
Abstract
Modern genomic and metabolomic tools have provided the possibility of generating and interrogating large datasets that can provide answers to previously imponderable taxonomic, evolutionary, ecological, and physiological questions. However, the curatorial tools needed to provide and maintain the relevant biological resources on which new knowledge can be built have not kept pace with this meteoric rise in scientific capacity, its associated activity, or the huge increase in published science. The availability of biological material of guaranteed identity and quality in Biological Resource Centers is fundamental for scientific research, but it crucially depends on there being adequate preservation/maintenance methods that are capable of ensuring phenotypic, genotypic, and functional security of the biological material(s). This article highlights the challenges to the long-term maintenance of genetic resources in general, focusing specifically on the issues associated with the maintenance of a large collection of strains of the ecologically significant diatom Skeletonema marinoi. This research collection, held at the Department of Marine Sciences, University of Gothenburg, has been systematically tested for its capacity to survive cryopreservation. A method, involving incubation in the dark for 20–24 hours before cryopreservation, followed by cryoprotection employing 10% dimethysulphoxide (DMSO) and conventional cooling in a passive cooler, before plunging into liquid nitrogen was successfully applied to ∼80% of the strains tested. In addition, the growth characteristics of exemplar strains were confirmed after storage.
Introduction
M
The organizations hosting BRCs encompass a mixture of academic, public service and private, government, and commercial entities that have as part of their core remit the provision of characterized cultures as “seed” stocks:
• for use in research, • for teaching and training purposes, • for the development of industrial processes, • for the control of testing for quality assurance of products, • as referenced standards in the published scientific literature, • as reference strains for biological assays, • as reference material in biomedical assays, • as type and/or authentic strains for taxonomical studies, and • as centers for conservation of biodiversity.
It is widely agreed that the provision of cultures from BRCs is of little value unless these cultures are accompanied by information on their identity, provenance, and characteristics. 1 Thus, culture collections are important storehouses of information that can be accessed by direct contact with individual collections, or by contacting microorganism database organizations such as MIRCEN at http://wdcm.nig.ac.jp. Information resources can grow with time, as work on cultures from BRCs is published and increasingly individual collections provide more comprehensive data, images, bibliographic data, and links to EMBL/Genbank directly from their websites; for example, the Culture Collection of Algae and Protozoa (CCAP) 2 and the Belgian Co-ordinated Collections of Micro-organisms Diatom Culture Collection. 3 All of these services necessitate the long-term maintenance of “alive,” genetically, phenotypically, and functionally stable cultures. In the case of algae, maintenance by serial transfer has been the historic method of choice 4 and although there is evidence that some algae are stable in culture for many decades, 5 others have been reported to lose or modify key phenotypic characteristics, or metabolite production on prolonged maintenance by serial transfer. 6
With the exception of the Cologne, Coimbra, Roscoff, and UTEX culture collections, algal service culture collections all maintain less than 3000 strains in their open collections (Table 1), in comparison to bacterial or fungal collections, which in some cases hold tens of thousands of strains. Although almost all of these major collections now use cryopreservation as a curatorial tool, many algal taxa remain recalcitrant to conventional methodologies, and resource constraints (financial, staffing, and infrastructural) limit the number of strains that can be maintained by cultivation-based methods. This is a significant restriction to the types of studies that can be undertaken on algae, for example, projects that might generate 100s or even 1000s of individual isolates. Modern analytical approaches, including the development of molecular sequencing and their associated bioinformatic toolkits, have revolutionized the capacity to generate and interrogate large datasets. Furthermore, the increase in interest in the biotechnological exploitation of algae will inevitably result in the generation of thousands of mutants through selective breeding, conventional mutagenesis, as well as via genetic manipulation.
Algal, including the prokaryotic cyanophyta.
Protistan, numbers listed include holding of achlorophyllous heterotrophic protists.
Formerly the Culture Collection of Marine Phytoplankton (CCMP).
CCAP, Culture Collection of Algae and Protozoa.
Diatoms, one of the most species-rich groups of microalgae, are of key ecological significance and hold an interesting biotechnological potential, but they represent a particular challenge for long-term preservation. Due to their gradual reduction in cell size, vegetatively dividing clones cannot be kept in culture for extended time by serial transfer, as cells lose viability beyond a species-specific minimal cell size. Experimentally inducing sexual reproduction is one way to maintain species (but not genotypes) in culture, but it is not possible for many species. In particular, in the case of centric diatoms, the sexual cycle is poorly understood and controlled sexual reproduction is currently in its infancy. Precious strains, possessing unique phenotypic characteristics, or those originating from archived sediment samples are easily lost in culture. 7 Furthermore, in the case of genetically transformed strains, there is a high risk of losing insertions if strains are maintained asexually. 8
The challenges cited earlier provide a stimulus for exploring the options available to conserve large collections of strains belonging to the same taxon. Of the long-term preservation approaches employed in BRCs, only cryopreservation (storage at ultra-low temperatures) has the potential to provide a practical solution to these challenges. In addition, this approach has the advantage that, after the initial preservation procedure has been performed, routine maintenance costs are limited to costs of refrigerant and viability can effectively be guaranteed for decades, if not hundreds of years. 9
In this study, the centric diatom Skeletonema was selected, because it is an important contributor to the pelagic community and often dominates phytoplankton assemblages in temperate coastal regions. It is a widespread genus, except in the polar regions, 10 and to date, 13 species have been described.11–14 In Scandinavian waters, so far only one species, Skeletonema marinoi Sarno et Zingone, has been reported15,16 and it is especially abundant during spring bloom, when densities of millions of cells per liter occur.
