
Abstract
Quantitative evaluation of the inherent correlation between cell cryoinjuries and intracellular ice formation (IIF) together with recrystallization (IIR) is of primary importance for both optimization of biopreservation and cryotherapy. The objective of this study is to thoroughly explore the roles of IIF on cell viability by using pig iliac endothelium cells (PIECs) as model cells during freezing and thawing. The experimental results indicated that both the probabilities of IIF (PIF) and IIR (PIR) increased along with the increase of cooling rates (p < 0.05) during the freeze–thaw cycles at cooling rates of 40, 60, 80, 100, and 150°C/min and the same warming rates of 100°C/min in phosphate-buffered saline-based solutions with or without 1 M DMSO. Viability evaluation with Hoechst 33342/propidium iodide double staining showed that most of the cells were killed (viability <20%) by the abovementioned freeze–thaw cycles, which indicated that the cooling rates investigated were all too rapid since large amounts of IIF and IIR were introduced. Another interesting phenomenon is that the presence of a low concentration of DMSO (1 M) tends to improve cell viability while increasing the PIF and PIR during freezing/thawing, contrary to the common belief that larger PIF corresponds to greater cryoinjury. This may be attributed to the intrinsic protection effect of DMSO by reduction of solution injury or other potential injuries. These findings may be of potential application value for both cryopreservation and cryosurgery by providing helpful additions to the existing studies on investigation of cryoinjuries of PIECs.
Introduction
T
During the freezing process of cells, the IIF phenomenon can be observed under a cryomicroscope as a sudden darkening of the cell cytoplasm. 20 Meanwhile, in some situations, IIF may also be characterized by cellular twitching due to ice crystals that are too small to scatter light in cell cytoplasm.21–23 The study of IIF events led to a successful cryopreservation protocol through the adjustments of CPA concentration and cooling rate. 24 In general, different kinds of cells show various phenomena during the process of freezing and thawing. Thus, the balance between the probability of IIF (PIF) and the concentration of CPAs results in different optimal cooling rates and concentration of CPAs for different cells. In our previous research, human embryonic kidney cells (HEK293) and HeLa cells were studied with regard to PIF and the optimal cooling rates,25,26 and the results showed that these two kinds of cells have different optimal cooling rates and different PIFs at the same cooling rate.
Currently, it remains controversial whether IIF produces lethal damage to cells. Most studies have shown that IIF is potentially lethal to cells,1–3,8,27,28 but as Morris et al. proposed, horse spermatozoa lost viability due to osmotic imbalance during thawing, not IIF during a rapid cooling process. 29 To date, most existing studies focused on IIF during freezing, but there are few reports on IIF during thawing. Huang et al. found that IIF does not occur in all cells during freezing, but IIF starts to occur again during the warming process. 30 Seki and Mazur found also that mouse oocytes underwent no IIF during cooling, but blackened from IIF (recrystallization of intracellular ice) during warming in the post freeze-thaw process, and that IIF during thawing was related to the warming rate. 31 Intracellular ice recrystallization (IIR) is responsible for the death of cells during warming, 32 unfortunately, and recent studies have not significantly discussed this important topic. The related research has recently focused more on ice recrystallization inhibition, and moreover, the relationship of cooling rate with IR has not been elucidated. Tam et al. reported that some small molecules could inhibit IR by hydration. 33 Chaytor et al. 34 pointed out that CPA has an important property to enhance cell viability postfreeze–thaw by inhibiting IIR, while Huang et al.'s investigation on IIF of murine embryonic stem cells (mESCs) indicated that the presence of low concentration of CPA could increase the occurrence of IIF in mESCs during freezing and thawing. 30
At present, most researchers believe that microvascular injury plays a major role in tissue damage in cryosurgery,35–38 and endothelial cells lining the vessel wall are thought to be the initial target of freezing.37,39,40 Therefore, it is very important to study the physiological phenomena of endothelial cells in subzero degree Celsius environments. The present related studies focus more on the effects of duration of cold exposure, end temperature, and so on, but few on the cooling rate and warming rate. The pig iliac endothelial cell (PIEC) is a good model cell for the study, which has been mostly used in biological experiments. This work aims to study the relationship between CPA, cooling rate, IIF, and IIR during freezing–thawing and post-thaw cell viability. We wish to explore the effect of CPA and cooling rate on both IIF and IIR during freezing and thawing, and the relationship between IIF, IIR, and cell injury.
