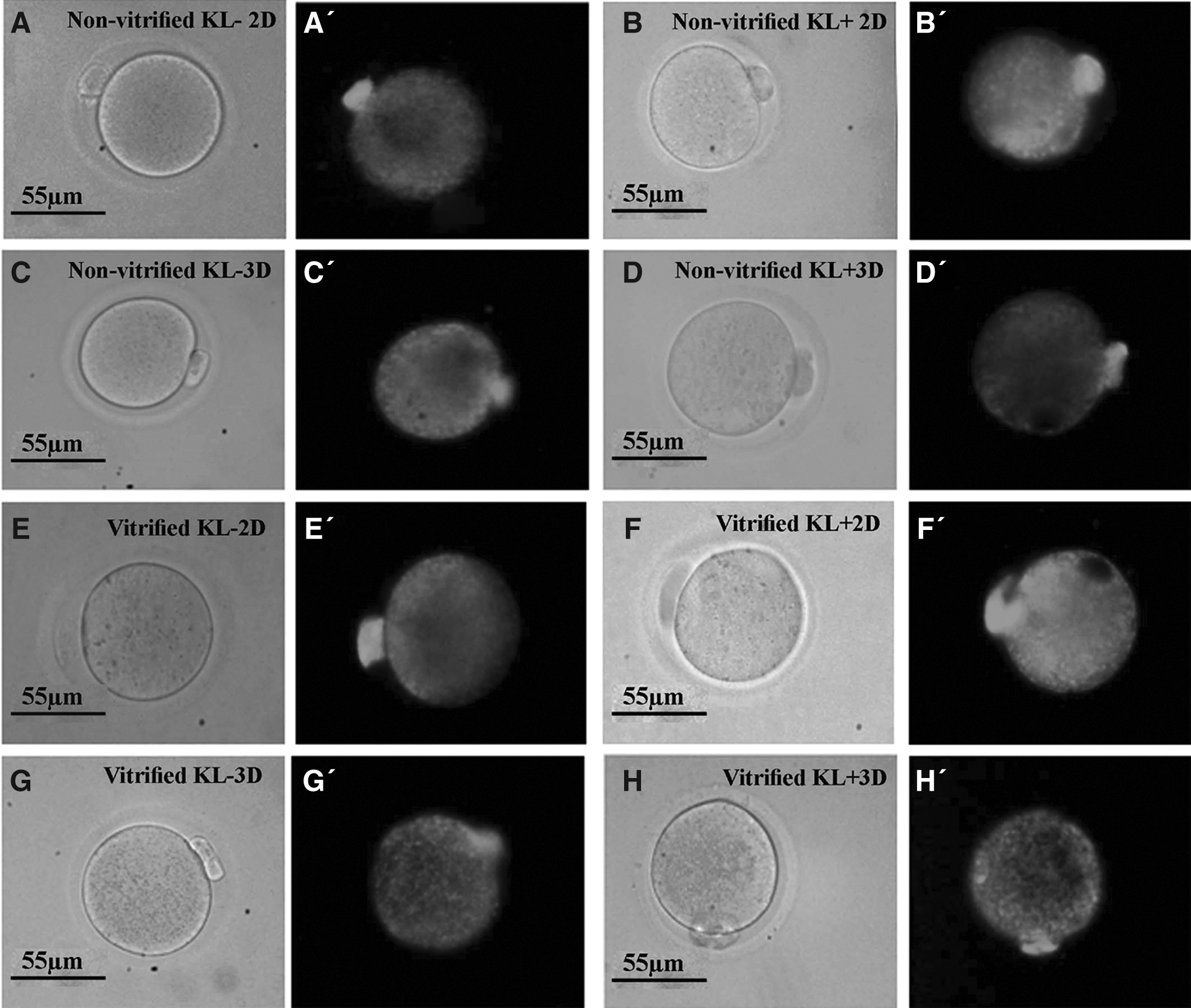
The MII oocytes of all groups were stained by MitoTracker green (Molecular Probes; Invitrogen). The stock solution of 1 mmol MitoTracker green in dimethyl sulfoxide was prepared and stored at −20°C. The MII oocytes (n = 5 in each subgroup) were stained for 12 minutes with MitoTracker Green at a concentration of 0.2 mmol in PBS at 37°C. Then MII oocytes were washed in PBS and visualized using a fluorescent microscope at 490 wavelength. 41
Statistical analysis
The results are expressed as the mean ± SD. The survival and developmental rates of cultured follicles and embryos in all groups of study were assessed by one-way ANOVA, and Tukey's HSD was used for post hoc tests. ROS and ATP levels were analyzed statistically by the t-test. Statistical significance was defined as p < 0.05.
