
Abstract
In the present study, we examined various freezing protocols, effects of controlled seeding, and changes in cooling rate and determined the endpoint (temperature at which sample could be plugged into liquid nitrogen (LN) without visible effect on survival rate after thawing) to reveal the relative importance of each different stage of cooling on freezing success during cryobanking of carp sperm. Sperm samples from different individual carp males were frozen in 0.5 mL straws by conventional freezing. Cooling rates were determined by monitoring the sample's internal temperature. We compared four freezing protocols, which involved placing sperm samples at various levels (1, 3, 6, and 9 cm) above the LN surface (corresponding to −190°C, −150°C, −110°C, and −70°C, respectively) for 20 minutes followed by transferring the samples into LN. Freezing at 3 cm above the LN surface resulted in the highest motility (33% ± 8%) and velocity (118 ± 9 μm/s) of spermatozoa after thawing and diluting in swimming medium. We determined that −90°C is an optimal temperature at which immersing the samples in LN does not affect sperm motility after thawing and shorten the process of freezing for around three times. Motility of spermatozoa cryopreserved with or without a seeding procedure was not significantly different after thawing. Therefore, we hypothesize that supercooling the sample during the conventional freezing procedure is not the main damaging factor during carp spermatozoa cryopreservation. However, the cooling rate itself is important, because it determines the ability of the sperm to dehydrate and survive cryopreservation.
Introduction
S
This study is focused on determination of the relative importance of different stages of the cooling process on sperm freezing success during cryopreservation of carp sperm. We selected one cryoextender among several published earlier (with a reliably high post-thaw survival rate) to conduct our experiments: (1) study the effect of different cooling rates, (2) detect the optimal endpoint (temperature at which sample could be plugged into LN without visible effect on survival rate after thawing), and (3) study the effect of controlled seeding and supercooling on survival of common carp sperm after thawing.
Materials and Methods
Ethical approval
The study was approved by the Ethics Committee for the Protection of Animals in Research of the University of South Bohemia in Ceske Budejovice, Research Institute of Fish Culture and Hydrobiology, Vodnany, based on the EU-harmonized Animal Welfare Act of the Czech Republic.
Gamete collection
Fish were maintained in a pond and transferred after harvesting into individual 4 m3 hatchery tanks with 0.2 Ls−1 water flow rate. Temperature and oxygen ranged from 16°C to 18°C and 6–7 mg L−1, respectively. Before handling, fish were anesthetized in a 1:1000 aqueous solution of 2-phenoxyethanol. Males were injected with carp pituitary extract at 1 mg kg−1 24 hours before striping according to fisherman practice in the Czech Republic. 10 Sperm was obtained by abdominal massage and was collected directly into 10 mL plastic syringes. The process was carried out very carefully to avoid contamination with urine, mucus, feces, or water. Sperm samples from different males were not pooled and were stored under aerobic conditions at 4°C not longer than 2 hours.
Evaluation of sperm motility parameters
Hatchery water was used as a hypotonic activating medium (AM). In the case of freshly collected milt, 50 μL of AM was placed on a glass slide in a microscope stage; sperm was added by using the tip of a dissecting needle; and the sperm suspension was thoroughly mixed for 2 seconds. The same activation procedure was used for cryopreserved sperm immediately after thawing. After dilution and mixing, motility was video-recorded for 1 minute postactivation by using a CCD video camera (Sony, SSCDC50AP; Sony) that was mounted on a dark-field microscope (Olympus BX50, × 200) and illuminated with a strobe light by using an ExposureScope set to a flash frequency of 50 Hz, synchronized to the camera frame frequency. Several video records per male (2–4) were collected by using a video recorder (Sony SVHS, SVO-9500 MDP) and were subsequently analyzed to estimate the average velocity (μm s−1) and percentage of sperm motility (%). Both parameters were evaluated at 10 seconds postactivation by analyzing five successive frames using a micro-image analyzer (Olympus Micro-Image 4.0.1. for Windows). Ten to fifty spermatozoa were evaluated from each frame. Spermatozoa with a velocity lower than 10 μm/s were considered immobile and were excluded from further analysis. Velocity was measured only for motile spermatozoa in total 30–100 per male.