Materials and Methods
Cell culture and sample preparation
The PIECs (purchased from the Institute of Biochemistry and Cell Biology, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences) were cultured in RPMI 1640 medium (Gibco), with 10% (v/v) fetal bovine serum (Hyclone; Thermo Fisher Scientific, Inc.), 100 U/L penicillin G, and 100 U/L streptomycin (Hyclone; Thermo Fisher Scientific, Inc.), and incubated at 37°C, with a 5% CO2 humidified atmosphere. The culture medium was changed every 2 days. The cells were collected at ∼80% confluence. Adherent cells were washed with phosphate-buffered saline (PBS) (GIBCO Invitrogen GmbH) and detached from the culture flasks by addition of 0.2 mL/cm2 0.25% (w/v) trypsin (Sigma-Aldrich) for 5 minutes at 37°C. Then, the cells were centrifuged at 100 g for 5 minutes and resuspended to 1 × 105/mL with 1 × PBS.
CPA addition before freezing
Suspended cells mentioned previously were centrifuged at 100 g for 5 minutes and resuspended to 1 × 105/mL with 1 M dimethyl sulfoxide (DMSO; Sigma). The cells with 1 M DMSO were equilibrated for 5 minutes at room temperature before freezing for the loading of DMSO into the cells.
Cryomicroscopy study
A clean coverslip was placed on the center of the cryostage, and then a drop (∼8 μL) of the abovementioned cell suspension was pipetted onto the center of the coverslip with another coverslip covered on it slightly. The cryostage was mounted on a special copper wire that was used for ice seeding on its edge,25,41 and then about 10 μL PBS was dropped onto the copper wire to seed ice. This method was different from the conventional ice-seeding method that uses a syringe needle precooled in liquid nitrogen and could avoid the dim view during freezing that may cause a negative influence on subsequent observation.
25
The FDCS196 cryostage was fixed to a microscope (BX 53; Olympus). Simultaneously, a Qimaging (Survey) MicroPublisher 5.0 RTV CCD camera was used for real-time monitoring of IIF and IIR (resolution, 960 × 720; frame rate, 5 fps). The PIF and PIR were defined as follows:
Assessment of cell morphology and cell viability
Cell morphology before/after freezing was monitored using a microscope (BX 53; Olympus). Hoechst 33342/propidium iodide (PI) (Keygen) fluorescent stain was applied to determine the cell viability. The cells were directly incubated with Hoechst 33342 and PI for up to 10 minutes in the dark before freezing. Stained cells were visualized by fluorescence microscopy (BX 53; Olympus) connected to a cooled charge-coupled device (CCD) camera. The cellular viability exhibited by the death rate was defined as follows:
Freezing protocols of PIECs
The freezing and thawing protocol is shown Table 1. The samples were cooled from 20°C to −1°C (−4°C for 1 M DMSO) with the constant cooling rate of 10°C/min. To promote ice seeding with copper, the temperature was maintained at −1°C (−4°C for 1 M DMSO) for 30 seconds. To minimize the initial cell dehydration during the ice-seeding process, the sample was warmed back to −0.6°C (−2.5°C for 1 M DMSO) at 10°C/min and held for 3 minutes to achieve equilibrium (ice and water coexist).
Cooling is indicated by negative rates; warming by positive rate.
EIF, extracellular ice formation; PBS, phosphate-buffered saline; PIEC, pig iliac endothelium cell.
Statistical analysis
Statistical analysis was performed using one-way analysis of variance within a group and Student's paired two-tailed t-test between two groups with significance set at p < 0.05. All data are presented as mean ± standard deviation.