The predominant means of propagation is through vegetative division, but the formation of auxospores and sexual reproduction has been documented. 17 The generation time is 24 hours, which makes it ideal for studies of phenotypic response. Furthermore, benthic cells act as a resting stage; they can survive for at least 100 years and provide a short-term evolutionary archive in sediments. 7 S. marinoi is easy to collect, isolate, and maintain in culture, and the survival of monoclonal cultures after single-cell isolation is almost 100%.
Skeletonema is routinely used as a model in ecological and other studies18–20 and at present, hundreds of strains are held in the Göteborg University Marine Algal Culture Collection (GUMACC) as an “in house” research resource. Like all such collections, the maintenance of the resource is a challenge with respect to its curatorial costs and the long-term conservation of the materials cannot be guaranteed. Initially, a pilot study was undertaken on three well-documented strains. This was subsequently expanded to test the suitability and robustness of a standardized protocol across the holdings of the GUMACC.
Materials and Methods
Isolates studied and cultivation conditions
Skeletonema marinoi CCAP 1077/5 was obtained from the Culture Collection of Algae and Protozoa (CCAP). All other isolates studied were from the Göteborg University Marine Algal Culture Collection and their origin, isolation date, and where available Genebank barcode data are listed in Table 2. All the strains were grown in 50 mL Nunc Nunclon™Δ EasYFlasks™ with a vent closure permitting gas exchange, at irradiance ca. 40 μmol photons m2 s−1 (measured at lid height and provided by fluorescence tubes, L36W/865 Lumilux® Cool Daylight, Osram GmbH, Augsburg, Germany), with a 12 hour:12 hour light-dark photoperiod at ca.10°C. Growth medium was prepared with filtered seawater autoclaved and enriched with nutrients according to the standard recipe for f/2 medium 21 plus sodium metasilicate.
Cryopreservation methods
Controlled rate cooler methods (Methods A–D)
A 10% v/v solution of DMSO 99.9% (Sigma-Aldrich Ltd., United Kingdom) was prepared and filter-sterilized. A 2.5 mL aliquot of the 10% (v/v) DMSO solution was added to 2.5 mL of late-log phase cells to give a final DMSO concentration of 5% (v/v). Aliquots (1.0 mL) were dispensed into cryovials (Greiner Bio-One GmbH, Frickenhausen, Germany). These were then incubated at room temperature (ca.20°C) for 15 min before cryopreservation, to ensure that the cryoprotectant (CPA) had entered the cells. The cryovials were then transferred to the cooling chamber of a Planer Kryo 360-3.3 (Planer, United Kingdom) programmable freezer and cooled from room temperature at −1°C min−1 (Method A), −5°C min−1 (Method B), or −10°C min−1 (Method C) to −40°C and then plunged into a dewar of liquid nitrogen (LN2) as previously detailed by Day and Deville. 25
Alternatively, samples were cooled by employing the standard protocol used at the National Center for Marine Algae and Microbiota (NCMA) (Method D), as detailed by Day and Brand (2005). 26 Aliquots (0.5 mL) of cryoprotected culture, as outlined earlier, were held at room temperature (ca. 20°C) for 15 min. Then, the cryovials were transferred to the Planer Kryo 360-3.3 programmable freezer and cooled at −1°C min−1 to −9°C, then at −20°C min−1 to −45°C, then warmed at +20°C min−1 to −12°C, then cooled at −1°C min−1 to −45°C, then cooled at −20°C min−1 to −90°C, and finally plunged into LN2.
The samples were then transferred to a cryostorage container, and the location was logged on the CCAP cryostorage database for overnight storage before being thawed.
Passive freezer methods (Methods E and F)
Method E: CPA solutions and cultures were prepared as outlined earlier, and 1 mL aliquots were aseptically transferred into pre-labeled cryovials. The Passive freezer unit, Mr. Frosty™ (Nalgene NUNC International, Rochester, NY) was prepared as per instructions, with the addition of 250 mL of isopropanol to the reservoir adjacent to the cryovials tube holder and was then placed in a refrigerator overnight to equilibrate at ∼4°C. The cryovials containing CPA-treated algae were transferred into the Mr. Frosty (which has been designed to have a cooling rate of ca. −1°C min−1), and the unit was sealed and placed in a −80°C freezer. This was then held in the freezer for 1.5 hours, by which time the temperature of the contents of the vial was less than −50°C. The frozen samples were then plunged into LN2 and transferred to a cryostorage container, and the location was logged on the CCAP or GUMACC cryostorage database for overnight storage before being thawed.