Experimental design
Before the experiments, four different cryoextenders were selected, based on following criteria: (1) all of them have been reported as successfully applied for carp sperm cryopreservation2,4,5; (2) they consist of different cryoprotectants, different saline composition, and egg yolk presence; (3) they were tested in our laboratory earlier and provided relatively stable results. The composition of cryoextenders used in the study is as follows:
(1) 59 mM NaCl, 0.68 mM KCl, 0.68 mM CaCl2, 2.1 mM MgSO4, 27 mM NaHCO3, 3.4 mM sucrose, 69 mM D-mannitol, 118 mM Tris-HCl pH 8.1, 16% ethylene glycol, and 10% egg yolk (modified after Kopeika, 1986
5
); (2) the same as number 1, but without egg yolk; (3) 62 mM NaCl, 134 mM KCl, 1.5 mM CaCl2, 0.39 mM MgCl2, 2.4 mM NaHCO3, and 11% dimethyl-sulfoxide (modified after Kurokura, 1984
4
); and (4) 350 mM glucose, 30 mM Tris-HCl pH 8.0, and 20% methanol (Horvath, 2007
2
;).
Sperm samples from six individual males were frozen in 0.5 mL straws at a distance of 3 cm above the liquid nitrogen (LN) level for 20 minutes and then immersed in LN. Percentages of sperm motility were determined after thawing. First, cryoextender (with highest survival rate after thawing; see results section) was later used for three different sets of experiments:
(I) Comparison of sperm motility parameters of carp sperm cryopreserved under different cooling protocols.
Sperm from 13 individual males (1 straw per each) were frozen at distances 1, 3, 6, and 9 cm above LN with cryoextender 1. Sperm motility, velocity, and duration were measured after thawing. Thermograms of sperm cooling at different levels above LN are presented in Figure 1.