Results
The diameter of cultured isolated preantral follicles
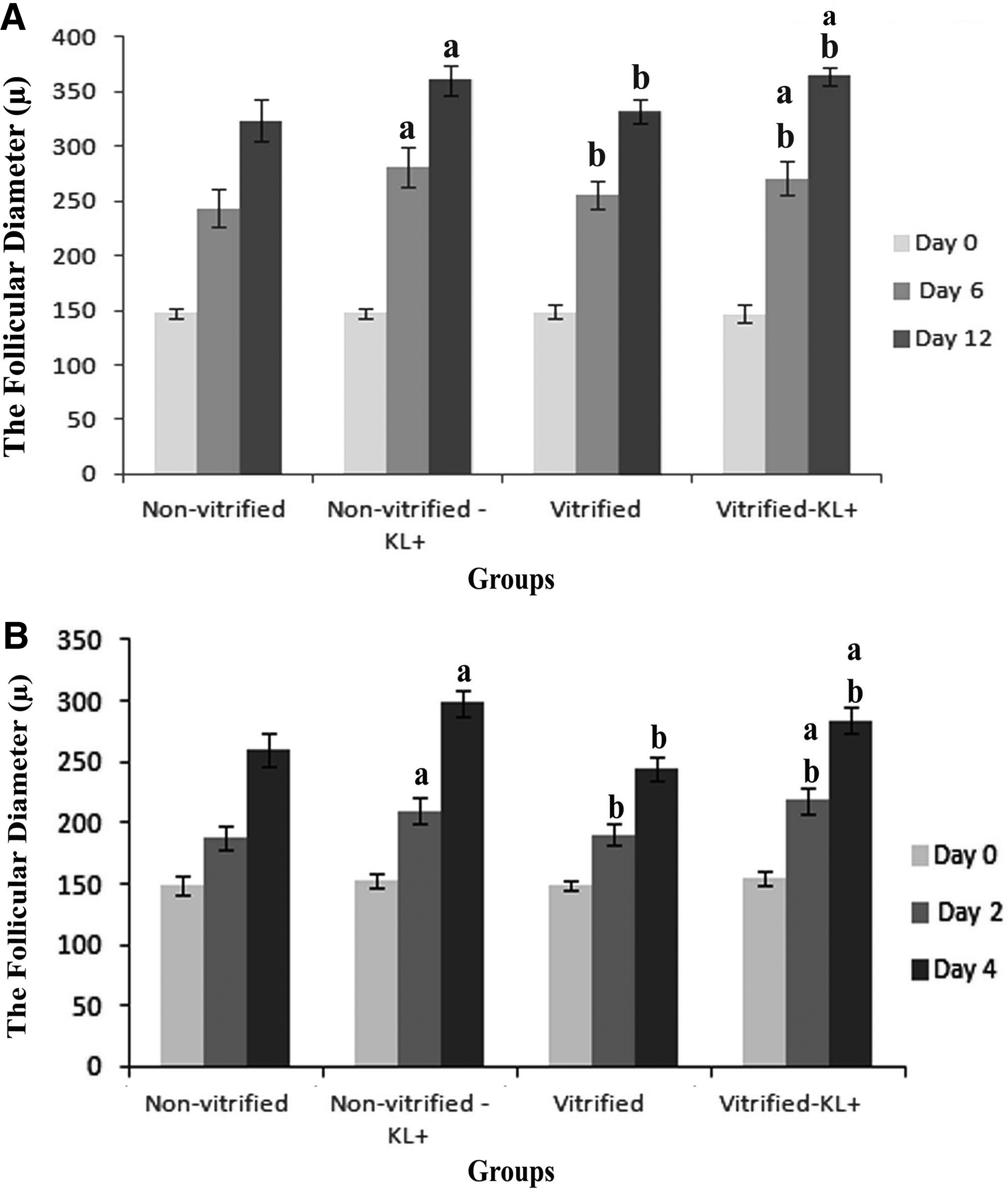
The phase contrast micrograph of the cultured isolated follicles related to both vitrified and nonvitrified groups in two-dimensional and three-dimensional culture systems is demonstrated in Figures 1 and 2, respectively. The cultured follicles in the two-dimensional culture system were attached to the floor of plates and had irregular outlines, thus their follicular measurements were performed up to the fourth day of culture. As Figure 3 shows, the size of follicles in all groups of study was increased during the culture period.