Method F: An alternative approach, previously successfully applied in the BCCM (Chepurnova and Vyverman, unpublished data), was also employed. This involved placing late-log phase S. marinoi cultures in complete darkness for 20–24 hours before cryopreservation. The Mr. Frosty unit was prepared as described earlier. A 40% (v/v) solution of DMSO was prepared, in the culture medium used to grow the alga, and filter sterilized. Then, for each strain, 6 mL of culture and 2 mL of 40% DMSO were pipetted into 15 mL Falcon tubes to give a final CPA concentration of 10% (v/v). The tubes were inverted several times to mix the CPA uniformly. The cultures with CPA were then transferred into pre-labeled 2 mL cryogenic vials and left to equilibrate at room temperature in darkness for at least 20 minutes. The cryogenic vials were then transferred into the pre-chilled Mr. Frosty and placed in an −80°C freezer for 90 min, as outlined earlier. The frozen samples were then plunged into LN2 and transferred to a cryostorage container, and their locations were logged on the CCAP or GUMACC cryostorage database for overnight storage before being thawed.
Thawing and culture recovery
The frozen cryogenic vials were rapidly transferred to a 37°C water bath. The vials were placed in floating vial holders and were gently agitated by hand to facilitate thawing. As soon as all the ice had melted, 1 mL from each vial was immediately transferred into a 250 mL flask containing 50 mL of fresh culture medium. This was then mixed to ensure homogeneous resuspension of the culture and reduction in the concentration of the CPA to <0.2%. The cultures were allowed to recover in complete darkness overnight and then incubated at 10°C, under a 12 hour:12 hour light-dark photoperiod (40 μmol photons m2 s−1). Samples were monitored for up to 48 days post-thaw, and fresh medium was added to the culture as soon as growth was observed.
Viability staining
Carboxyfluorescein diacetate succinimidyl ester (CFSE) is a non-fluorescent, membrane permeable molecule. When inside a viable cell, due to enzymatic modification, CFSE becomes highly fluorescent; in addition, it losses permeability to the cell membrane, and it can remain detectable inside the cell for several days. 27 CFSE in the form of a 5 mM stock solution in DMSO was freshly prepared and added to the samples to give a final concentration of 2 μM. The CFSE-labeled cells were incubated at room temperature for 15 minutes before examination under an Axioskop 2 epifluorescence microscope, and micrographs were taken under fluorescence and phase contrast at 400x magnification. Viability assessment was estimated, by calculating the percentage of CFSE positives (live cells, stained in green) out of a total of ca. 50 cells observed per sample. This was performed after 24 hours of incubation 26 to reduce the number of false-positive cells that were damaged by the procedure, but despite not being able to recover, still retained metabolic capabilities immediately after thawing.
Regrowth assays
Comparisons were made on the maximum growth rates, during the exponential phase, and both before and after cryopreservation to investigate possible phenotypical changes. The growth rates were obtained spectrophotometrically by using optical density (OD, λ 600 nm).
Three 250 mL culture flasks for each strain were filled with 160 mL of culture containing a concentration of 5000 cells mL−1. The flasks were kept in a controlled environment (10°C, under a 12 hour:12 hour light-dark photoperiod ca. 40 μmol photons m2 s−1) for 15 days. Each day, 700 μL from each sample was withdrawn onto a 48-well plate and density was estimated by OD using Varioscan Flash plate reader (Thermo Scientific). Post-cryopreservation, samples were taken identically as pre-cryopreservation samples from the surviving cultures.
Statistical analysis
Growth rate (μmax), defined as instantaneous rate of increase, was based on the longest possible period of exponential growth for each strain pre- and post-cryopreservation according to: μmax = (lnODt-lnODt-1)/Δt, where t = time, and OD reading at 600 nm. 28 The interval of exponential growth was determined from growth curves established from each experimental culture replicate. The statistical difference between pre- and post-cryopreservation growth was assessed with a t-test.