Results and Discussion
The effect of CPA addition and cooling rate on IIF
The freezing and thawing process of PIECs was imaged and analyzed using the Irfan View (free from www.irfanview.com). Figure 1 shows the representative photomicrographs of PIECs in PBS and in the presence of 1 M DMSO during the freezing/thawing process at the cooling rate of 60°C/min and warming rate of 100°C/min. The cells were cooled to −1°C/−4°C at 10°C/min and held for 30 seconds to induce ice seeding with copper wire, and then the cells were warmed to −0.5°C/−2.5°C at the same rate (Fig. 1(a1)/(a2)), and temperature was maintained for 3 minutes to achieve equilibrium. Afterward, the samples were cooled to −60°C at 60°C/min, and IIF occurred during this process as the temperature further increased (Fig. 1(b1)/(b2)). On further heating of the cells at 100°C/min, IIF occurred again during the thawing process (Fig. 1(c1)/(c2)). At about 7.8°C/9.5°C, both extracellular and intracellular ice disappeared (Fig. 1(d1)/(d2)). In videos recorded at a frame rate of 25 fps, the occurrence of IIF in a specimen under bright field is characterized by darkening of the cell. Under some situations, IIF may also be characterized by cellular twitching owing to small ice crystals formed in the cell. High-speed video cryomicroscopy studies show that twitching typically precedes darkening, 42 suggesting an initial process where small ice crystals are formed, followed by a recrystallization process where ice crystals grow and begin to scatter light. The ice recrystallization process can also happen during cooling, but most studies focus more on IIR during thawing.8,31,43 Meanwhile, IIR during freezing is visible only by using high-speed CCD (while the common CCD was used in our experimental setup), so we only investigated IIR during thawing in this work. For accurate statistics in the PIF and PIR, we must compare the change of cells between two consecutive frames. As shown in Figure 1, when PIECs were incubated in 1 M DMSO, IIF and IIR were more than the samples in PBS alone. To explain the relationship, IIF and IIR were studied further.
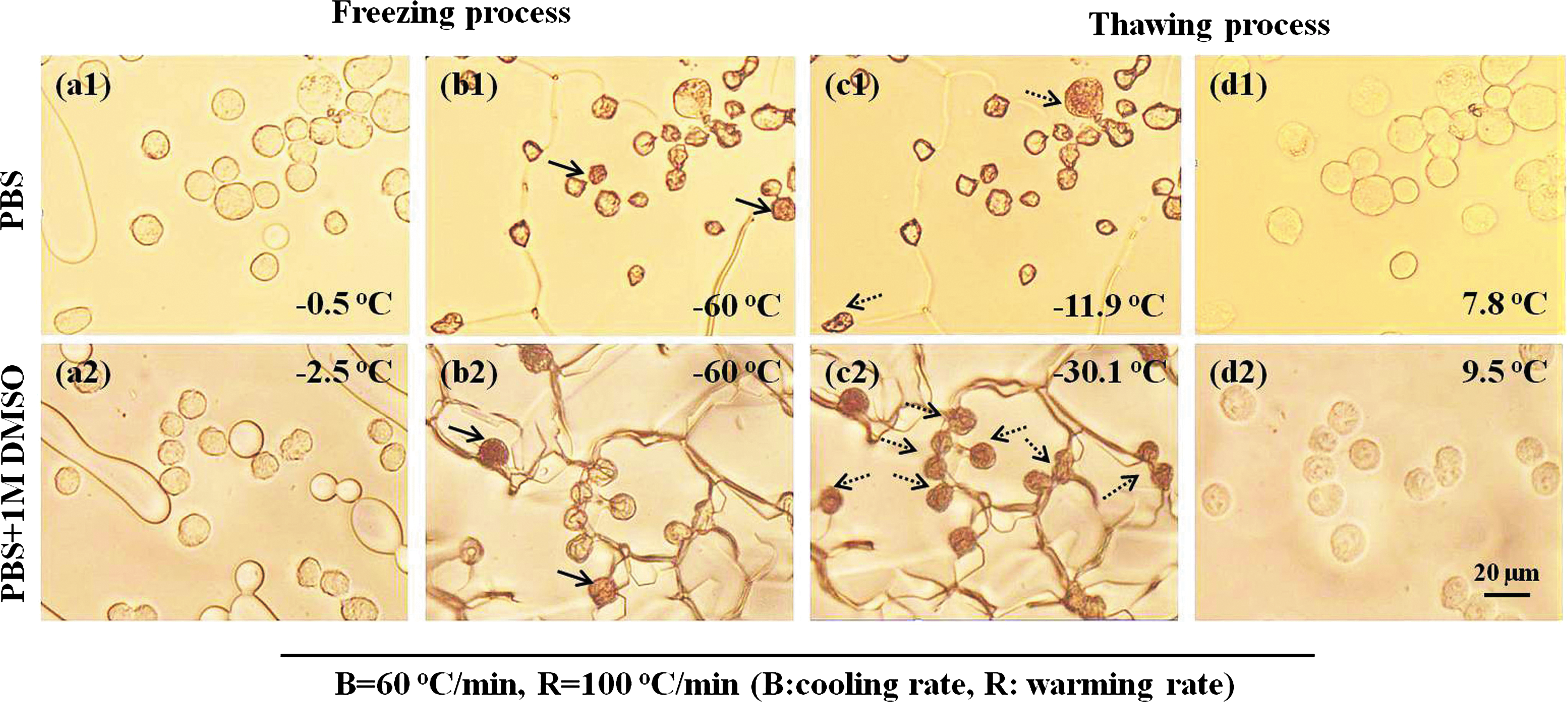
Typical micrographs depicting IIF of PIECs during freezing and thawing (cooling rate: 60°C/min, and warming rate: 100°C/min).
