
Abstract
The addition of antioxidants to semen cryopreservation extenders has been employed for combating oxidative damage. This work aimed to evaluate the addition of carotenoid canthaxanthin to a cryopreservation extender of ram semen. Three breeder rams were used and, after semen collection, with 48-hour intervals between collection, the samples were included in the pool formation (n = 6). The experimental groups comprised 0 (control), 0.1, 1, 10, and 25 μM of canthaxanthin. After thawing (37°C/30 s) and incubation at 37°C for 2 hours, semen aliquots from each group were evaluated for sperm kinetics (CASA), the integrity of the plasma and acrosomal membranes (iPAM), intracellular reactive oxygen species (ROS) production, and lipid peroxidation (LPO) by flow cytometry associated with the image. The control group and canthaxanthin 1 μM after incubation at 37°C for 2 hours showed increases of curvilinear velocity and amplitude of lateral head displacement with decreases of linearity, straightness, and wobble (p < 0.05), which were not observed for the canthaxanthin 10 and 25 μM. The supplementation of a Tris-egg yolk extender with canthaxanthin had no effect on the iPAM, intracellular ROS production in viable spermatozoa, or LPO. In conclusion, supplementation with 10 and 25 μM of canthaxanthin in a Tris-egg yolk extender used for ram semen cryopreservation is able to protect ovine sperm from kinetic changes after incubation at 37°C for 2 hours post-thawing.
Introduction
T
However, this feature makes spermatozoa vulnerable to lipid peroxidation (LPO) because the PUFAs are susceptible to oxidative attack, resulting from the action of reactive oxygen species (ROS), which causes loss of motility, damage of the plasma and acrosomal membranes, apoptosis, and cell death.6,7 Although spermatozoa need ROS to perform their physiological functions, such as sperm capacitation, hyperactivation, acrosome reaction, and interaction with the oocyte,8–10 the imbalance between the production of ROS and the antioxidant cell system can lead to oxidative stress.11,12
Cryopreservation, in turn, exposes the sperm cells to osmotic stress during the freezing and thawing processes, which leads to oxidative stress due to excessive ROS production. 13 It has been demonstrated that the ram semen subjected to cryopreservation suffers an increased production of ROS responsible for the loss of motility, LPO of the membrane, and DNA damage, thereby decreasing the fertilization capacity.14–16
Several studies have reported the ability of different antioxidants, which are added to the freezing extenders of ram semen to protect sperm from oxidative damage, but varied results have been reported on sperm quality after thawing.17–20
Carotenoids are natural pigments that are considered essential in several important biological processes, such as the regulation of membrane fluidity 21 or their function as antioxidants.22,23 Canthaxanthin (β, β-carotene 4,4′ dione) is a carotenoid found in plants, green algae, bacteria, crustaceans, fish, and birds. 24 Some reports have supported the protective effects of canthaxanthin as an antioxidant in cell cultures in vitro.25,26 Moreover, it has been shown that canthaxanthin supplementation in the diet of breeder hens improved the antioxidant capacity of tissues, reducing susceptibility to LPO.27–29
However, no previous studies have reported the effects of canthaxanthin on the functional properties of mammalian sperm. Therefore, this study aimed to investigate the effects of canthaxanthin added to a Tris-egg yolk extender on the variables, sperm kinetics, integrity of plasma and acrosomal membranes, intracellular ROS production, and LPO of the plasma membrane, in ram semen after cryopreservation.
Materials and Methods
Reagents
The reagents used in this study were purchased from Sigma-Aldrich (St. Louis, MO), except those specified. Canthaxanthin was diluted with 1 mL of DMSO heated to 37°C, yielding a stock solution of 10 mg/mL, which was maintained at −20°C until use.
Animals
All procedures involving animals were conducted with the approval by the Ethics Committee for Animal Experimentation of the Universidade Federal Rural de Pernambuco—UFRPE (Brazil), under license number 046/2015 CEUA/UFRPE.
Three Santa Ines rams aged between 2 and 3 years and with a history of fertility were used, receiving tifton hay, commercial concentrated feed (400 g/animal/day), water, and mineral supplementation ad libitum as feed. The animals were kept in the Department of Veterinary Medicine of UFRPE, Pernambuco, Brazil (08° 03′14″ S, 34° 52′ 52″ W).