Results
A pilot study was undertaken at the CCAP by using a CCAP strain (S. marinoi CCAP 1077/5) and two GUMACC: one strain isolated from a sediment sample (S. marinoi St54) and the second from a water sample (S. marinoi 0603AA) to test five commonly employed protocols. These methods utilized either a controllable programmable cooler, which allows the cooling rate to be varied mechanically, or a passive cooler with a nominal cooling rate of ca. −1°C min−1. Post-thaw viability levels were assessed by light microscopy and vital staining was assessed by using fluorescence microscopy (Fig. 1), and the capacity to regenerate “normal” cultures. Cells with ultrastructural damage induced by cryopreservation were readily identified by using phase-contrast microscopy, and vital staining confirmed that they were metabolically inactive and/or had a ruptured cell membrane. On the basis of the vital staining data, all methods tested were effective for the CCAP strain, with high levels (64%–85%) of viable/metabolically functional cells being observed and revival of all the cultures (Table 3). Levels of post-thaw viability, on the basis of vital staining, were much lower for the GUMACC strains (0%–39%) and controlled rate cooling at 1°C min−1 using the standard CCAP protocol resulted in the best recovery with all three replicated regrowing for S. marinoi St54 and two of the three replicates of S. marinoi 0603AA regenerating normal cultures (Table 3). It was noted that the vital staining method of viability assessment was not robust and that although high levels of CFSE-positive cells were observed 24 after thawing, for cultures that failed to recover, these levels fell on subsequent incubation under standard cultivation conditions and after prolonged incubation no morphologically normal or CFSE positively stained cells were observed.
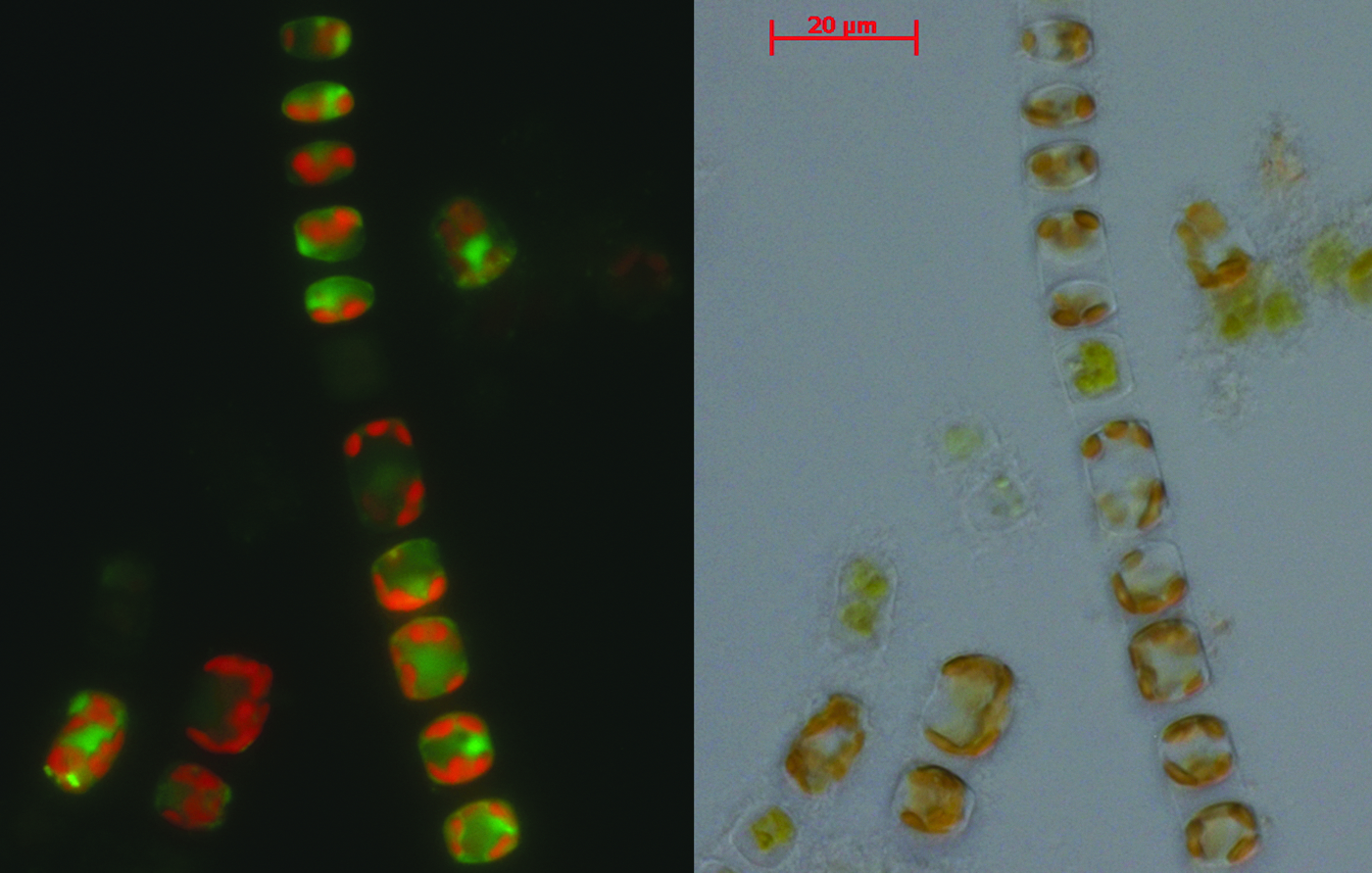
Viability assessment by microscopy. Epifluorescence and phase-contrast micrographs of Skeletonema marinoi. Live cells stained green with CFSE correlate with morphologically normal cells under phase contrast. Dead and damaged cells remain unstained, or where cells were intact appear red due to auto-fluorescence of chlorophyll. CFSE, carboxyfluorescein diacetate succinimidyl ester.