The inverted microscope images of two-dimensional in vitro follicular development on day 0 (first row), day 6 (second row), and day 12 (third row) in nonvitrified KL−
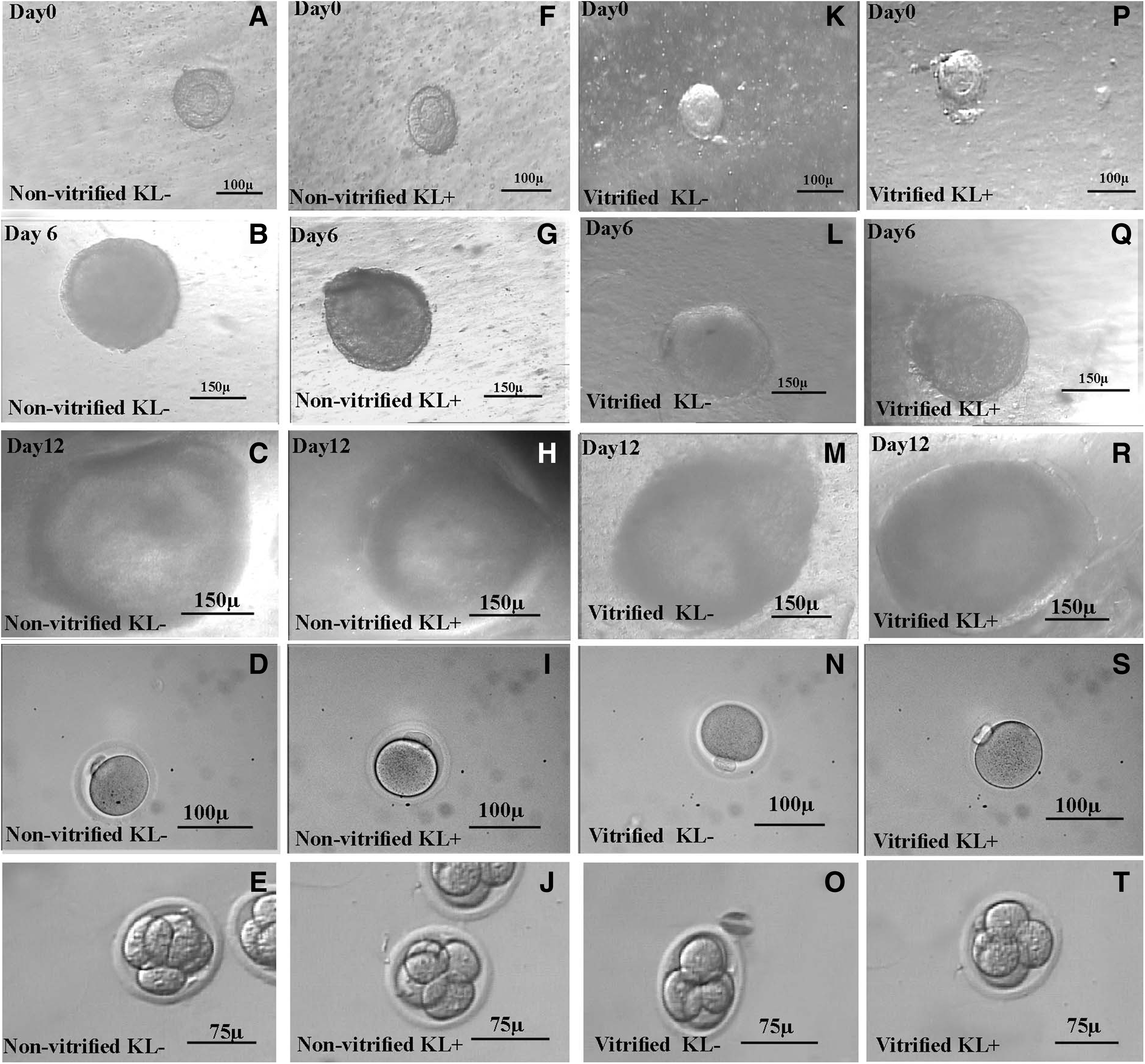
The inverted microscope images of three-dimensional in vitro follicular development in alginate on day 0 (first row), day 6 (second row), and day 12 (third row) in nonvitrified KL−
The diameter of preantral follicles (μm) during the two-dimensional
The diameter of follicles at the beginning of culture was 150 ± 3 μ for all groups under study, however, the size of cultured follicles in the two-dimensional culture system on day 4, and in the three-dimensional culture system on day 12, was significantly increased in all KL-treated groups compared to non-KL-treated groups (p < 0.001). However, there was no significant difference between vitrified and nonvitrified groups (Fig. 3).
Follicular developmental rate
The developmental rate of follicles in all groups of study is summarized in Table 1. There were significant differences regarding the survival rates of follicles at the end of the culture period between KL-treated groups and non-KL-treated groups in both two- and three-dimensional culture systems. A higher percentage of survival rate was seen in the three-dimensional culture system than two-dimensional culture groups (p < 0.001), and there was not a significant difference between vitrified and nonvitrified groups.
Significant differences with respect to non-KL-treated group in the same column (p < 0.005).
Significant differences with respect to 2D group in the same column (p < 0.001).
2D, two-dimensional culture system; 3D, three-dimensional culture system; KL, Kit Ligand; MII, metaphase II.
In nonvitrified groups, the higher MII rate was achieved in KL-treated samples (p < 0.001). Two-dimensional culture groups had a significantly lower MII rate than three-dimensional groups (p < 0.001). There was no significant difference between vitrified and nonvitrified groups in this regard.
Fertilization rate and embryo development