Semen collection and raw semen evaluation
Six semen collections per animal at intervals of 48 hours were performed for a total of 18 ejaculates. The ejaculates were obtained by the artificial vagina method using a female as dummy. Immediately after collection, the semen samples were evaluated subjectively in an optical phase-contrast microscope (100 × ; Olympus, Tokyo, Japan) for mass movement and motility. Each ejaculate was evaluated separately, and only ejaculates with mass movement and motility values ≥3% and ≥70%, respectively, were used for the pool formation (n = 6).
After the pool formation, the sperm concentration was determined using a Neubauer chamber. Then, the sperm morphology was evaluated by the moist chamber method at 400× magnification, counting 200 cells per slide. Abnormalities were classified as a percentage of normal cells and major and minor defects. 30 The pools were approved when they presented the values of sperm concentration ≥1 × 109 spermatozoa mL−1 and sperm pathologies ≤20%.
Semen cryopreservation
The semen pool was diluted in the Tris-egg yolk extender (375 mM Tris, 124 mM citric acid, 41.6 mM fructose, 20% egg yolk, 5% glycerol, pH 6.8) 19 to a final concentration of 200 × 106 spermatozoa mL−1. After dilution, the sample was divided for formation of experimental groups, into which canthaxanthin was added for five different final concentrations: 0 (control), 0.1, 1, 10, and 25 μM. The final DMSO concentration in all groups was 0.05%, including the control group. The samples were packaged in 0.25-mL straws, identified, and frozen in an automated system (TK Tecnologia em Congelação Ltd., Uberaba, Brazil), beginning at room temperature (27°C) and using a refrigeration curve of −0.25°C/min up to 5°C. The semen was subjected to a stabilization period of 120 minutes, and immediately after this period, a freezing curve of −15°C/min was initiated until −120°C was reached. Then, the straws were stored in liquid nitrogen (−196°C) until the time of analysis.
Thawing and semen evaluation
Two straws of semen from each experimental group were thawed (37°C/30 s) and grouped. The experimental groups were evaluated at two periods: immediately after thawing and after 120 minutes of incubation at 37°C. Six replicates for all experimental groups were performed.
Kinetic assays
Immediately after thawing and after 120 minutes, a semen aliquot (10 μL) was diluted with a Tris-egg yolk extender without glycerol (375 mM Tris, 124 mM citric acid, 41.6 mM fructose, 20% egg yolk, pH 6.8) preheated (37°C) to a cencentratrion of 50 × 106 spermatozoa mL−1. The evaluation of sperm kinetics was performed with a computerized system for sperm analysis (CASA, SCA™; Microptics, SL, Version 5.1, Barcelona, Spain). An aliquot of 2.5 μL of sample was placed on a slide and covered with a coverslip (18 × 18 mm); both were preheated at 37°C and evaluated by phase-contrast microscopy (Eclipse 50i, Nikon, Japan). Images of the sperm cells were captured using a video camera (Basler Vision Tecnologie™ A312FC, Ahrensburg, Germany) at 100× magnification. For each sample, five random and nonconsecutive fields were recorded where at least 500 sperm cells were captured.
The following variables were assessed: total motility (TM; %), progressive motility (PM; %), straight-line velocity (VSL; μm/s), curvilinear velocity (VCL; μm/s), average path velocity (VAP; μm/s), beat-cross frequency (BCF; Hz), amplitude of lateral head displacement (ALH; μm), straightness (STR; %), linearity (LIN; %), and wobble (WOB; %). The parameters of the CASA system were measured with the following settings: temperature, 37°C; magnification, 100 × ; number of images, 25; frames per second, 25; area of the head, 20–70 μm2; VAP: slow 10 μm/s < medium 45 μm/s < rapid 75 μm/s; progressiveness, 80% STR; and circular, 50% LIN.
Flow cytometry assays
The procedures described below were performed to analyze the integrity of the plasma and acrosomal membranes (iPAM), intracellular ROS production, and LPO of the plasma membrane. In each analysis, aliquots of 40 μL of semen (200 × 106 spermatozoa mL−1) were diluted in 1 mL of PBS (phosphate-buffered saline, Dulbecco's Ca+2 and Mg+2 free) and centrifuged (600 g/10 min) to remove the extender and cellular debris. After the supernatant was discarded, the sperm pellet was resuspended in 40 μL of PBS.
