
Abstract
In this study, the protective effects of monosaccharides (glucose and fructose) and sugar alcohols (mannitol, sorbitol, and xylitol) on frozen ram spermatozoa were evaluated and compared. The motility, moving velocity, and hypoosmotic swelling capability of spermatozoa frozen with monosaccharide or sugar alcohol were measured using a computer-assisted spermatozoa analyzer system. The acrosome status, membrane integrity, distribution of phosphatidylserine (PS), and mitochondrial membrane potential (MMP) were analyzed using fluorescence staining and flow cytometry. The results indicated that similar to glucose or fructose, the presence of sugar alcohol in the freezing extender cannot significantly improve the motility and moving velocity of ram spermatozoa equilibrated at 5°C. In terms of motility, pathway velocity, curve velocity, hypoosmotic swelling capability, acrosome and membrane integrity, and MMP, the inclusion of mannitol or sorbitol in the extender can significantly improve the quality of frozen–thawed ram spermatozoa compared to glucose or fructose. However, the effects of mannitol or sorbitol on linear velocity and PS distribution of frozen–thawed spermatozoa were similar to those of the monosaccharides (p > 0.05). In addition, the ability of xylitol to protect acrosome and maintain MMP in frozen–thawed spermatozoa was significantly higher compared with glucose or fructose (p < 0.05), although it could not improve the other evaluated parameters. Finally, there is no significant difference existing between mannitol and sorbitol with respect to the above evaluated parameters. In conclusion, the replacement of glucose or fructose by mannitol or sorbitol in a freezing extender can improve the postthaw quality of ram spermatozoa under specific freezing conditions. Moreover, the protective effects of mannitol and sorbitol on frozen–thawed ram spermatozoa are superior to that of xylitol. However, in the presence of sugar alcohols, the cryoinjury on spermatozoa membrane is still serious. In the future, the question of protecting the membrane of frozen–thawed spermatozoa needs further research.
Introduction
S
During freezing and thawing, mammalian spermatozoa face various kinds of stresses, such as ice formation, cold shock, chemical toxicity, osmotic stress, oxidative stress, apoptosis, and so on.10,11 These stresses mainly influence spermatozoa membrane and consequently result in a lower viability and fertility.10–12 The membranes of ram spermatozoa have a particular lipid composition, which makes them more sensitive to cryoinjury compared to other livestock species.3,13
Currently, low pregnancy rates can be achieved when cervical artificial insemination (AI) with frozen–thawed semen is performed. 14 However, as an alternative method, intrauterine AI with frozen–thawed semen can generate better fertility results compared to cervical AI, especially when a lower spermatozoa number is needed for intrauterine AI than for cervical AI.8,10 However, this method needs a specialized instrument and qualified technicians.15,16
As important cryoprotectants, sugars or sugar alcohols have been used for cryopreservation of mammalian spermatozoa.6,17,18 However, it must be noted that some sugars and sugar alcohols are naturally present in seminal plasma in varying concentrations.17,19 Besides fructose, as an important sugar alcohol, sorbitol is present in the seminal plasma of rams or bulls.17,20 Sorbitol can be produced during the conversion of blood glucose to fructose in the ram. 17 In addition, other kinds of sugar alcohols, such as inositol, 21 mannitol, erythritol, and glycerol, 19 have been identified in seminal plasma of livestock species.
During freezing, the osmotic difference between inner membrane and outer membrane benefits cell dehydration and subsequently reduces or even prevents lethal intracellular ice formation.6,8 However, cell dehydration–induced osmotic stress is also one of the major factors causing cellular injury during cryopreservation.22–25 Spermatozoa have been shown to be particularly susceptible to a hypertonic environment; even a slight increase in osmotic stress can cause an irreversible loss of spermatozoa motility.4,26
According to some reports, sugars or sugar alcohols can mitigate the negative effects of osmotic shock. 27 The effects of monosaccharides on cryosurvival of mammalian spermatozoa have been extensively investigated.6,17,18 Besides acting as primary substrates for glycolysis, monosaccharides can also increase the tolerance of mammalian spermatozoa to osmotic stress and prevent lethal intracellular ice formation, especially in the presence of penetrating cryoprotectants, such as glycerol.3,18,28 Currently, glucose and fructose have been frequently used as supplemental freezing extenders during cryopreservation of livestock semen.2,18,29 For example, the addition of 1.25% fructose to the citrate–egg yolk–glycerol-based extender can significantly increase the nonreturn percentage at 3 months from 56.9% to 66%. 30
Compared to fructose, the addition of glucose in the freezing extender can improve the viability of frozen–thawed stallion spermatozoa. 18 In addition, other monosaccharides can also be used for cryopreservation of mammalian spermatozoa. Arabinose can significantly improve the motility of bull spermatozoa cryopreserved at −79°C compared to fructose. 31 It has been reported that the addition of xylose in either egg yolk–citrate- or milk-based extenders can improve the viability of frozen–thawed spermatozoa. 32
There are some previous investigations supporting the beneficial effects of sugar alcohols on frozen–thawed spermatozoa,17,18,33 which may be due to their structural similarity to glycerol. 33 For example, as an important sugar alcohol, the cryoprotective effects of sorbitol have attracted some investigators' attention.17,18,34 Sorbitol can fuel mouse spermatozoa motility and protein tyrosine phosphorylation by sorbitol dehydrogenase. 35 Sorbitol can also increase the tolerance of plasma membranes to osmotic shock. 36 In addition, sorbitol has been used to improve cryosurvival of spermatozoa collected from rams, 33 stallions, 18 bulls,17,37 and men. 34 It has been found that the addition of sorbitol in an egg yolk–citrate-based freezing extender can provide certain cryoprotection for bull spermatozoa. 37
A freezing extender containing sorbitol has been used for successful cryopreservation of stallion spermatozoa. Moreover, the quality of frozen–thawed stallion spermatozoa was better compared with spermatozoa frozen in the presence of glucose or fructose. 18 Except for sorbitol, the effects of other sugar alcohols on frozen–thawed spermatozoa have also been evaluated in some previous investigations.37–39 For example, adonitol, erythritol, and xylitol have been used for cryopreservation of bull37,38 and boar spermatozoa. 39 In an investigation carried out by Molinia et al., sugar alcohols, such as mannitol, xylitol, and adonitol, may have some cryoprotective effects on frozen–thawed ram spermatozoa; this is not the case for inositol. 33
According to our knowledge, the investigations about the cryoprotective role of sugar alcohols on frozen–thawed ram spermatozoa are relatively few and need further evaluation. To determine whether sugar alcohols can improve the cryosurvival of ram spermatozoa, the effects of monosaccharides (glucose or fructose) and sugar alcohols (mannitol, sorbitol, or xylitol) on the postthaw viability of ram spermatozoa were evaluated and compared in this study. The motility, moving velocity, acrosome status, hypoosmotic swelling capability, mitochondrial membrane potential (MMP), distribution of phosphatidylserine (PS), and membrane integrity of frozen–thawed ram spermatozoa were measured in this study.