The representative figures of MII oocytes and 4 cell embryos derived from all groups of study are shown in Figures 1 and 2. The fertilization rates of MII oocytes collected from two- and three-dimensional culture systems are summarized in Table 2. The higher percentages of fertilization and embryo development were seen in three-dimensional culture systems compared to two-dimensional culture systems (p < 0.001). There was no significant difference between vitrified and nonvitrified groups.
Significant differences with respect to non-KL-treated group in the same column (p < 0.005).
Significant differences with respect to 2D group in the same column (p < 0.001).
Hormonal assays
The concentrations of E2, P4, and DHEA in collected media at the end of follicular culture period are compared and summarized in Table 3 for all groups of the study. The hormonal production was significantly higher in follicles that were cultured in the presence of KL in the three-dimensional culture system compared to other groups (p < 0.05), and the levels of these hormones were significantly higher in the three-dimensional culture group compared to the two-dimensional culture group (p < 0.001). There was no significant difference between vitrified and nonvitrified groups in this regard.
Significant differences with respect to non-KL-treated group in the same column (p < 0.005).
Significant differences with respect to 2D group in the same column (p < 0.001).
Significant differences with respect to nonvitrified group (p < 0.01).
DHEA, dehydroepiandrosterone; SD, standard deviation.
ROS level
The ROS levels in MII oocytes collected from all groups of study are given in Table 4. Data are shown as μM H2O2, and there was no significant difference between the ROS levels in all cultured groups.
There was no significant difference between groups.
ATP, adenosine triphosphate; ROS, reactive oxygen species.
The ATP content
The ATP level of the individual oocytes was measured by luciferin–luciferase reaction, and these levels are summarized in Table 4 as pmol per each MII oocyte. The ATP concentration of oocytes was not significantly different in all cultured groups.
Distribution of mitochondria
Mitochondrial distribution in the cytoplasm of MII oocytes in all cultured groups showed a similar pattern, and the mitochondria mainly diffused within the cytoplasm homogenously (Fig. 4).