For iPAM evaluation, the samples were stained by addition of 1.0 μL of fluorescein isothiocyanate–peanut agglutinin (FITC-PNA; 200 μg/mL in PBS) and 2.0 μL of propidium iodide (PI; 0.5 mg/mL in PBS), fixed with 5.0 μL of 4% paraformaldehyde, incubated for 10 minutes at room temperature, and evaluated.
For the LPO analysis, 1.0 μL of C11-BODIPY581/591 (2 mM in DMSO, Molecular Probes, Life Technologies, Eugene) was added, and for the intracellular ROS production analysis, 2.0 μL CM-H2DCFDA (0.5 mM in DMSO; Molecular Probes, Life Technologies, Eugene, Oregon) was added. Both samples were incubated at 37°C for 30 minutes, subsequently diluted with 1.0 mL PBS, and centrifuged (600 g/10 min) to remove the excess stain probe. The pellets were resuspended in 40 μL of PBS, fixed with 5.0 μL of 4% paraformaldehyde, incubated for 10 minutes at room temperature, and then evaluated. In the intracellular ROS production analysis, before the addition of paraformaldehyde, 1.0 μL PI was added (0.5 mg/mL).
The cytometric analyses were performed using an ImageStream®x Mark II cytometer (EMD Millipore Corp., Seattle, Washington) equipped with a microscope with a 60× objective (0.5 μm pixel resolution) with an image rate of 5000 cells/s. The cell size was 7.0 μm, and the flow velocity was 44 mm/s. The samples were analyzed with a laser of 488 nm with an intensity set to 55.0 mW (iPAM analysis), 60.0 mW (LPO), and 80.0 mW (ROS production), and 5000 events for all samples were collected. Bright-field imagery was collected using an LED-based bright-field illuminator. FITC-PNA and CM-H2DCFDA were collected on channel 2 (505–560 nm), and PI was collected on channel 5 (640–745 nm). The images of C11-BODIPY581/591 were collected in channels 2 and 4 (595–640 nm). To acquire raw images, INSPIRE® software was used, and the image analysis was performed using the IDEAS® 6.0 image analysis software.