Regrowth (+), or failure to grow (−), cells died and lysed, in replicate thawed samples. Number in parentheses is the mean level of post-thaw viability on basis of vital staining and cell appearance 24 hours after thawing.
Cooling rate (°C min−1) to intermediate holding temperature.
CRC, controlled rate cooler.
As part of the EU-funded ASSEMBLE project Trans National Access program, a cryopreservation campaign was undertaken at the CCAP. In total, 10 batches of four GUMACC strains with 15 replicates per strain were cryopreserved and banked in the CCAP cryobank. The logistics of applying a campaign of cryopreservation to the GUMACC collection worked well, but subsequent recovery regrowth was much poorer than anticipated. Recovery of all three replicates, within 4 weeks, was only observed in two strains (S. marinoi St54 and S. marinoi GF04-3A). However, on prolonged incubation, full recovery was observed in two additional strains (S. marinoi 0603AA and S. marinoi Oslo AE). Of the remaining 36 strains, recovery of two replicates was observed for four strains and recovery of one replicate for a further seven strains after prolonged incubation, with the remaining 25 strains tested failing to show any capacity to regrow (Fig. 2).
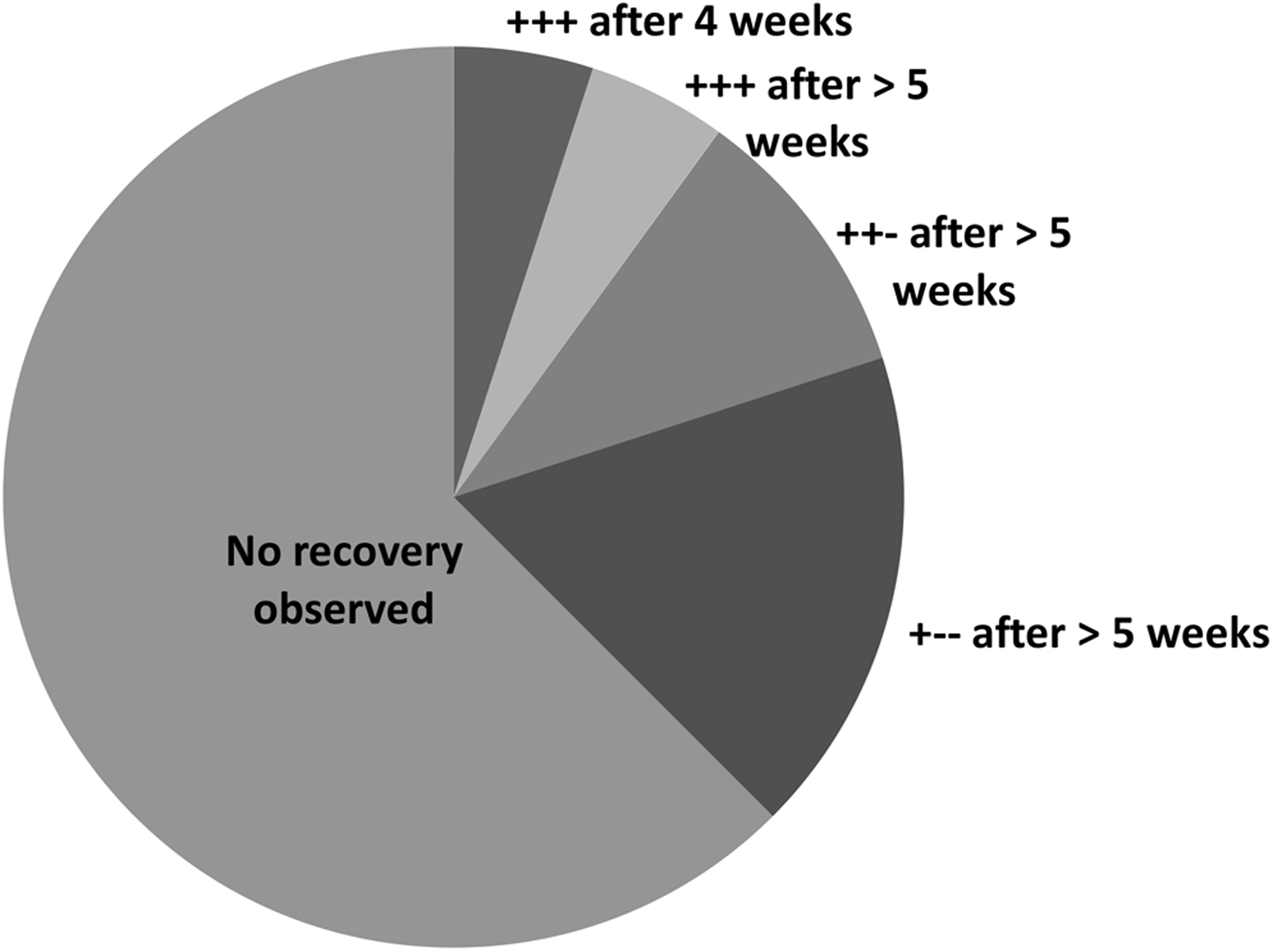
Application of the standard CCAP two-step controlled rate cryopreservation protocol (Method A) to 40 GUMACC S. marinoi strains. Regrowth (+), or failure to grow (−), in replicate thawed samples. CCAP, Culture Collection of Algae and Protozoa.