The phase contrast micrograph of in vitro MII oocytes collected from nonvitrified KL− 2D
Discussion
Different follicular culture systems have been introduced by several investigators, which provide a milieu that mimics the in vivo ovarian environment.42,43 However, successful oocyte maturation, fertilization, and embryo development depend on culture systems and media and research initiatives focused on culture improvement.43–45
In the present study, we achieved the highest developmental capacity of follicles, MII oocyte rate, fertilization rate, embryo developmental rate, and level of hormone production in the three-dimensional follicular culture system compared to the two culture system.
These data support the idea that capsulated follicle with alginate could maintain the spherical morphology of the preantral follicle similar to in vitro basal lamina architecture. It seems that this three-dimensional culture system could better preserve the communication between the oocyte and the granulosa cells that is essential for sharing of paracrine factors that promote the growth of both cell types during the follicular maturation process. However, our observations in the two-dimensional culture system suggest that the cytoarchitecture of follicles may change and alter the paracrine signaling between the follicular cells and oocytes. Gomes et al. showed that the traditional two-dimensional culture system resulted in a distortion of follicle morphology, marked extracellular matrix modifications, and high rates of follicle disruption. Moreover, the three-dimensional culture system can maintain follicular structure with an in vivo-like basal lamina architecture that minimizes the disruption of follicles. 46 It was demonstrated that three-dimensional culture of preantral follicles using an extracellular matrix provided a better milieu for in vitro growth and survival of follicles when compared with the conventional two-dimensional culture system.47,48
We propose that high production of steroid hormone (E2, P4, and DHEA) by follicles grown in the three-dimensional culture system may be due to a high level of androgen provided by theca cells, and accelerated increased aromatase activity, and increased granulosa cell numbers.
In other parts of this study, we showed that the follicles that were cultured in KL-supplemented media also had high rates of follicular survival, development, and steroid hormone production, especially in the alginate encapsulated group. The positive effect of KL on follicular growth and development, which was observed in the present study, is in agreement with similar previous reports.49,23 Moreover, the positive effect of KL as an antiapoptotic factor on improvement of follicular development of cultured vitrified whole ovaries was shown by our recent study. 16 KL acts directly through its receptor on the granulosa and theca cells and oocytes, as a proliferative factor. Indirectly, KL induces other functional receptors on follicular cells such as follicle-stimulating hormone and the fibroblast growth factors receptor, which has a role in the process of follicular development. 50
Moreover, data from the present study showed for the first time that the supplementation of follicular culture media with KL was more effective in the three-dimensional culture system than the two-dimensional culture system. It is suggested that intercellular communication and receptors were well preserved in the three-dimensional culture system, similar to the in vivo normal conditions. Thus, KL may activate the phosphoinositide-3 kinase pathway in the oocytes through the Kit receptor, and then it acts on granulosa cells. However, more study is needed to prove this suggestion.
The developmental and maturation rate of follicles and oocytes was similar in vitrified and nonvitrified groups. Therefore, our results reconfirmed the potential of the vitrification method using ethylene glycol for long-term storage of ovarian tissue, with a minimal harmful effect on the developmental capacity of mouse ovarian follicles.
Our results for the first time demonstrated that no significant difference was seen in all groups in the study regarding embryo development, ATP content, mitochondrial distribution, and the ROS level of oocytes. These results may be because the healthy and normal follicles were selected for further in vitro culture, thus, the yield of MII oocytes was not different. Another point of view concerning technical limitations is that it is difficult to detect differences between the ATP content and ROS level of individual oocytes in different groups of study.
In conclusion, our results demonstrated that KL supplementation during follicular culture in two- and three-dimensional culture systems appeared to significantly increase follicular function and development. The ovarian vitrification had no harmful effect on the steroidogenesis, growth, and maturation of follicles.
Footnotes
Acknowledgments
The authors thank Mr. Pour Beyranvand for providing excellent technical assistance.
This work was supported by Tarbiat Modares University for the PhD thesis and Iran National Science Foundation.
Author Disclosure Statement
No conflicting financial interests exist.