The following cells were considered: cells unstained with intact acrosomal and plasma membranes (PNA−/PI−), cells with damaged acrosomal membranes and intact plasma membranes stained only by FITC-PNA (PNA+/PI−), cells with intact acrosomal membranes and damaged plasma membranes stained only by PI (PNA−/PI+), and double-stained cells with damaged acrosomal and plasma membranes (PNA+/PI+) (Fig. 1). In the analysis of intracellular ROS production, cells with low ROS levels and intact plasma membranes were those that were not stained (DCFDA−/PI−), cells with high ROS levels and intact plasma membranes were stained only by CM-H2DCFDA (DCFDA+/PI−), and cells stained by PI presented damaged plasma membranes (Fig. 2). For the LPO in membranes, cells stained in orange did not show peroxidation, whereas cells stained in green show peroxidation (Fig. 3).
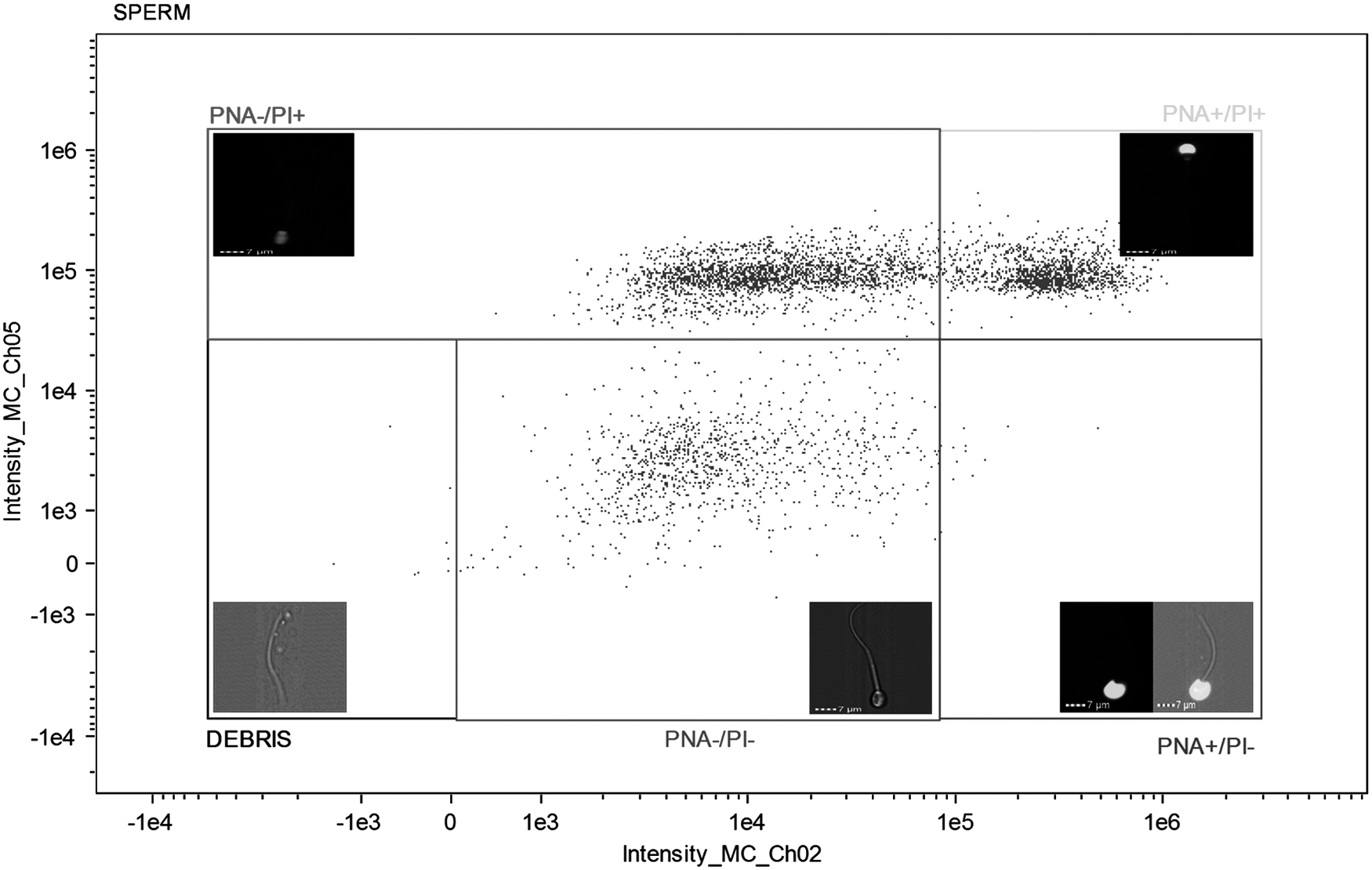
Plasma and acrosomal membrane integrity of ram semen after freezing–thawing in semen extender supplemented with canthaxanthin antioxidant. DEBRIS (extender waste and cellular debris tails); PNA−/PI− (cells with intact acrosomal and plasma membranes, unstained); PNA+/PI− (cells with damaged acrosomal and plasma membrane intact, only stained by FITC-PNA); PNA+/PI+ (cells with damaged acrosomal and plasma membranes, double-stained cells); PI+ (cells with damaged plasma membrane, only stained by PI). PI, propidium iodide.

Oxidative status of ram semen after freezing–thawing in semen extender supplemented with canthaxanthin antioxidant. DEBRIS (extender waste and cellular debris tails); DCFDA−/PI− (viable cells without oxidative stress, unstained); DCFDA+/PI− (viable cells with oxidative stress, only stained by DCFDA); PI+ (cells with damaged plasma membrane, only stained by PI).