Controlled Rate Cooling equipment is not available at the GUMACC; therefore, further optimization of this approach “in house” was not practical, so improvement of the Passive cooler approach was prioritized. Using a protocol trialed at the BCCM, post-thaw viability was improved for the reference genome strain S. marinoi St54, with all three replicates tested showing recovery and the time taken to regenerate a healthy culture reduced from 40 to <30 days (Table 4). Furthermore, the method was applicable to strains that had failed to recover when tested using the CCAP passive cooling protocol, including: S. marinoi A22, S. marinoi HAKH, and S. marinoi R05AC (Table 4). This approach has been tested for a total of 17 S. marinoi strains, with 14 showing recovery and only three strains failing to grow on thawing (Table 4). The stability/growth performance after employing this cryopreservation method was assessed for nine S. marinoi strains. On assessing algal biomass daily, based on OD (600 nm), no significant difference in maximum growth rate (μmax) before and after cryopreservation was observed in any isolate tested, with the exception of S. marinoi HAKH, which displayed significantly lower μmax after cryopreservation (p = 0.01) (Fig. 3). Furthermore, these cultures were subsequently maintained by serial transfer for >6 months, with transfers every 3–4 weeks without any observable change to the cells or their growth characteristics.
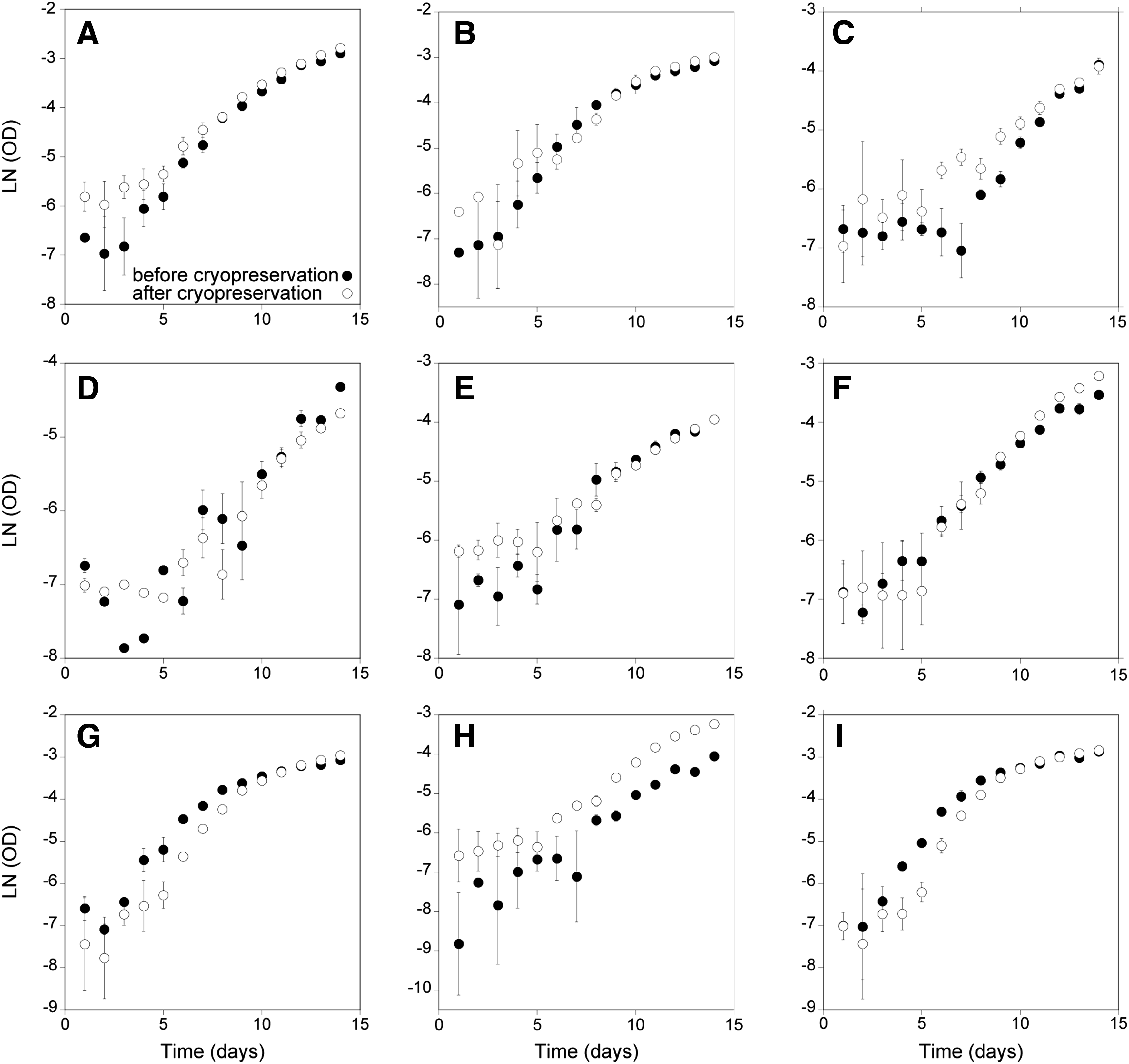
Growth curves of Skeletonema marinoi strains before (black) and after (white) cryopreservation. Algal biomass was estimated daily based on optical density (600 nm). There was no significant difference in maximum growth rate (μmax) before and after cryopreservation except for strain HAKH, which displayed significantly lower μmax after cryopreservation (p = 0.01).
Regrowth (+), or failure to grow (−), in triplicate thawed samples.
NT, not tested.
Discussion
Science built on the use of live materials, whether fundamental or applied, has an absolute requirement for the biological materials employed to retain reproducible characteristics. This need for reproducibility has profound implications for those who use, hold, and/or distribute microbial cultures, including microalgae. A traditional example of this is in taxonomy, where the botanical taxonomic community, including those who work with algae, have a requirement for type materials (i.e., the originally described organism to which the scientific name has been attached) that do not change. Indeed, this is a prerequisite in the International Code of Botanical Nomenclature. 29 Most commonly, type specimens are held as preserved specimens, in the case of diatoms as permanent slides, or alternatively, holotypes such as illustrations published in the hard-copy scientific literature. However, since the year 2000, fungi and algae, “if preserved in a metabolically inactive state (e.g., by lyophilization/freeze-drying, or deep-freezing), are acceptable as types.” 30 It has subsequently been suggested in the era of genomics and metabolomics that alternative strategies, such as cryopreservation at ultra-low temperatures that allow the possibility of restoration of metabolism, could be used to conserve type materials and that this would allow more comprehensive scientific exploitation of the type materials. 31 In addition to taxonomy, ecological, physiological, and biotechnological orientated users of biological resources all have a requirement for “biological standards” and optimally, these should be held/maintained in such a way as to minimize any genotypic, phenotypic, or functional change in the organism conserved. 32