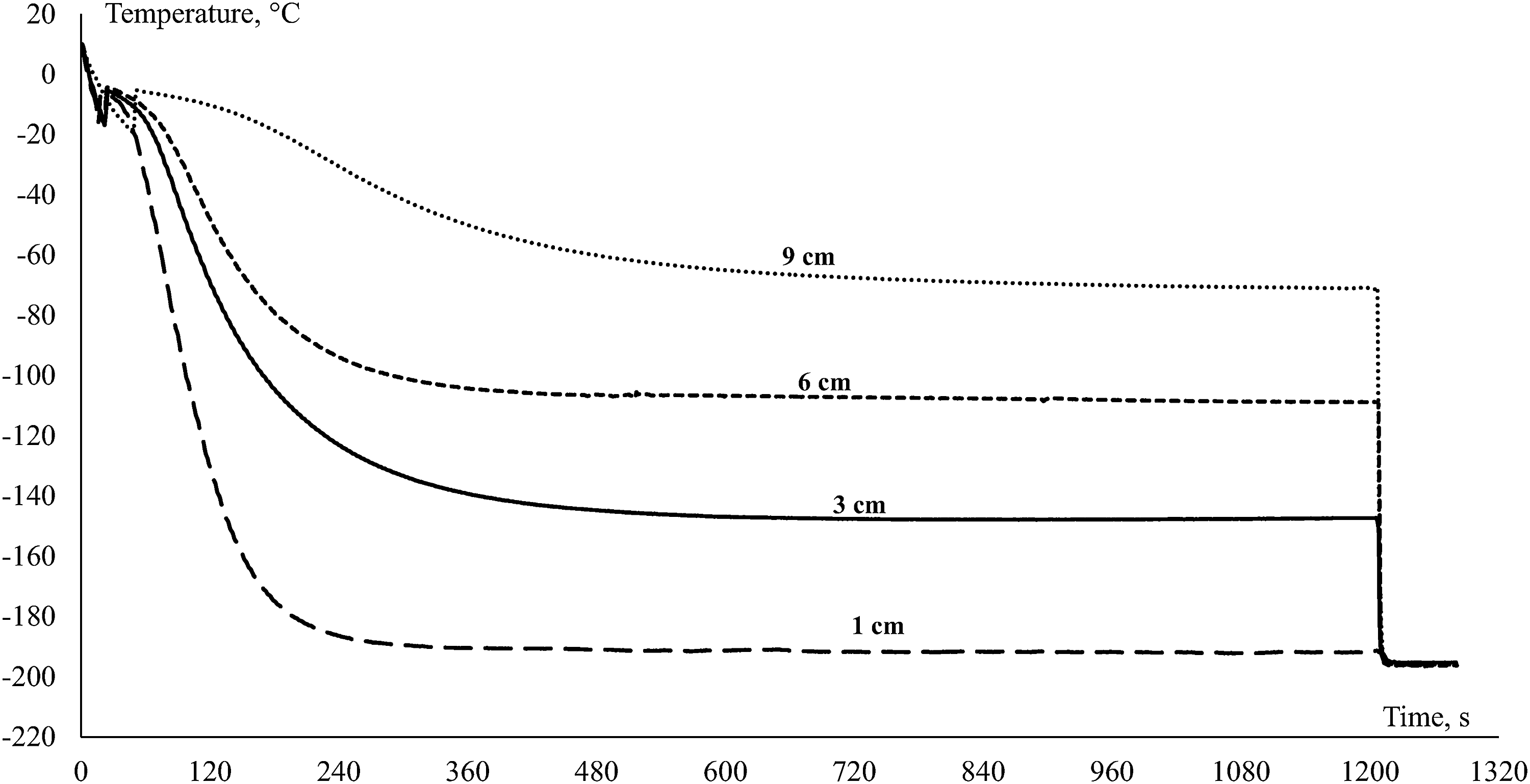
Representative thermograms of carp sperm freezing at different levels (1, 3, 6, and 9 cm) above the liquid nitrogen level.
(II) Determination of optimal endpoint.
Sperm samples in 0.5 mL straws with cryoextender 1 were set 3 cm above the LN surface and were immersed into LN at different time points (according to internal temperature inside sample). Sperm motility parameters were determined after thawing sperm samples from seven males and compared with control conditions (20 minutes of incubation before plugging in LN).
(III) The study of seeding procedure efficiency.
To investigate possibility to improve the success of carp sperm cryopreservation, we applied a seeding procedure to the protocol with the highest results of sperm survival after freeze/thawing (0.5 mL straw at 3 cm above LN with cryoextender 1; see results section). Sperm motility was measured after thawing of samples from seven males, and the results were compared with cryopreservation under the same conditions but without seeding (control condition). Seeding was applied at −6°C before crystallization beginning (temperature to which samples reached after initiation of crystallization under control condition, Fig. 3) and then samples were cooled similar to the control condition. Two different methods of seeding were used to induce crystal formation and to protect samples from overcooling:
(a) Dipping the raft with sperm samples into the LN for 1 second.