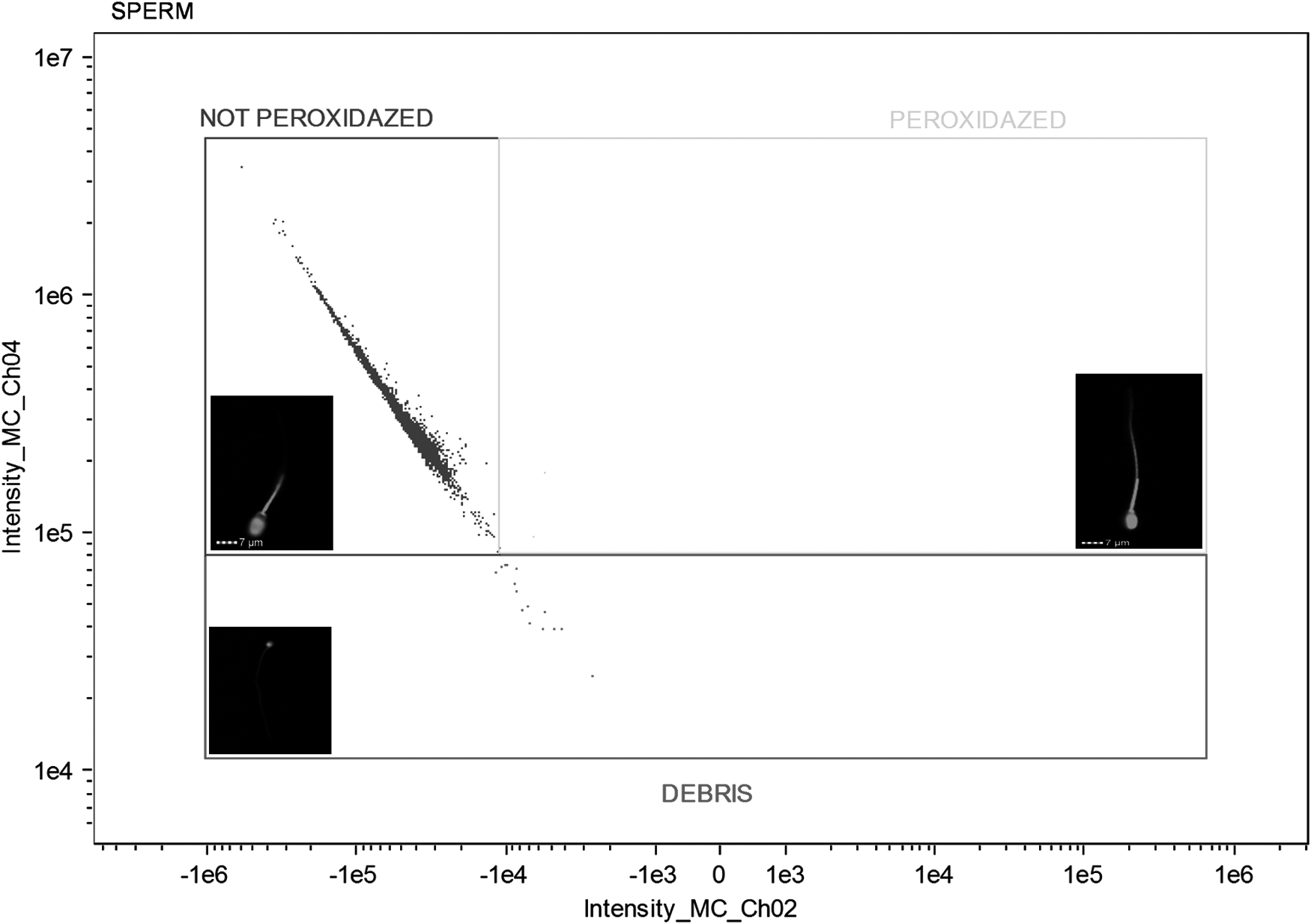
LPO in plasma membrane of ram semen after freezing–thawing in semen extender supplemented with canthaxanthin antioxidant. DEBRIS (extender waste and cellular debris tails, stained); C11- (cells without LPO, stained); C11+ (cells with LPO, stained or double stained). LPO, lipid peroxidation.
Statistical analysis
The results are expressed as the mean ± standard deviation. The data were tested for residual normality and homogeneous variances using the Kolmogorov–Smirnov and Bartlett's tests, respectively. Logarithmic transformation [log(X)] was used for LIN and VCL and square root transformation (√X) was used for TM and WOB to obtain a normal distribution. The other variables expressed as a percentage were transformed by the arcsine method (arcsine √P/100). All data were analyzed by an analysis of variance (ANOVA) and Tukey's test with a significance level of 5% (p < 0.05), which were processed with GraphPad InStat 3.0.
Results
The effects of supplementing the freezing extender with different canthaxanthin concentrations on sperm motility are presented in Table 1. For TM, no difference (p > 0.05) was detected among the control group and the treatments after thawing and 2 hours of incubation. However, a difference was observed (p < 0.05) between the 1 and 25 μM canthaxanthin concentrations after thawing, whereas no difference between these two groups was observed at 2 hours of incubation (p > 0.05).
Different small letters in the same column, when present, denote differences among treatments (p < 0.05) within each time; different capital letters in the same column, when present, denote a difference among the times (p < 0.05) within the same treatment if any exist.