The PIFs of PIECs at various cooling rates are shown in Figure 2a. Approximately 900 cells were chosen for data processing at each cooling/rewarming rate and they were further randomly divided into three subgroups for statistical analysis. For PIECs in PBS, as shown in Figure 2a, IIF occurred at temperature ranges of −7.7°C to −37.4°C, −7°C to −43.6°C, −7°C to −45.6°C, −6.4°C to −48.1°C, and −6.1°C to −48.6°C at 40, 60, 80, 100, and 150°C/min, respectively. These data showed that the starting temperature of IIF increased with the increase of the cooling rate, but meanwhile, the ending temperature of IIF correspondingly decreased.
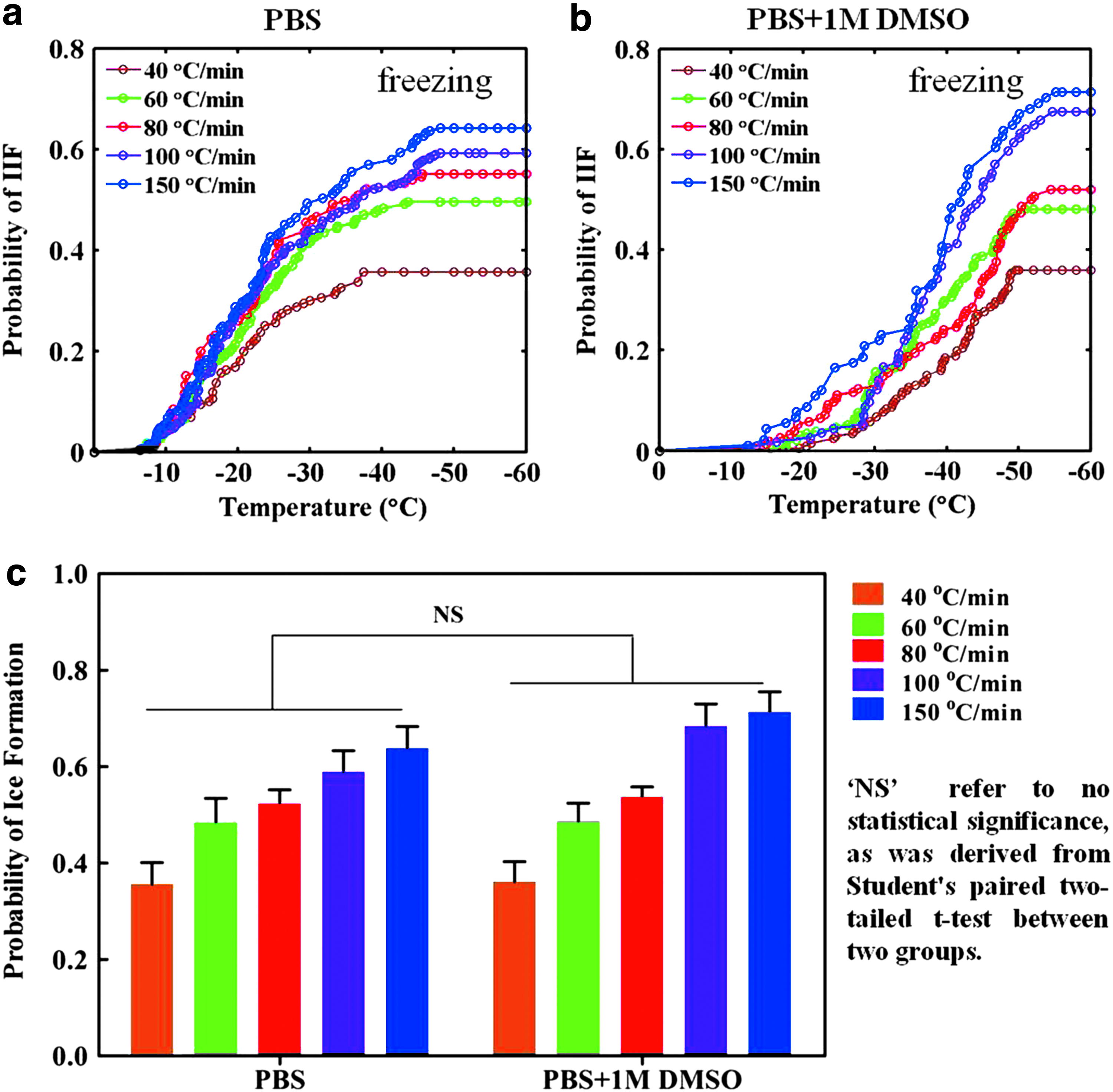
The effects of DMSO addition and cooling rates on IIF. The PIF of PIECs versus temperature at different cooling rates (40, 60, 80, 100, and 150°C/min) and at the warming rate of 100°C/min in the PBS alone
As shown in Figure 2b, in the presence of 1 M DMSO, IIF of PIECs occurred at temperature ranges of −19.5°C to −49.6°C, −15.5°C to −50.9°C, −14.9°C to −54.5°C, −13.5°C to −54.7°C, and −12.3°C to −55.1°C at 40, 60, 80, 100, and 150°C/min, respectively. The starting and ending temperatures of IIF were consistent with that in the absence of CPAs. The temperatures of IIF at various cooling rates for PIECs in PBS alone and in the presence of 1 M DMSO are summarized in Table 2. Compared with PIECs in PBS, the temperatures of IIF in the presence of 1 M DMSO were delayed, respectively, at various cooling rates. The most likely explanation is the greater cell dehydration in PBS. The other reasons that the CPAs could decrease the temperature of ice formation might be that (a) intracellular heterogeneous nucleation is inhibited by changing the thermodynamic stability of ice nuclei; (b) the contacting opportunity