(b) Touching the middle of the straw containing the samples with forceps (approximately for 1 second) previously cooled in LN. 18
Freezing and thawing procedures
Sperm from males with motility values of at least 90% were individually retained for freezing experiments according to methodology. 10 Before freezing, sperm was diluted (1:1) in different extenders according to the experimental design. Extender was added to the sample drop by drop during constant gentle mixing to avoid an osmotic shock of spermatozoa by cryoprotectant. After dilution, sperm samples were loaded into 0.5 mL straws (CRYO-VET) and stored for 10 minutes on ice. The straws were placed horizontally on Styrofoam rafts of various heights (1, 3, 6, and 9 cm, according to the experimental design) floating on the surface of LN. After a certain period (according to the experimental design), the straws were immersed in LN. After 2 days of storage in LN, samples were thawed in a 40°C water bath for 6 seconds. The temperature inside/outside of the straws during different freezing procedures (including seeding) was recorded with a copper/constantan thermocouple (0.2 mm in diameter) connected to a computer via a calibrated Data Logger (Omega), and temperature was recorded once per 2 seconds. The thermocouple inside of the sample was located straight in the middle of the straw. Each measurement was repeated five times. Cooling rates were counted in linear part of thermograms and presented as °C min−1.
Statistical analysis
Values of sperm motility in the tables and text are presented as mean ± 0.95 confidence interval of all males (n = 6–13; related on experimental conditions). Values of cooling rates are presented as mean ± 0.95 confidence interval of five measurements. Data were analyzed by STATISTICA software (StatSoft, Inc.) using ANOVA and Post Hoc Tukey HSD test (p < 0.05).
Results
Selection of cryoextender
Four different cryoextenders were tested in our study. Post-thaw sperm motility is presented as the percentage of all sperm in Table 1. The highest percentage of post-thaw sperm motility was observed in the samples that were cryopreserved in cryoextender 1 (p < 0.05, n = 6). Sperm cryopreserved with medium 2 or 3 showed lower percentages of movement and were not significantly different between each other. A correct measurement of sperm motility after cryopreservation with cryoextender 4 was not possible, because part of the spermatozoa population agglutinated. Therefore, cryoextender 1 was the most appropriate among the cryoextenders tested and was used for the experiments.
Values with different numbers of asterisks are significantly different (n = 6; p < 0.05).
n/a, none available.
Cryopreservation of carp sperm under different cooling protocols
Thermograms of carp sperm freezing at different levels (1, 3, 6, and 9 cm) above the LN level using the 0.5 mL straw can be seen in Figure 1. Temperature of freezing and cooling rate before/after crystallization point at corresponding level above LN is presented in Table 2. The rate of cooling before crystallization point at 9 cm above LN was much lower compared with 1, 3, and 6 cm. Sperm motility values after freezing in straws placed at different levels above the LN and using cryoextender 1 are presented in Table 3. The highest values of sperm motility parameters, such as motility percentage, velocity, and period of motility, were observed in fresh samples. Cryopreservation of sperm samples placed in a straw 1 cm above the LN resulted in the complete absence of motility. Samples cryopreserved at 3 cm above the LN had significantly higher post-thaw motility and velocity (p < 0.05, n = 13). However, the period of sperm motility was not significantly different among the samples frozen at 3, 6, and 9 cm above the LN.
Within a column, means without a common superscript differ (n = 5; p < 0.05).
Within a column, means without a common superscript differ (n = 13; p < 0.05).
Determination of optimal endpoint
Figure 2 presents the thermograms of carp sperm incubated at 3 cm above LN for different time periods before being immersed in LN (vertical arrows). Samples were immersed in LN at −6°C (after crystallization point), −90°C, and −145°C to determine the optimal endpoint of slow freezing. Sperm motility for the samples immersed in LN before or immediately after crystallization point (−16°C) was 0%. In the case of sperm samples, which were immersed in LN after cooling to −90°C or lower temperatures, motility after freeze/thaw was not significantly affected (29% ± 9% and 34% ± 8% if compared with control 33% ± 8%; p = 0.802 and p = 0.937, n = 7).