Materials and Methods
Extenders
Unless otherwise stated, all chemicals were of analytical reagent grade and purchased from Sigma Chemical Co.
The basic extender used in this study was composed of 224.0 mM Tris, 66.6 mM citric acid, and 55.5 mM monosaccharide (glucose or fructose) or sugar alcohol (mannitol, sorbitol, or xylitol). The freezing extenders were composed of 224.0 mM Tris, 66.6 mM citric acid, 55.5 mM monosaccharide (glucose or fructose) or sugar alcohol (mannitol, sorbitol, or xylitol), 10% (V/V) egg yolk, 5% (V/V) glycerol, 10,000 IU penicillin, and 10,000 IU streptomycin. Based on the first alphabet letter of the monosaccharide or sugar alcohol, the treating groups were named by G, F, M, S, and X, respectively. The extenders were adjusted to pH 7.0 using Tris. The osmolarity of the basic extender and freezing extenders was measured using FISKE ONE-TEN Osmometer (FISKE Associates). The osmometer can only measure crystal osmotic pressure and cannot be used for measurement of colloid osmotic pressure, so the effects of egg yolk and glycerol on measurement of crystal osmotic pressure should be precluded. The osmolarity of the freezing extenders in which egg yolk and glycerol were substituted by water was ∼330 mOsm. The freezing extenders were centrifuged at 15000 g for 1 hour at 4°C. Finally, the supernatants were filtered through a 0.45 μm membrane filter (Millipore Corporation).
Animals and semen collection
This study was performed at Yunnan Animal Science and Veterinary Institute, which is located at Kunming city, Yunnan province, China (103°40′ E; 26°22′ N). The Yunnan semifine wool rams were raised under uniform feeding, housing, and light conditions. The rams had free access to water and a salt lick. Semen was collected from six adult (3–4 years) rams with proven fertility using electric stimulation. The semen collection was performed six times for each ram. The time interval between each semen collection was 2 days.
The tubes containing semen were placed in a 37°C water bath and immediately transported to the laboratory within 15 min. The semen quality assessment was performed immediately after semen collection. The concentration and motility of spermatozoa were evaluated using a computer-assisted spermatozoa analyzer system (CASAS) (MaiLang; Songjingtianlun Co.). The semen used in this study must meet the following criteria: volume of 0.75–2 mL; minimum concentration of 3 × 109 spermatozoa/mL; and motility higher than 75%. Following the quality assessment, the ejaculates of six rams, each time, were pooled equally to minimize individual differences. 40 The quality assessment of ram semen was performed by the same person during the experiments.
Cooling, freezing, and thawing procedure
Each time, the pooled semen was divided into five equal parts and diluted with the corresponding extender prewarmed at 37°C, respectively. The final spermatozoa concentration in the diluted semen was ∼5 × 108/mL. The diluted semen was loaded into 0.25 mL plastic straws (IMV), cooled to 5°C at 0.0035°C/s, and then further equilibrated for 2 hours. After cooling and equilibration, the straws were put on an aluminum plate (100 × 2 × 400 mm) and frozen in liquid nitrogen vapor. The freezing rate was ∼80°C/min. The distance between the aluminum plate and the surface of liquid nitrogen was 5 cm. After prefreezing for 7 minutes in the liquid nitrogen vapor, the straws were directly plunged into liquid nitrogen and preserved for 1 month.
In this study, the cooling/freezing rate was measured using a thermometer (Testo 925; TESTO). The thermocouple probe was inserted into the middle of a plastic straw to record the changing trends of temperature during cooling and freezing. Simultaneously, the cooling time (from 37°C to 5°C) and freezing time (from 5°C to −50°C) were recorded. The cooling rate was calculated using the following formula: (5°C − 37°C)/the cooling time (seconds). The freezing rate was calculated using the following formula: (−50°C − 5°C)/the freezing time (seconds). After storage in liquid nitrogen for 1 month, the straws were directly thawed in a 37°C water bath for 30 seconds. The concise schematic diagram describing the freeze–thaw cycle is shown in Figure 1.
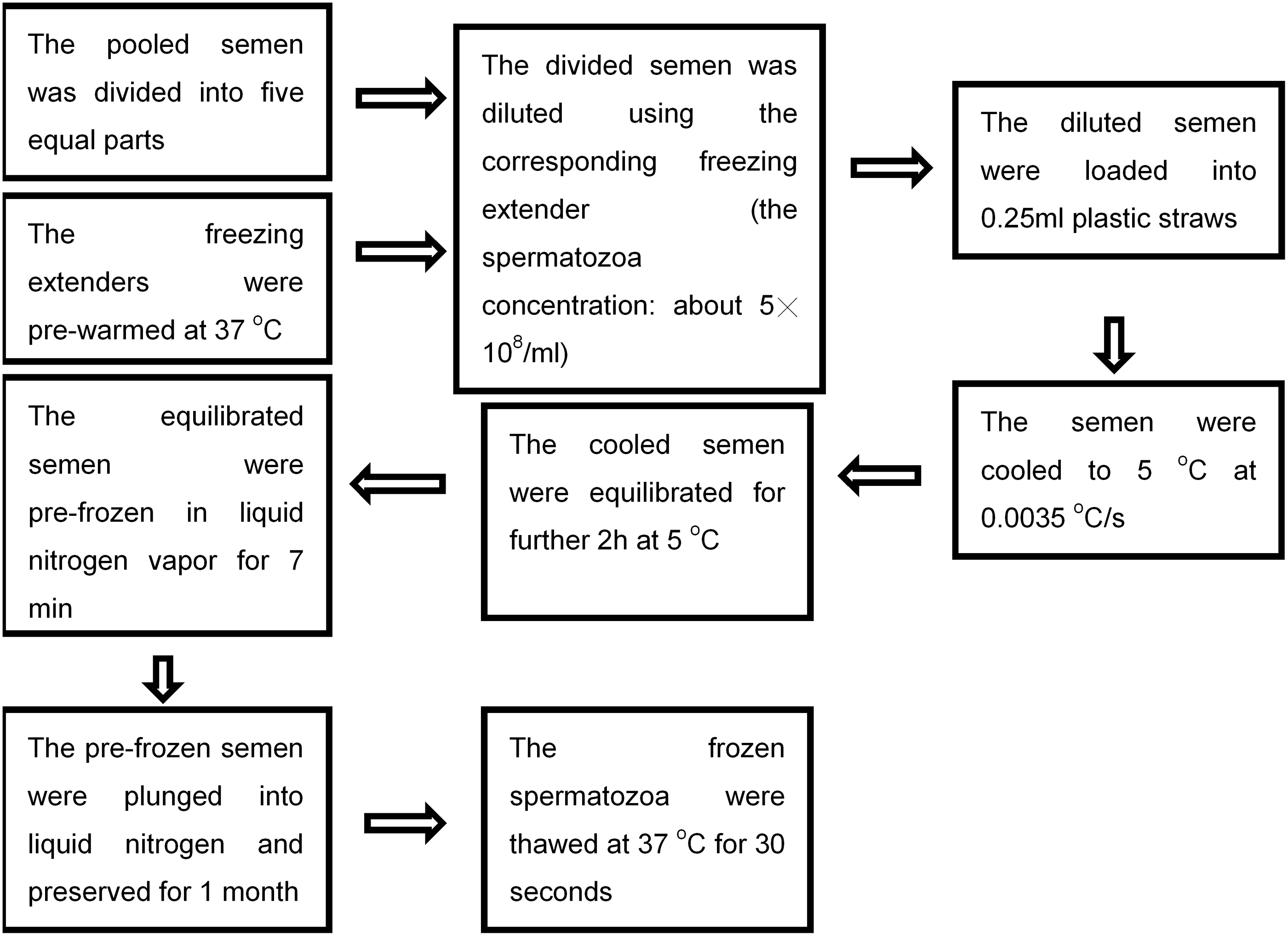
The concise schematic diagram showing the freeze–thaw cycle.