of extracellular ice crystals and cell membrane is decreased owing to increasing the amount of intracellular water; and (c) the possibility of extracellular ice crystals into the cell through the cell membrane is reduced by strengthening the stability of the cell membrane. 44
IIF, intracellular ice formation.
The PIFs of PIECs in the presence of 1 M DMSO at various rates of cooling are shown in Figure 2b. It is clear that the cells, which were cooled at higher rates, exhibited higher final values of the PIF and that is concordant with the previous PIFs of PIECs in the absence of CPAs. As shown in Figure 2c, the difference among various rates of cooling was statistically significant (p < 0.05, n = 3). Comparing the PIF of PIECs in PBS alone and in 1 M DMSO, the latter was higher than the former, but the statistical analysis showed no significant difference.
The effect of CPA addition and cooling rates on the PIR
A successful cryopreservation must avoid IIF and IIR. Usually, the PIF can be controlled by slow equilibrium freezing and vitrification in the freezing process, but IIR is hard to avoid during the thawing process. The extremely small ice crystals formed should readily recrystallize and form large crystals during thawing. Slow thawing is recognized to be a prime destructive factor for cryopreservation. The time of thawing is longer, cell damage is greater, and this is because of increased solution effects and maximal growth of ice crystals. 36 Many reports showed that the thawing rate was faster and the PIR was less. However, there are few reports about the quantitative relationship between the cooling rate and IIR during thawing. Our experiment investigated this relationship when PIECs were frozen at 40, 60, 80, 100, and 150°C/min and warmed at 100°C/min.