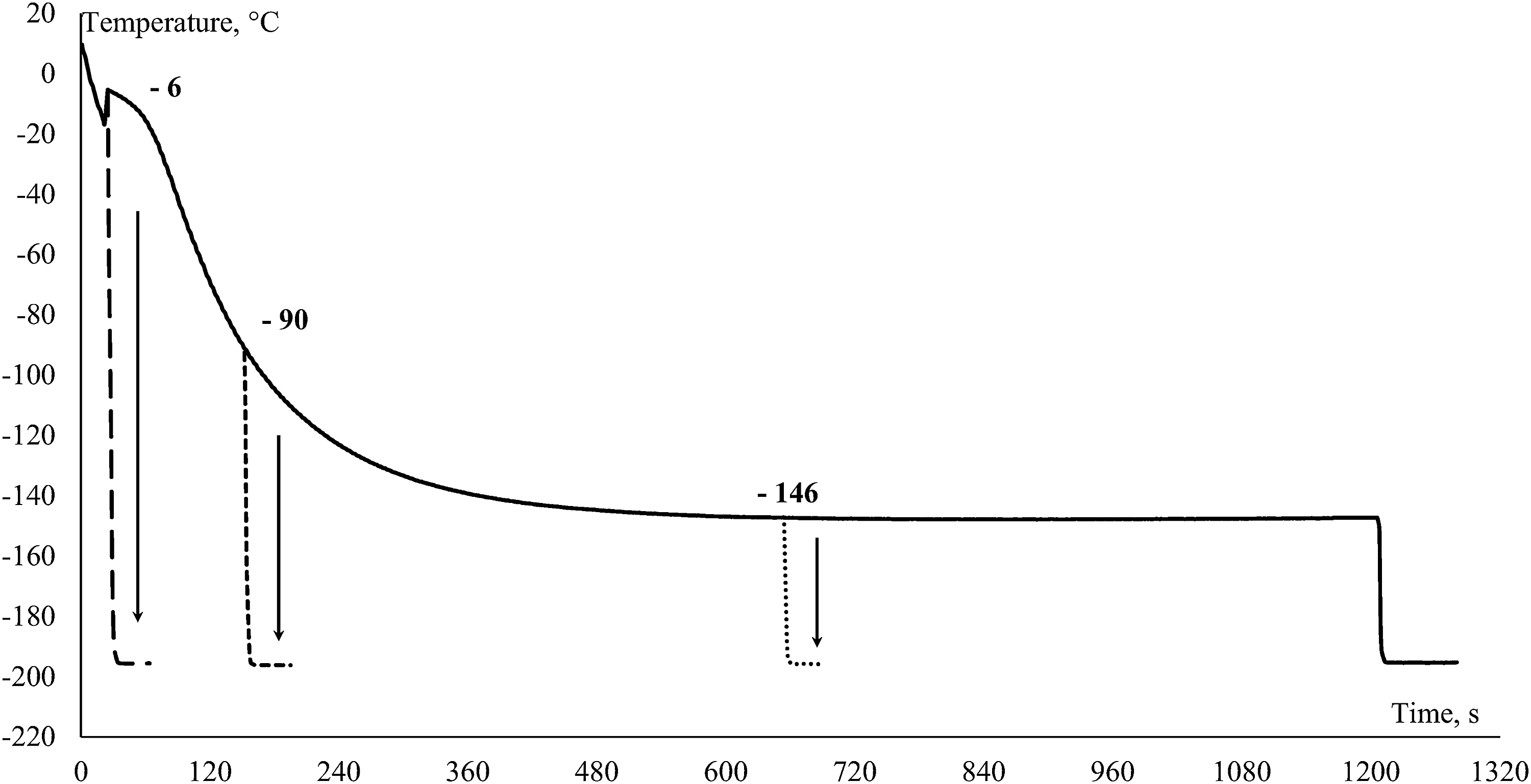
Representative thermograms of carp sperm incubated 3 cm above liquid nitrogen for different time periods before being immersed in liquid nitrogen (vertical arrows).