Motility and velocity
The frozen–thawed spermatozoa were further diluted using phosphate-buffered saline (PBS) in a ratio of 1: 5 (V/V) and equilibrated in a 37°C water bath for 10 minutes. Then, the motility and moving velocity of ram spermatozoa were measured using the CASAS. The parameters of the CASAS were set as follows: frame required = 25, the fields of vision acquired = 10, the gray threshold = 60, minimum size of spermatozoa = 5, and maximum size of spermatozoa = 50.
Acrosome status
The acrosome status of spermatozoa was assessed using fluorescein isothiocyanate-stained lectin from pisum sativum (FITC-PSA). The operating procedure has been described previously.2,41 Briefly, aliquots (15 μL) of semen samples were fixed in 4% paraformaldehyde for 30 minutes at room temperature. Then 30 μL of FITC-PSA diluted in PBS (200 μg/mL) was added to the upper suspension. The mixed suspensions were incubated in a dark and moist chamber for 30 minutes at 37°C. Finally, the percentage of spermatozoa stained with FITC-PSA was analyzed by flow cytometry. The assessment of acrosome status was based on the fluorescence intensity. The fluorescence intensity of spermatozoa with intact acrosome is higher compared with spermatozoa with damaged or partly damaged acrosome.
Hypoosmotic swelling test
The functional integrity of spermatozoa membrane was evaluated using the hypoosmotic swelling test (HOST). The procedure was based on the investigations carried out by Matsuoka et al. 42 and Kulaksiz et al. 43 Briefly, after freezing and thawing, 25 μL of semen was mixed equally with 250 μL of a 100 mOsm hypotonic medium (57.6 mM fructose and 19.2 mM sodium citrate) and incubated at 37°C for 30 minutes. After incubation, 0.1 mL of the spermatozoa suspension was smeared on a slide with a cover slip. Four hundred spermatozoa were counted using the microscope equipped on the CASAS. The number of spermatozoa with looped tails was recorded. The percentage of spermatozoa with normal hypoosmotic swelling capability was calculated by the following formula: the number of spermatozoa with looped tails/400 × 100%.
Membrane integrity and PS distribution
The effects of monosaccharide or sugar alcohol on membrane integrity and PS distribution of frozen–thawed ram spermatozoa were assessed using an Apoptosis Detection Kit (KeyGEN Biology). Propidium iodide (PI) and Annexin V were included in this kit. PI cannot penetrate intact membrane of normal cells. However, it can penetrate the membrane of necrotic cells to stain the nucleus, therefore, PI can be used to determine membrane integrity of mammalian cells. Exposure of PS is a typically apoptotic feature. Annexin V can specifically stain exposed PS. In this study, 5 × 105 ram spermatozoa were equally mixed with 250 μL of binding buffer. Five microliters of PI and Annexin V-FITC were then supplemented to the above suspension.
After incubation for 15 minutes in the dark at room temperature, the percentages of spermatozoa stained with PI or Annexin V were analyzed using flow cytometry. Spermatozoa were classified into four populations as follows: PI-positive population, dead; Annexin V-FITC-positive and PI-negative population, early apoptotic; both Annexin V-FITC- and PI-positive population, late apoptotic; and both Annexin V-FITC- and PI-negative population, normal. In this study, the spermatozoa with intact membrane included early apoptotic and normal spermatozoa.
Mitochondrial membrane potential
Disruption of MMP is one of intracellular events occurring after induction of apoptosis. A MitoLight™ Apoptosis Detection Kit (Catalog No. APT142; Chemicon International) was used to evaluate MMP of frozen–thawed spermatozoa. As a lipophilic indicator, MitoLight (5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolylcarbocyaninechloride) can differentially stain mitochondria with high or low membrane potential. 44 MitoLight that has aggregated in mitochondria of healthy cells fluoresces red. However, if MitoLight cannot accumulate in mitochondria of apoptotic cells, it remains as monomers in the cytoplasm and fluoresces green.
Briefly, the semen suspension containing ∼1.0 × 106 spermatozoa was centrifuged at 500 g for 5 minutes to pellet spermatozoa. The cell sediment was then suspended in 1 mL of the prediluted MitoLight solution and incubated at 37°C in a 5% CO2 incubator for 20 minutes. Following incubation, the cell suspension was centrifuged to remove the supernatant. Finally, the cell sediment was resuspended in 1 mL of 1× incubation buffer (dilute 100 μL of 10× incubation buffer with 900 μL of deionized water) before analysis using flow cytometry.
Flow cytometry
The percentages of ram spermatozoa stained with FITC-PSA, MitoLight, PI, or Annexin V-FITC were analyzed using a FacStar-plus flow cytometry (FACSCalibur; Becton Dickinson and Co.). The excitation wavelength was set as 488 nm. The emission wavelength was set as 530 nm. The green fluorescence of FITC and MitoLight monomer was detected by the FL1 detector, and the red fluorescence of PI and MitoLight aggregates were detected by the FL2 detector. Data were collected from 10,000 events for further analysis with the Cell-Quest software (Becton Dickinson).