In total, more than 2.4 million microbial strains are held by BRCs registered at the World Data Centre for Microorganisms (WDCM). The vast majority of these holdings are bacterial and fungal and although there are exceptions, the use of long-term conservation methods, such as lyophilization, or cryopreservation, is the norm. For algae, although freeze-drying may be applicable for some cyanobacteria, 33 this method results in unacceptably low levels, or no viability when used for eukaryotic algae.26,34 However, cryopreservation has increasingly become a standard approach in larger algal culture collections (Table 1), and methodologies have been developed for a wide range of algal taxa.26,35
A further stimulus to the need for cryopreservation methodologies for algae is the logistic and resource implication of routine maintenance. The number of strains held in the major algal collections is constrained by the cost and staffing needs, and in recent years, these collections have stabilized in size or, in some cases such as the CCAP, reduced slightly as more strains have been lost through culture failure and Q/A exercises than have been accessed. Furthermore, significant expansion of ecologically oriented collections like the GUMACC and their long-term maintenance, without guaranteed long-term core funding, are extremely challenging.
New large projects necessitating accession of multiple isolates, or the development of genetically transformed algae, which generate 1000s of transformed clones,36,37 are equally problematic and the instability of some transformed algae, including the model diatom Phaeodactylum tricornutum, 37 means that conventional ex situ maintenance by serial transfer is inappropriate and not sustainable.
Cryopreservation provides a methodology that is capable of holding preserved material in effective “suspended animation” so that their genetic or physiological potential remains unchanged. There is a growing literature (evidence based) on this in algae where the capacity to retain genetic integrity, morphology, metabolic functionality, and growth characteristics has been reported in organisms that have been revived from cryopreservation.8,38–40 For diatoms, this is particularly relevant, as on repeated serial transfer of inoculum, irreversible shrinkage can occur in some species 41 and this has been a major constraint on their use in sequential scientific publications, as well as the capacity of collections to maintain them in culture and to provide identical strains to different research groups. Thus, particularly in studies, or applications, where stability is critical to performance, cryopreservation provides the most practical solution to ensuring a “fit for purpose” inoculum is employed.