Seeding of carp sperm
Thermograms of sperm freezing using two seeding conditions are presented in Figure 3. Using the method “a” of seeding (where a raft was plunged into the LN for 1 second) led to a change in the cooling rate of samples from −59°C ± 4°C to −115°C ± 10°C min−1 and resulted in 0% post-thaw motility. In contrast, the method “b” of seeding (cooled forceps) did not lead to changes in the cooling rate (−58°C ± 6°C min−1), except the absence of supercooling (Fig. 3). Using these conditions, we did not observe any increase in sperm motility after thawing (35% ± 10%) compared with the control condition (without seeding 33% ± 8%; p = 0.701, n = 7).
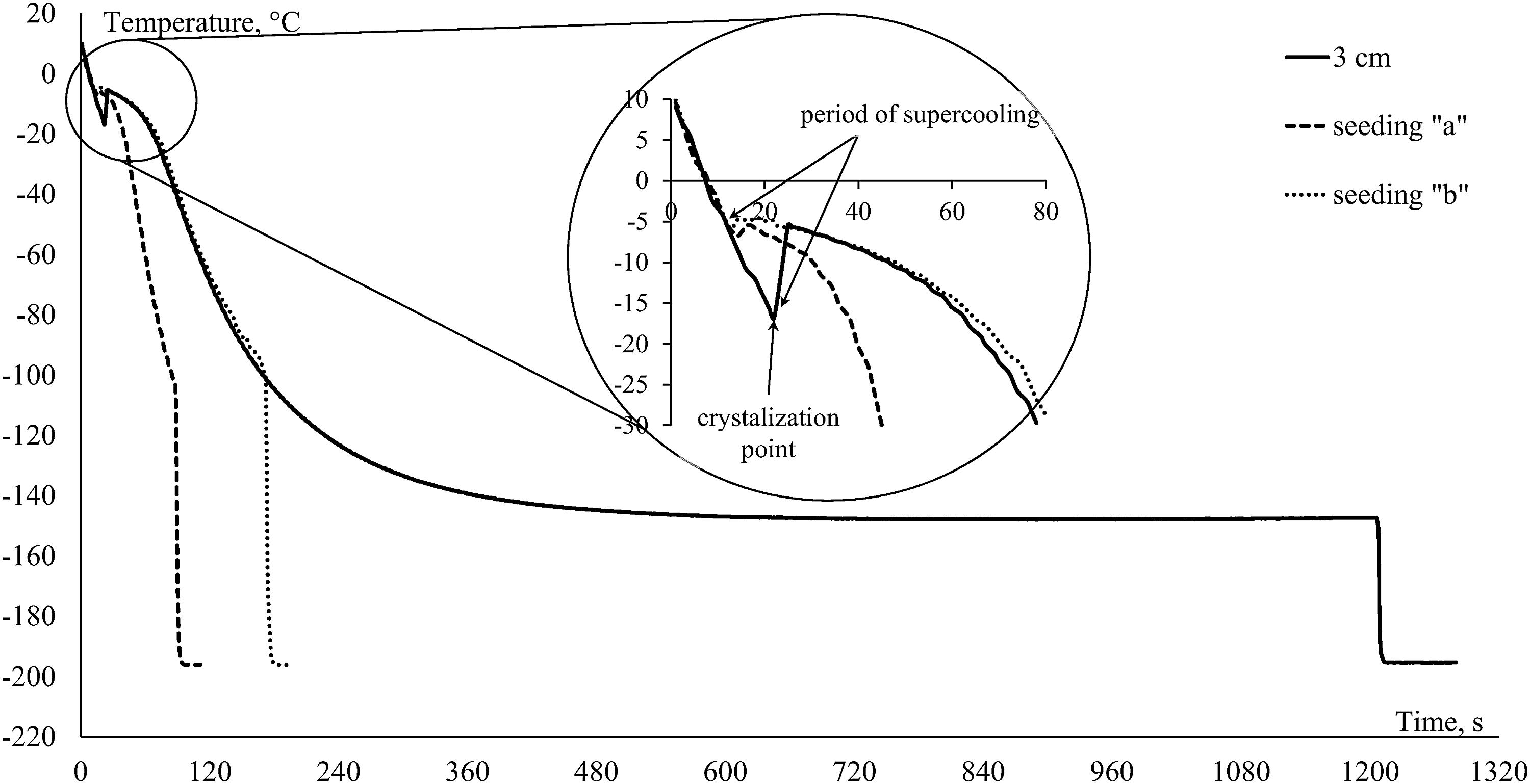
Representative thermograms of sperm in the process of freezing by using two conditions for seeding compared with the control condition (3 cm above liquid nitrogen). “a”—dipping the raft containing sperm samples into the liquid nitrogen for 1 second; “b”—touching the middle of the straw containing the samples for 1 second with forceps previously cooled in liquid nitrogen.