ALH, amplitude of lateral head displacement; BCF, beat-cross frequency; LIN, linearity; PM, progressive motility; SD, standard deviation; STR, straightness; TM, total motility; VAP, average path velocity; VCL, curvilinear velocity; VSL, straight-line velocity; WOB, wobble.
The percentage of WOB was higher (p < 0.05) in the control group compared with the 25 μM canthaxanthin group, whereas ALH was lower (p > 0.05) in the control group than in the 25 μM of canthaxanthin treatment immediately after thawing. However, these differences were not observed at 2 hours of incubation. Regarding the BCF, differences were observed (p < 0.05) between the control and the treatments of 0.1 and 10 μM of canthaxanthin after thawing, whereas no difference was observed in the evaluation at 2 hours.
After the 2-hour incubation period, no differences between groups for the kinetic variables were observed (p > 0.05). However, the VCL and ALH variables increased within the same group (p < 0.05) during the incubation period for the control and 1 μM of canthaxanthin. This was associated with the decrease in LIN and WOB (p < 0.05). BCF, however, increased (p < 0.001) only in the control group, whereas no difference was observed for the canthaxanthin concentrations of 10 and 25 μM. The STR values for all treatments except with 25 μM of canthaxanthin present decreased during the incubation time. For the other kinetic variables, PM, VSL and VAP, no difference was observed (p > 0.05).
Table 2 shows the effects of supplementing the extender with different concentrations of canthaxanthin on sperm variables as evaluated by flow cytometry. The results show no difference between treatments within each time (0 and 2 hours) or between the times for each treatment in the analysis of the iPAM, intracellular ROS production, and LPO of the plasma membrane (p > 0.05).
No differences were observed either among the treatments or among evaluation times (p > 0.05).
PNA−/PI−, cells with intact plasma and acrosomal membranes (FITC-PNA+PI); DCFDA−/PI−, cells with low ROS levels and intact plasma membrane (CM-H2DCFDA+PI); C11-BODIPY-, cells without lipid peroxidation in the plasma membrane (C11-BODIPY591/581).
ROS, reactive oxygen species.
Discussion
Our results demonstrate that addition of canthaxanthin to a Tris-egg yolk extender caused changes in some parameters of sperm kinetics. Although the canthaxanthin 1 μM group presented a 4.9% higher TM than the control group at 0 hours post-thawing, this difference was not statistically significant, possibly due to the high standard deviation for this variable.
Regarding the TM, the 25 μM canthaxanthin group had a lower TM than the 1 μM group, which may indicate a dose-dependent effect of canthaxanthin on sperm motility immediately after thawing as this difference did not persist after 2 hours of incubation. This effect can be explained by the fact that canthaxanthin interacts with lipid membranes, exerting a strong influence on the physical properties of the membrane, leading to destabilization, restrictions to the segmental molecular motion of lipids, and changes in the membrane surface. 21 This lipid membrane–canthaxanthin interaction can also relate to the superiority of the control group compared with the 25 μM canthaxanthin group for variables WOB and ALH immediately after thawing.
Canthaxanthin can be incorporated into lipid membranes in two ways: across the lipid bilayer more or less perpendicular to the membrane surface or horizontally, in parallel to the membrane in the region of hydrophilic heads of phospholipids. 31 With the increases in concentration, an increase in fraction of the horizontal population occurs, which can lead to modification of the dynamic and structural properties of the lipid membrane. 32 This behavior as well as the fact that sperm cells undergo an intense osmotic imbalance at the time of thawing 33 may have influenced the motility of cells exposed to a higher concentration of canthaxanthin (25 μM). This was not observed after 2 hours of incubation at 37°C, suggesting that a reaccommodation of the membrane with the recovery of its functionality occurred.
The behavior was exhibited by the control and the 1 μM canthaxanthin groups after 2 hours of incubation at 37°C in the freezing extender, where increases in VCL, ALH, and BCF associated with the reduced LIN, STR, and WOB were observed, indicating a movement pattern similar to hyperactivation sperm, which is observed in the spermatozoa in the oviduct at the time of fertilization. 34 The results indicate that the higher canthaxanthin concentrations used (10 and 25 μM) were able to maintain the kinetics of cells during the incubation period without allowing spermatozoa to exhibit a change in swimming pattern compatible with hyperactivated motility.