As shown in Figure 3a and b, the temperature ranges of IIF during warming of PIECs in PBS alone and in 1 M DMSO were −24.7°C to −14.7°C, −29.3°C to −9.3°C, −31.5°C to −10.3°C, −30.8°C to −8.4°C, −30.2°C to −13.4°C and −42.9°C to −29.8°C, −44.3°C to −27.7°C, −40°C to −30.1°C, −44°C to −31.4°C, and −42.9°C to −31.2°C at cooling rates of 40, 60, 80, 100, and 150°C/min and a warming rate of 100°C/min, respectively. The initial and the finishing temperatures of IIR were different between the PIECs in PBS alone and in 1 M DMSO, and the latter occurred earlier than the former. As exhibited in Figure 3c, the PIR of PIECs in PBS alone and 1 M DMSO increased with increasing cooling rate (p < 0.05, n = 3). The PIFs of PIECs during thawing in 1 M DMSO showed an increase when compared with the cells in PBS alone (p < 0.05). The reason for this phenomenon may be that DMSO can bring more water into the cell (with the cell volume increasing) that is conducive to the formation of ice. The other reason is the greater cell dehydration in PBS. Usually, the presence of DMSO can inhibit IIF, but small amounts of DMSO could accelerate IIF. According to Mazur's two-factor hypothesis of freezing injury, IIF could cause mechanical damage to the cell membrane; meanwhile, the cell is cooled faster than optimum cooling rate and the cell is killed by IIF and its subsequent IIR during thawing. These experimental results indicate that the different cooling rates in the freezing process could impact not only the PIF but also the PIR in the thawing process.

IIR in PIECs from cryomicroscopy studies. The PIR in PIECs versus temperature at different cooling rates and at the same warming in the PBS alone
Assessment of cell morphology and cell viability
It is important to know that cell morphology changes in the absence/presence of 1 M DMSO during the freezing/thawing process. From Figure 4, we can observe typical cell morphology at some special temperatures, and meanwhile, it can be seen that the cell volume is gradually reduced during the freezing process and gradually increased during the thawing process, and the cell volume became bigger than the initial volume through the freezing and thawing process. In the presence of DMSO, the PIEC initial and ending volumes were bigger than PIECs in PBS alone due to the DMSO bringing more water into cell. There is a direct correlation between IIF and a disrupted cell membrane. 45

The cell morphology of PIECs in PBS
Furthermore, the PI was used for measuring cell survival in situ, and the results are shown in Figure 5. When the samples were incubated in PBS alone, the cells were dead after the freezing/thawing, no matter whether the cells encountered IIF and IIR or not (Fig. 5(a1)–(f1)). Meanwhile, when the samples were incubated in 1 M DMSO, a few cells were living after the freezing/thawing. As shown in Figure 5(a2)–(f2), the arrow indicates a cell that did not encounter IIF or IIR, and the cell was living after the freeze/thaw cycle.

Typical images of PIECs in PBS
To study further the impact of the freezing and thawing process on cell viability, the Hoechst 33342/PI fluorescent double-stain assay was used in this experiment. As shown in Figure 6a, the cells in the absence of DMSO were almost dead after the freezing/thawing cycle, but there were still a few surviving cells when the cells had been cultured in 1 M DMSO. From Figure 6b, the death rates of PIECs in PBS alone are higher than those in the presence of 1 M DMSO (p < 0.05, Student's paired two-tailed t-test, n = 5) and the death rates increased with the increasing cooling rates. This was because PIECs in the absence of CPA lost their membrane integrity and more water could move freely in and out of the cell, and finally, the cells swelled excessively and lost viability. In addition, the osmotic pressure around cells changed dramatically due to IIF, and too high or too low osmotic pressure could also cause irreversible injury. Although the PIFs of PIECs in the presence of the DMSO were higher than those in PBS alone, the DMSO could alleviate the osmotic injury during warming. Therefore, the death rates of PIECs in the presence of 1 M DMSO were lower than in PBS alone.