Statistical analyses
All statistical analyses were performed using the SPSS Software (Version 11.0.1; SPSS, Inc.). The data about the effects of monosaccharide or sugar alcohol on frozen–thawed ram spermatozoa were collected from six different rams in this study. Each time, the collected semen from six rams was pooled to preclude the effects of individual variation. The whole experiment was repeated six times (n = 6). The percentage data were subjected to arcsine transformation before statistical analysis. The data were expressed as mean ± SEM. The differences between the treating groups were analyzed by one-way ANOVA combined LSD's test. All results with a value of p < 0.05 or p < 0.01 were considered to be statistically significant.
Results
Effects of monosaccharide or sugar alcohol on the motility and moving velocity of ram spermatozoa after equilibration at 5°C or cryopreservation
The data in Table 1 indicated that after equilibration at 5°C, there was no significant difference existing among these treated groups regardless of monosaccharide or sugar alcohol. The motility and moving velocity of ram spermatozoa treated with monosaccharide were similar to those of spermatozoa treated with sugar alcohol after the cooling and equilibration procedure (p > 0.05).
“G, F, M, S, and X” represented “glucose, fructose, mannitol, sorbitol, and xylitol,” respectively. Data points are expressed as mean ± SEM, n = 6.
The data about the effects of monosaccharide and sugar alcohol on frozen–thawed ram spermatozoa are presented in Table 2. When mannitol or sorbitol was presented in the freezing extender, the postthaw motility of ram spermatozoa was 52.59% ± 5.82% and 53.71% ± 5.58%, respectively, which was significantly higher than that in the other groups (p < 0.05). Among the five treatment groups, the motility of spermatozoa frozen in the presence of xylitol was only 35.98% ± 6.21%, which was significantly less than the monosaccharide groups (p < 0.05). In addition, compared to mannitol or sorbitol, the protective effect of xylitol showed a significant decrease (p < 0.01). However, there is no significant difference existing between mannitol and sorbitol with respect to the motility (p > 0.05).
Data points are expressed as mean ± SEM, n = 6. The data labeled with “*” or “**” within a column are significantly different from spermatozoa frozen in the M or S group (p < 0.05 or p < 0.01). Different superscript letters (a, b) indicate significant difference existing between either G or F and X (p < 0.05). The data in same column labeled with same superscript letter or no superscript letter indicate no significant difference (p > 0.05).
As indicated in Table 2, the changing trend of (VAP, the average velocity of the smoothed cell path) of frozen–thawed spermatozoa was similar to that of the motility. The VAP of spermatozoa frozen in the presence of mannitol or sorbitol showed a significant increase compared to the other groups. When xylitol was used, the VAP of frozen–thawed spermatozoa was only 60.27 ± 7.34 μm/s and significantly less than those in the other groups (p < 0.05). Compared to either glucose or fructose, both mannitol and sorbitol can increase the postthaw VAP of spermatozoa to ∼70 μm/s. Moreover, there is no significant difference existing between glucose and fructose or between mannitol and sorbitol with respect to the VAP (p > 0.05).
With respect to the linear velocity (VSL, the average velocity measured in a straight line from the beginning to the end of the track), the postthaw VSL of spermatozoa frozen in the presence of xylitol was only 46.38 ± 6.42 μm/s and significantly less than the other groups (p < 0.05). However, the VSL of spermatozoa frozen in the presence of either mannitol or sorbitol was not significantly different from that of spermatozoa frozen with either glucose or fructose (p > 0.05).
Concerning the curve velocity (VCL, the average velocity measured over the actual point to point track followed by the cell), its changing trend was similar to that of the VAP. When either mannitol or sorbitol was presented in the freezing extender, the VCL of frozen–thawed spermatozoa was 84.18 ± 8.92 μm/s and 82.57 ± 5.16 μm/s, respectively, which were significantly higher than the other groups (p < 0.01). In addition, there is no significant difference existing between mannitol and sorbitol (p > 0.05). When xylitol was present in the freezing extender, the VCL of frozen–thawed spermatozoa was 68.29 ± 7.14 μm/s and significantly less compared with spermatozoa frozen with either glucose or fructose (76.03 ± 6.29 μm/s and 74.38 ± 5.28 μm/s, p < 0.05). Moreover, the VCL of spermatozoa frozen with glucose was not significantly different from that of spermatozoa frozen with fructose (p > 0.05).
Effects of monosaccharide and sugar alcohol on acrosome status of frozen–thawed ram spermatozoa
As seen in Figure 2, when monosaccharide was replaced by either mannitol or sorbitol, the percentage of frozen–thawed spermatozoa with intact acrosome was 51.36% ± 4.68% and 49.32% ± 7.18%, respectively, which was significantly higher compared with spermatozoa frozen with either glucose or fructose (35.24% ± 5.47% and 32.78% ± 4.97%, p < 0.01). In addition, the frequency with intact acrosome in spermatozoa frozen with xylitol was significantly higher than the glucose or fructose group (p < 0.05), although less than mannitol and sorbitol. Moreover, there is no significant difference existing between mannitol and sorbitol or between glucose and fructose with respect to acrosome status (p > 0.05).