Although it is difficult to determine a swimming pattern for the hyperactivated spermatozoa for all species, this state is characterized by a more vigorous and less progressive movement, a pattern that has already been established in some species such as rats, humans, and bovine and ovine species.35–38 Hyperactivation is associated with the capacitation process and gives the spermatozoa the skill to overcome the mucus of the oviduct and the zona pellucida to fertilize the oocyte.39,40
However, this process should occur in the oviduct near the time of ovulation. 41 Thus, an early hyperactivation can be related to the low fertility rates in vivo of cryopreserved semen. On the other hand, the ram semen with a higher proportion of spermatozoa with linear movement have a greater fertilizing potential by presenting more efficient migration through the female reproductive tract, thereby cell populations with characteristics of hyperactivation (rapid and nonprogressive) are correlated with subfertility. 42
Moreover, Del Olmo et al. affirmed that evaluation of the kinetics of ram semen after incubation for 2 hours at 37°C in a freezing extender could present a correlation with fertility when compared with the post-thawing evaluation and that the pretreatment of semen before its evaluation is an important tool for predicting their fertilizing capacity. 43 This reinforces the beneficial results of 10 and 25 μM of canthaxanthin in the evaluation 2 hours after thawing.
The dose-dependent effect of the canthaxanthin concentration on sperm kinetics observed at 0 hours was not present in the integrity of plasma and acrosomal membranes. It is probable that the disruption caused by the concentration of 25 μM in the kinetic variables (TM, WOB, and ALH) could not cause impairment to the integrity of membranes. Alternatively, we can cite a study where astaxanthin, a carotenoid that like canthaxanthin belongs to the class of xanthophylls, was used to supplement an extender egg yolk based on the concentrations of 2 and 4 μM, which improved the integrity of the plasma membrane and the total and PM of ram sperm after 72 hours of liquid storage at 4°C. 44
It is noteworthy that these results were observed in cooled semen that unlike our experiment, did not suffer intense osmotic shock, as noted in the freezing and thawing processes. In addition, the gametes in this study were submitted to a long period of storage (72 hours), which manifested through elevation in the ROS and malondialdehyde (MDA) levels, which can explain the protective effect of astaxanthin to spermatozoa.
The evaluation of intracellular ROS production showed that the use of canthaxanthin at these concentrations resulted in no difference compared with the control in the number of viable cells with low ROS levels post-thawing in any of the times evaluated (0 and 2 hours). In addition, Kadirvel et al. working with buffalo sperm and Santiani et al. working with ram sperm found that intracellular ROS levels in thawed viable spermatozoa did not differ significantly from fresh semen.45,46 These results demonstrate that under certain conditions, the ROS levels are not affected by cryopreservation; thus, a reduction in these levels was not observed, unlike in the use of an antioxidant therapy.
Although several studies have demonstrated that an increase in LPO in the plasma membrane of frozen and thawed spermatozoa occurs as a result of oxidative stress during the cryopreservation processes in different species of mammals,47–50 our results from evaluating LPO using C11-BODIPY591/581 associated with flow cytometry demonstrated that cryopreservation performed under the conditions established in this experiment did not lead to peroxidation of lipids of the plasma membrane of ram spermatozoa, with 0.8% and 1.8% of sperm presenting LPO in fresh and thawed semen, respectively.
These results corroborate those reported by Guthrie and Welch, which demonstrated very low levels of sperm presenting LPO in fresh (1.3%) and thawed semen (3.4%) of boar, evaluated with the C11-BODIPY591/581 probe, 51 as well as those reported by Peris et al., which showed no change in MDA levels after the cryopreservation of ram semen. 14
In conclusion, supplementing 10 and 25 μM of canthaxanthin into a Tris-egg yolk extender used for ovine sperm cryopreservation can protect the semen after incubation at 37°C for 2 hours from changes caused by the freezing/thawing process, such as hyperactivation. Moreover, no effect of supplementation was observed on the iPAM, intracellular ROS production, and LPO of the plasma membrane during the cryopreservation process.
Footnotes
Acknowledgments
The authors are grateful to the following Brazilian fostering agencies: Fundação de Amparo à Ciência e Tecnologia do Estado de Pernambuco (FACEPE) for awarding a scholarship to the doctoral student and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for funding.
Author Disclosure Statement
No conflicting financial interests exist.