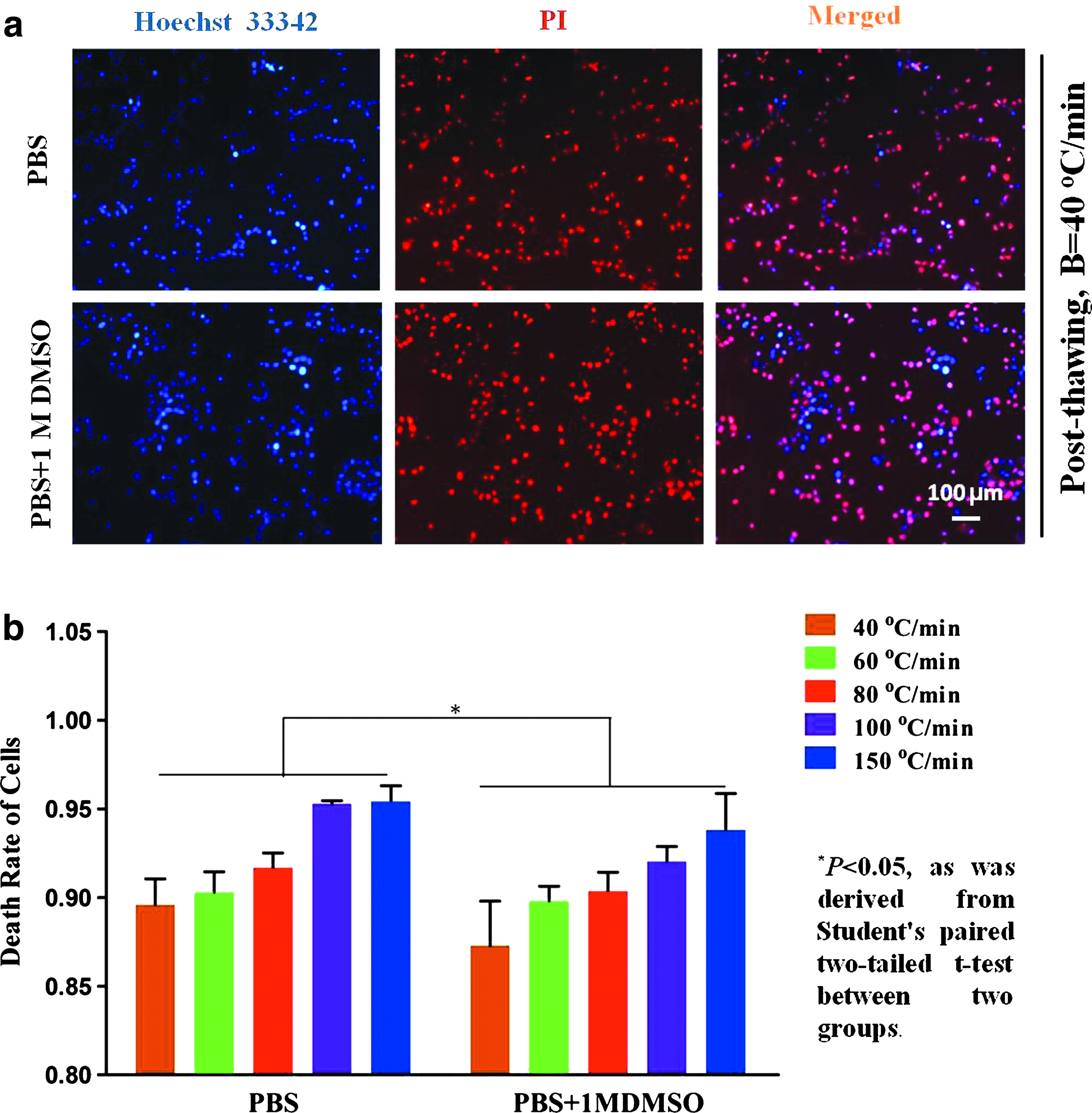
The viability of PIECs in PBS and in 1 M DMSO after freezing at 40°C/min and thawing at 100°C/min.
Conclusions
In this study, the PIF, PIR, and viability of PIECs were investigated experimentally, and quantification of IIR was displayed first in PIR. Our results showed that the PIF and PIR increased along with the increasing cooling rate and the same warming rate in PBS and in the presence of 1 M DMSO. It should be noted that during the cooling, IIF increases at higher subzero temperatures in PBS than in 1 M DMSO, although PIF at −60°C is somewhat greater. During warming, IIR occurs at lower temperatures. The most likely explanation is the greater cell dehydration in PBS, while the change in cell dehydration may also be attributed to the subsequent introduction of DMSO. The PIF, PIR, and viability of PIECs incubated in 1 M DMSO were higher than those in PBS alone, contrary to the common sense that larger PIF or PIR corresponds to greater cryoinjury. This may be attributed to the intrinsic protective effect of DMSO by reduction of solution injury or other potential injuries. It is common sense that the DMSO can inhibit IIF and IIR, but the presence of a low concentration of DMSO (1 M) could increase both IIF and IIR. These experimental results may be of potential application valuable for optimizing protocols for cryopreservation of the cell and cryosurgery.
Footnotes
Acknowledgment
This work was partially supported by grants from NSFC (Nos. 51276179, 51476160, and 51528601).
Author Disclosure Statement
No conflicting financial interests exist.