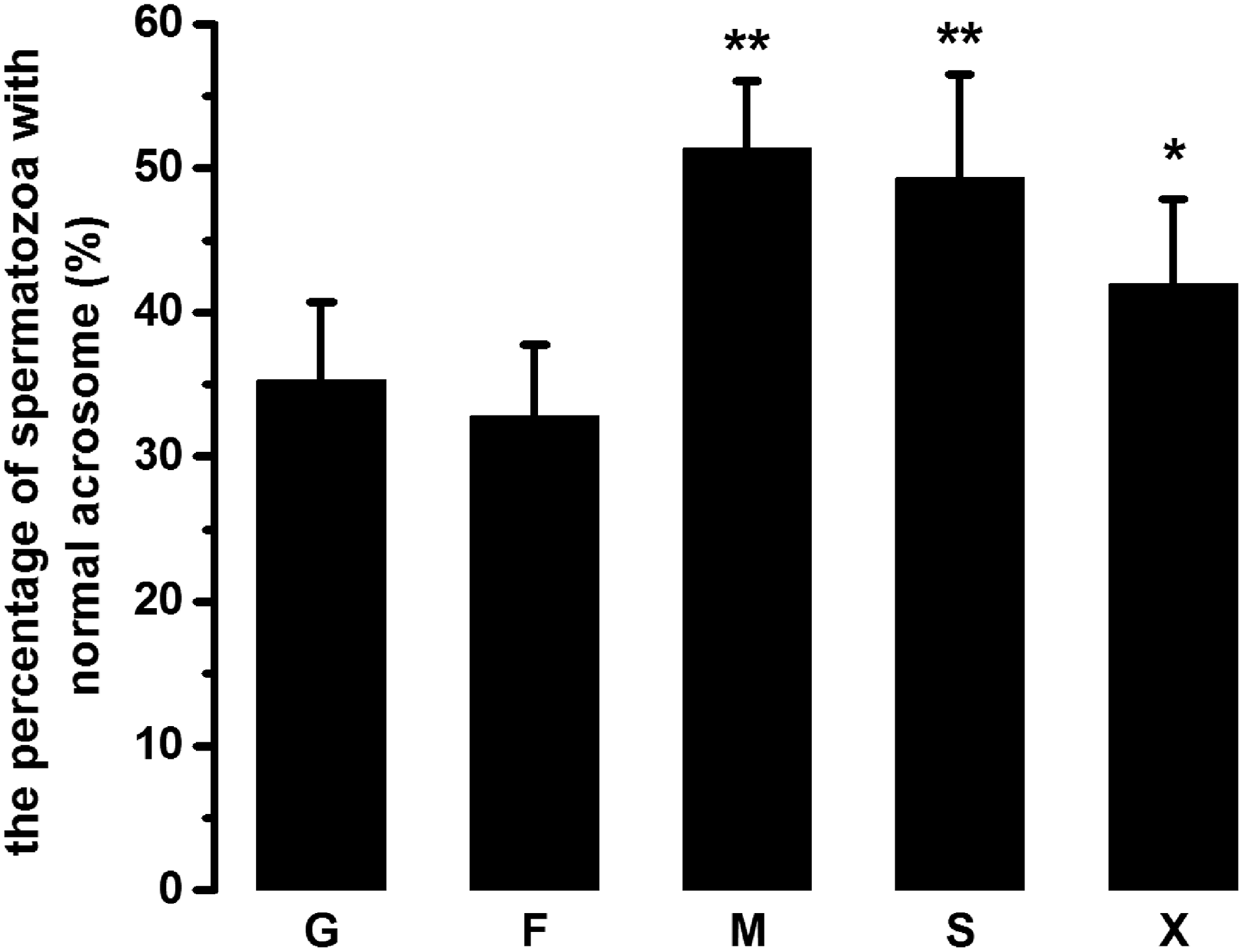
The effects of either monosaccharides or sugar alcohols on acrosome status of frozen–thawed ram spermatozoa. The data points are expressed as mean ± SEM, n = 6 in each group. **p < 0.01 versus either glucose or fructose. *p < 0.05 versus the other treating groups.
Effects of monosaccharide and sugar alcohol on hypo-osmotic swelling capability of frozen–thawed ram spermatozoa
As indicated in Figure 3, when either mannitol or sorbitol was presented in the freezing extender, the percentage of spermatozoa with normal hypoosmotic swelling capability was 28.92% ± 3.86% and 30.28% ± 5.94%, respectively, which was significantly higher than the other three groups (p < 0.05). Moreover, there is no significant difference existing between these two groups (p > 0.05). In addition, the frequency with normal hypoosmotic swelling capability in spermatozoa frozen with either glucose or fructose was not significantly different from spermatozoa frozen with xylitol (p > 0.05).
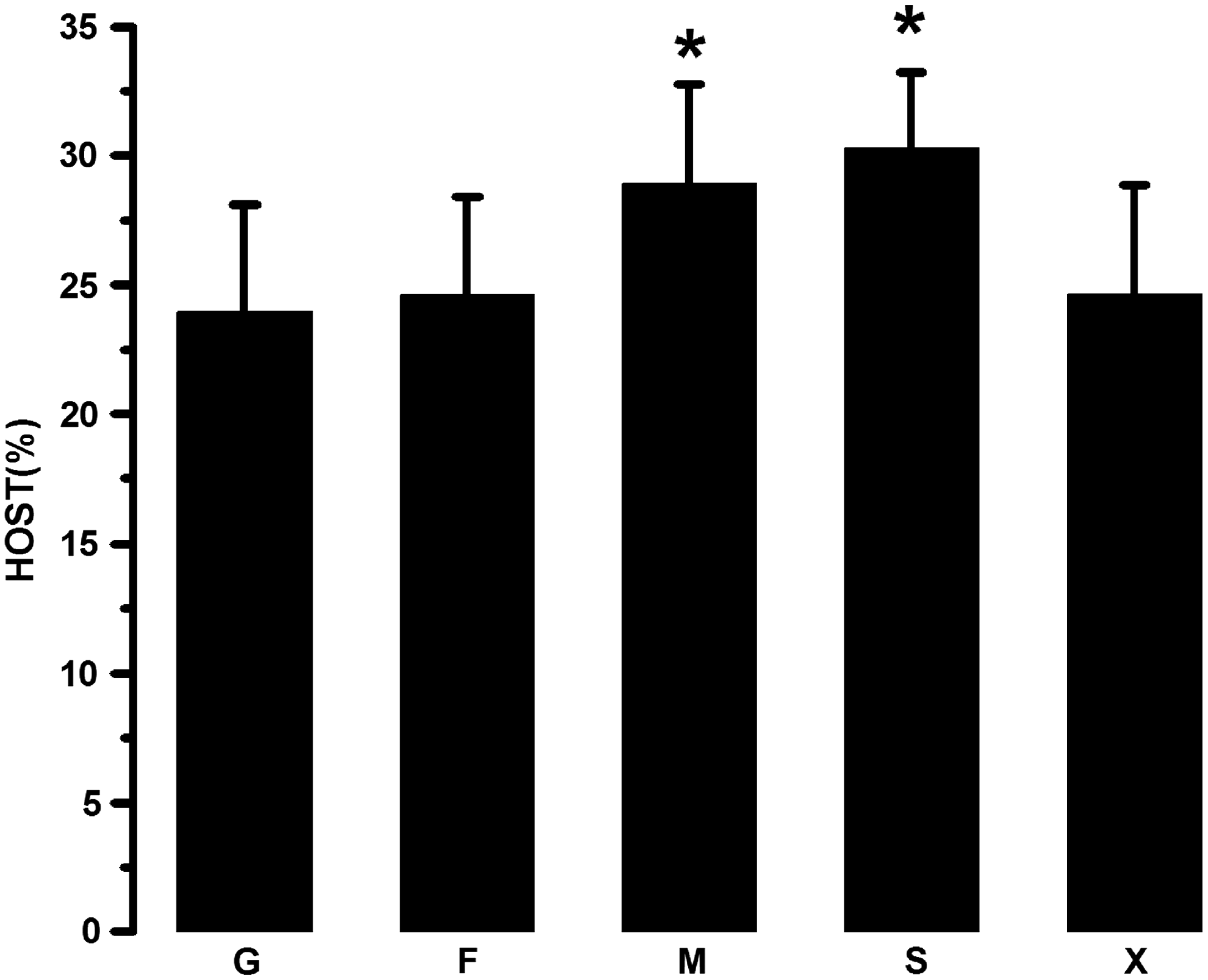
The effects of monosaccharides or sugar alcohols on hypoosmotic swelling capability of frozen–thawed ram spermatozoa. The data were expressed as mean ± SEM, n = 6 in each group. *p < 0.05 versus the other treating groups.