As outlined earlier, there is an expanding published literature on the cryopreservation of algae. In the case of diatoms, much of the emphasis has been on methodological development, particularly of taxa with biotechnological or aquaculture relevance.42–44 These papers explored the influence of key parameters, including culture age, CPA choice, cooling rate etc. with the objective of developing methodologies that are applicable to a range of algal taxa, including diatoms. Other studies have focused specifically on diatoms exploring both the cryobiological implications of subjecting diatoms to ultra-low temperatures and how this knowledge may be employed to improve post-thaw viability and cell recovery.45–48,49
To date, there have been no publications exploring the logistics of attempting to conserve a large diatom collection by cryopreservation. However, Morris 49 previously reported the successful application of a conventional two-step cryopreservation method to >250 members of the Chlorococcocales. The method employed was similar to the optimal approach used in this study insofar as it utilized a relatively cheap, constant temperature alcohol bath (Fryka model KB 3000) maintained at a sub-zero temperature, with subsequent immersion into liquid nitrogen. In this study, despite the possibility of more strictly controlling the cooling profile, the methodology that had been optimized in a pilot study for three S. marinoi strains proved unsuccessful when applied to a much larger number of strains, with only approximately 35% of the strains tested surviving. This was unexpected, as one might have anticipated on the basis of prior experience25,26,35 that slow cooling using a controlled rate cooler (CRC) would result in more success than the alternative use of less controlled passive coolers. The standard CCAP Passive cooler cryopreservation protocol 26 was found to be effective for some strains, for example, S. marinoi GB3, with recovery being observed in all culture replicates, but unsuitable for other strains with no recovery observed after up to 48 days post-thaw incubation. However, employing higher levels of CPA, 10% in the BCCM method (Method F), versus 5% in the CCAP method (Method E) resulted in survival and regrowth in >80% of strains tested. The use of higher levels of DMSO has been observed in the BCCM to improve survival in a range of marine diatoms, with concentrations in the range of 10%–15% being optimal. This has also been observed for S. marinoi CCMP2092, which has been cryopreserved by using the standard NCMA/CCMP cryopreservation protocol (Method D) using 15% DMSO as CPA. 50 The BCCM protocol also has the advantage of practicality and involves use of standard lab equipment, rather than an expensive specialized cooling unit. For most practitioners, particularly those in smaller laboratories where there is no access to specialized cryobiological cooling devices, the use of low-tech approaches is the only realistic option. Here, a conventional two-step, colligative cryopreservation approach was used successfully, with good recovery in most strains and acceptable culture performance in terms of growth characteristics on thawing. An alternative strategy that was not explored in this study is the use of vitrification, whereby cells are subjected to treatments that result in high levels of intracellular dehydration and then ultra-rapid cooling, which leaves samples in a vitrified glassy state. This has been successfully used to cryopreserve the marine diatoms Nitzschia closterium and Chaetoceros muelleri using pre-treatment with a high osmotic potential solution (PVS2), followed by encapsulation in calcium alginate beads and rapid cooling in liquid nitrogen. 51 Such methodologies have the potential to be more widely employed. However, from a practicality perspective, they have a greater number of handling steps, with their potential for error and contamination. Furthermore, conventional vitrification solutions, such as PVS2, are known to be toxic to a range of algae, so encapsulation dehydration methodologies using sucrose as a vitrification solution and/or air flow dehydration have been found to be more applicable to a range of microalgae.52–54
In conclusion, there is an increasing demand for the maintenance of both a wider diversity of taxa and many more cultures of individual microalgal species. The traditional approach of employing serial transfer and maintenance under conditions that extend the culturing interval 4 is no longer capable of meeting this challenge, without significant investment in infrastructure and manpower. Only cryopreservation has the potential as a long-term conservation approach that is suitable for maintaining viability and functionality. Choice of methodology is crucial, as methods need to be robust, reproducible, and applicable to as large a number of strains as possible. The designing of individual methodologies for each isolate is neither practical nor efficient, so compromises need to be accepted with regards to what is an appropriate viability level or how long a period is required to regenerate a healthy normal culture. The choice of method and cooling equipment is dictated by resource availability, but in general, standardized simple methodologies that do not require specialized equipment will be more readily adoptable by the user community.
Footnotes
Acknowledgments
This work was supported by the European Commission (ASSEMBLE, Integrated Infrastructures Initiative, grant agreement no. 227799). JGD, OC, and WV acknowledge funding from BRAIN-be for the PRESPHOTO project. In addition, AG acknowledges funding from the Swedish Research Council FORMAS (217-2012-692).
Author Disclosure Statement
No conflicting financial interests exist.