Effects of monosaccharide and sugar alcohol on membrane integrity and PS distribution of frozen–thawed ram spermatozoa
The data about the effects of monosaccharide and sugar alcohol on membrane integrity and PS distribution of frozen–thawed ram spermatozoa are present in Figure 4. As indicated in Figure 4A, when either mannitol or sorbitol was presented in the freezing extender, the percentage of frozen–thawed spermatozoa with intact membrane was 46.32% ± 5.74% and 45.95% ± 6.26%, respectively, which were significantly higher than those of the other groups (p < 0.05). In addition, no significant difference was observed between mannitol and sorbitol (p > 0.05). The frequencies with intact membrane in spermatozoa frozen with glucose, fructose, or xylitol were ∼36%. There is no significant difference existing among these three treatment groups (p > 0.05).
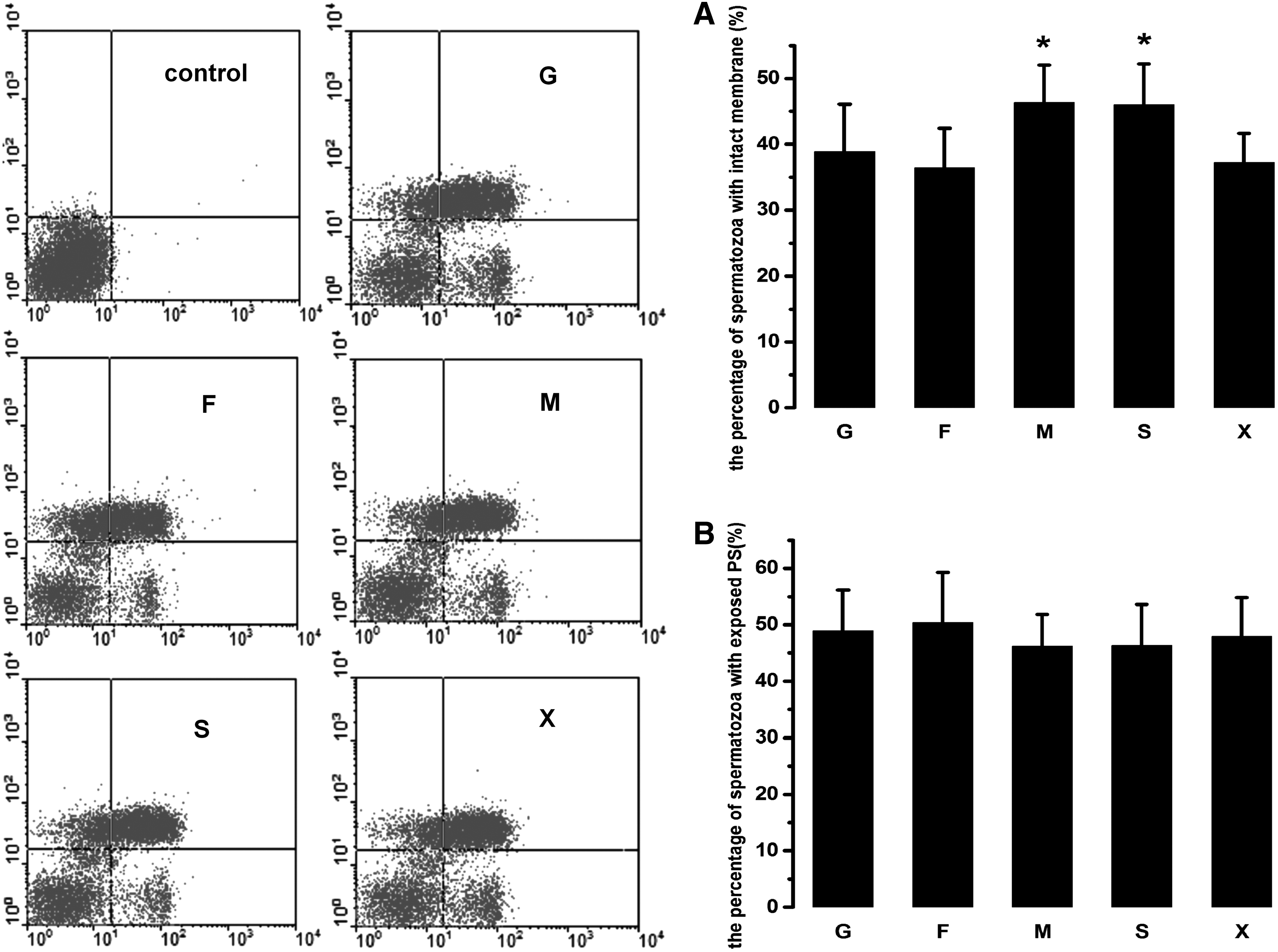
The effects of monosaccharides or sugar alcohols on membrane integrity and PS distribution of frozen–thawed ram spermatozoa. “Control” represents frozen–thawed spermatozoa without staining with both Annexin V and PI. The capitals G, F, M, S, or X represent the scatter plots of ram spermatozoa frozen with glucose, fructose, mannitol, sorbitol, and xylitol, respectively.
As shown in Figure 4B, with respect to PS distribution, the percentages of frozen–thawed spermatozoa with exposed PS in the sugar alcohol groups were similar to those in the monosaccharide groups (p > 0.05), although the frequency with exposed PS in spermatozoa frozen with either glucose or fructose was slightly higher than the sugar alcohol groups.
Effects of monosaccharide and sugar alcohol on MMP of frozen–thawed ram spermatozoa
As shown in Figure 5, when the sugar alcohols were present in the freezing extenders, the frequency with normal MMP was ∼70% and significantly higher than that in the glucose or fructose group, which was only ∼60% (p < 0.05). In addition, there is no significant difference existing among the monosaccharide groups or the sugar alcohol groups (p > 0.05).
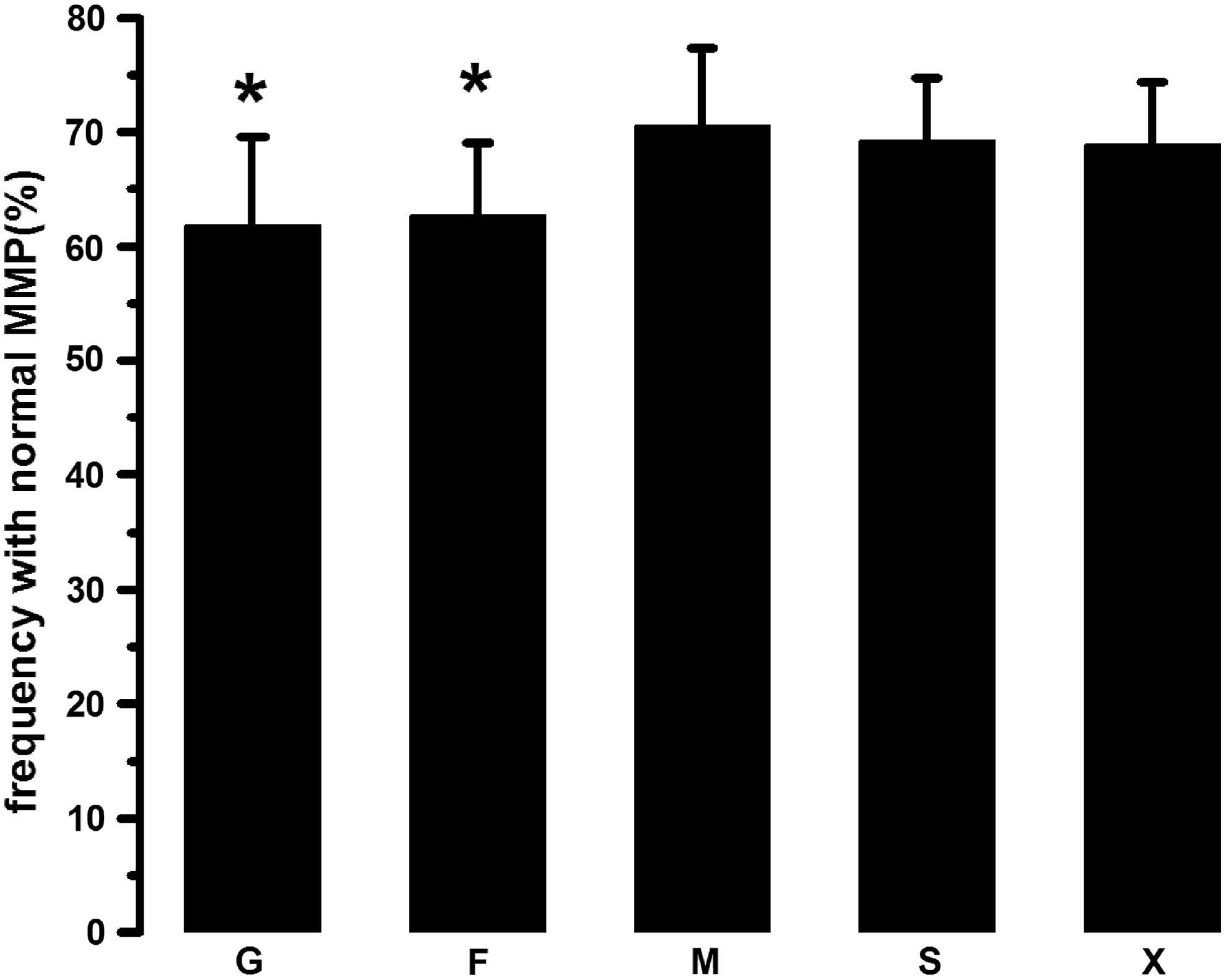
The effects of monosaccharides or sugar alcohols on MMP of frozen–thawed ram spermatozoa. The capitals, such as G, F, M, S, and X, represent ram spermatozoa frozen with glucose, fructose, mannitol, sorbitol, and xylitol, respectively. The data are expressed as mean ± SEM, n = 6 in each group. *p < 0.05 versus the sugar alcohol groups. MMP, mitochondrial membrane potential.
Discussion
Currently, as primary components, either glucose or fructose is generally included in dilution or freezing extenders during cryopreservation of mammalian spermatozoa. 6 According to some reports, monosaccharides can act as primary substrates for glycolysis, maintain osmotic balance, and stabilize cell plasma membrane during cryopreservation.3,18,28
However, investigations about the effect of sugar alcohol on frozen–thawed spermatozoa were relatively few compared to monosaccharide. Moreover, owing to the components of extenders, freezing and thawing procedures, and animal species, disputes regarding whether sugar alcohols are beneficial for cryosurvival of mammalian spermatozoa still exist. In this study, the effects of the main monosaccharides and sugar alcohols on frozen–thawed ram spermatozoa were investigated and compared. To preclude the effects of osmotic difference, the osmotic pressure of the freezing extenders used in this study was held constant.
Our data indicated that the cryoprotective effects of sugar alcohol, especially mannitol or sorbitol, were superior to monosaccharides with respect to the parameters evaluated in this study. For example, when a monosaccharide was replaced by mannitol or sorbitol in the freezing extender, the motility, moving velocity, acrosome and membrane integrity, HOST, and MMP of frozen–thawed spermatozoa showed a significant improvement. Some investigations have indicated that sorbitol or mannitol can provide beneficial cryoprotection for mammalian spermatozoa.18,33,37 Sorbitol has been demonstrated to improve cryotolerance of spermatozoa from bull, 37 ram, 33 stallion, 18 and human. 34 Mannitol can improve the motility of frozen–thawed ram spermatozoa. 33
However, the mechanism underlying the protective effects of sugar alcohols against cryoinjury still needs further elucidation. According to some investigations, both mannitol and sorbitol have a similar structure and molecular size, which mean that they may have a similar physiological function. 19 Sugar alcohols are sugar molecules in which the aldehyde group is replaced by an alcohol group. 17 Therefore the beneficial cryoprotection of sugar alcohols may be due to their structural similarity to glycerol. 33 In addition, a synergetic effect existing between sugar alcohols and glycerol can be used to explain the cryoprotective action of sugar alcohols.18,33
According to some reports, sugar alcohols may strengthen the tolerance of mammalian spermatozoa to oxidative stress. 45 For example, sorbitol can stabilize the plasma membrane when spermatozoa are exposed to oxidative stress. 45 Finally, sorbitol can act as an alternative energy substrate for spermatozoa glycolysis and oxidative phosphorylation21,46 and can be converted into fructose by the sorbitol dehydrogenase enzyme to allow it to penetrate the cell plasma membrane. 47 In the investigation carried out by Cao et al., sorbitol enhanced the motility of mouse spermatozoa by protein tyrosine phosphorylation. 35
Cryopreservation and thawing can seriously damage the spermatozoa membrane. How to efficiently protect structure and physiological function of spermatozoa membrane is a great challenge. It has been widely recognized that the HOST is a simple and efficient method with good reliability and repeatability for assessment of spermatozoa membrane. 42 Moreover, the results of HOST have been proposed to tightly correlate with actual fertility. 48 This study indicated that the replacement of monosaccharide by either mannitol or sorbitol in the freezing extender could efficiently improve the postthaw functional status of spermatozoa membrane, which may correlate with the fact that sugar alcohol can enhance the osmotic tolerance of mammalian spermatozoa.
To further determine whether sugar alcohols have positive effects on membrane integrity of frozen–thawed spermatozoa, both fluorescence staining and flow cytometry were also used to assess the membrane status in this study. We found that when either mannitol or sorbitol was included in the freezing extender, the percentage of frozen–thawed spermatozoa with intact membrane was significantly higher than those of spermatozoa frozen with monosaccharide, which demonstrated that the capability of sorbitol or mannitol to protect spermatozoa membrane against cryoinjury might be superior to that of glucose or fructose.
Cryopreservation can result in the exposure of PS, which is an early feature of cell apoptosis. 49 In this study, the capability of either mannitol or sorbitol to inhibit PS exposure of frozen–thawed spermatozoa was slightly higher than those of both glucose and fructose, although the difference is not significant. In addition, as another important sign of cell apoptosis, the changing trend of MMP was also evaluated. Our data indicated that compared to either glucose or fructose, sugar alcohols, such as mannitol, sorbitol, or xylitol, can efficiently maintain the postthaw stability of MMP in ram spermatozoa.
Concerning the effects of monosaccharide or sugar alcohol on PS distribution and MMP, this study indicated that sugar alcohols might postpone spermatozoa apoptosis caused by cryopreservation and thawing in the ram. In this study, it must be noted that apoptosis caused by cryopreservation may correlate with oxidative stress. According to some reports, sugar alcohol may have certain antioxidative action. 45 As described in an investigation carried out by Hincha and Hagemann, sorbitol can enhance the tolerance of spermatozoa to oxidative stress. 45 In addition, mannitol can delay rapid peroxidation in ram spermatozoa when they are exposed to reactive oxygen species (ROS)-generating systems. 50
During cryopreservation, long-term cooling and equilibration at reduced temperatures are necessary steps for cryosurvival of ram spermatozoa. However, during this period, reduced temperature cannot completely inhibit the metabolism of ram spermatozoa. Therefore, the accumulation of toxic products, including free radicals, may lead to apoptosis of ram spermatozoa. In addition, owing to the limitation of experimental conditions, cryopreservation of ram spermatozoa is commonly performed in an aerobic or even a partially aerobic environment. Contact with oxygen still has a capability of maintaining the aerobic metabolism of ram spermatozoa during equilibration at reduced temperature. 51 So the production of ROS is inevitable during freezing and thawing.
In addition, monosaccharides are the principal reducing sugars in seminal plasma and thus serve as the main glycolyzable substrates for ram spermatozoa. 52 Depending on the animal species, the fructose concentrations in seminal plasma vary from high levels in bull and ram to lower concentrations in other animal species, such as dog or stallion, where it is essentially absent. 21 We speculate that compared to sorbitol, ram spermatozoa tend to use fructose as the primary glycolyzable substrate. So the oxidative injuries caused by metabolism of either fructose or glucose may be more serious than either mannitol or sorbitol. Although the effects of oxidative stress on the viability of frozen–thawed ram spermatozoa were not investigated in this study, an approach to decrease the negative effects of oxidative stress on ram spermatozoa is essential for the cryosurvival of ram spermatozoa.
This study also demonstrated that there was inferior cryoprotection of xylitol on frozen–thawed spermatozoa compared to mannitol or sorbitol. Our results were contrary to the reports of Molinia et al. 33 In their investigation, xylitol showed a significant protection for frozen–thawed ram spermatozoa. They concluded that xylitol might more easily penetrate cell membranes because its molecular size is less compared with either mannitol or sorbitol. 33 In addition, in an investigation carried out by Salamon et al., xylitol also provided some cryoprotection to boar spermatozoa during cryopreservation. 39 These differences may be attributed to the specific experimental conditions and animal species.
Glycerol is not presented in the freezing extenders in the investigation of Molinia et al., 33 which may explain why the cryoprotective effects of xylitol are compromised in this study. In addition, the optimal concentrations of xylitol in the freezing extender used by Salamon et al. varied from 0.75 to 1.5 M and were largely higher than that used in this study. 39
Conclusion
This study demonstrated that the presence of either monosaccharides or sugar alcohols in the freezing extenders exerted different protective roles against cryoinjury during cryopreservation of ram spermatozoa. The replacement of glucose or fructose by mannitol or sorbitol in the freezing extender may improve the postthaw viability of ram spermatozoa under specific freezing conditions.
In addition, except for the protective effects on acrosome and MMP, xylitol does not show positive cryoprotection on ram spermatozoa concerned with the motility, velocity, PS distribution, and membrane integrity. However, the plasma membrane of ram spermatozoa still experienced serious cryoinjury during freezing and thawing, although the protective effects of either mannitol or sorbitol on plasma membrane of ram spermatozoa are superior to glucose or fructose. In the future, the approach to reducing cryoinjuries on ram spermatozoa will require further research.
Footnotes
Acknowledgments
The authors would like to thank Mr. Jian Li from Kunming Institute of Zoology for his help with flow cytometry. We also sincerely thank Dr. Shen from Qingdao Agriculture University and Dr. Wang from China Agriculture University for polishing the English writing in this article. This investigation was funded by the National Natural Science Foundation of China (Grant No. 31360551 and 31560635), the National Wool Caprine Industrial Technology System (Grant No. CARS-40), the Middle and Young Academic Leaders of Yunnan Province (Grant No. 2014HB051), the High-Tech Enterprise Development Plan of Yunnan Province (Grant No. 201404), and the National Infrastructure of Animal Genetic Resources (NIAGR).
Author Disclosure Statement
No conflicting financial interests exist.