Discussion
High variability among carp subpopulations all over the world results in differences in the physiology of these fish, in general, and in reproduction physiology, in particular. 19 This phenomenon could result in differences in sperm motility, fertilization success, or cryoresistance, which stimulate further studies on development of new cryopreservation methods. In the present study, we examined four cryoextenders previously described. Our results in carp sperm post-thaw survival rate are different from those reported earlier (the same cryoextenders and similar freezing conditions were used)2,4,8,9 and indicate low repeatability of sperm cryopreservation methods for different populations of common carp. However, in our study, the cryoextender proposed by Kopeika 5 resulted in the highest post-thaw motility and was retained for subsequent experiments. To better understand the principles governing fish sperm cryopreservation, only physical factors (such as cooling rate) varied during the experiment, and the composition of the medium 5 was constant.
Different rates of cooling and sperm damage
Our results of post-thaw motility demonstrate a significant difference among the samples frozen at different levels above the LN (different protocols of cooling). Thus, an optimal freezing rate was −81°C min−1 before the crystallization point and −59°C min−1 after it; this rate corresponds to the temperature of samples incubated at −150°C (3 cm above the LN, Fig. 1). Using this method resulted in the highest velocity values and percentage of motility after thawing in comparison with the other protocol (1, 6, or 9 cm above LN); however, the values were still lower than those obtained with fresh sperm. This can be a result of osmotic changes during crystal formation, which would lead to nonlethal (decrease in velocity) versus lethal (decrease in motility) damage to spermatozoa during cryopreservation. 17 Thus, different cooling rates could influence the speed of ice crystal formation and the structure of the corresponding crystals, which, in turn, induces different levels of damage and affects post-thaw motility.
The influence of the cooling rate on the success of cryopreservation is a well-known fact and has been described by several authors.3,6,15 Basically, when the rate of cooling is slower than optimal, the cell damage could be associated with a longer period of time when cells are exposed to extremely high osmolality. 20 Thus, the slower the cooling rate is applied, the less cells could survive the osmotic stress to which they are exposed during freezing (in our experiments: cryopreservation at 6 and 9 cm above LN). At higher cooling rates, the damaging factor of cell exposure to high osmolality is reduced, but in this case cells have less time for water and cryoprotectant transport across the membrane. However, fast cooling rates (faster than optimal) may lead to intracellular crystal formation and as a result, lethal damage of the cells. It corresponds to our results of freezing at 1 cm above LN, when 0% of the cell population survives cryopreservation.
Optimal endpoint for slow cooling rates
In this study, we also determined an optimal endpoint (temperature less than −90°C) at which samples could be transferred into LN, and subsequent post-thaw motility would be not affected. Our results correspond to those from previous studies5,18 in which the samples were moved to LN at −80°C. However, Viveiros et al. 15 demonstrated that the sample could be transferred to LN after some period of exposure at an even higher subzero temperature (around −40°C). In comparison to those results, our experiments were conducted at faster cooling rates, suggesting that cooling rate and optimal endpoint are dependent parameters. 15 This result makes our finding useful in practice, when a large number of sperm samples need to be cryopreserved, as the time needed to freeze one single sample is reduced to approximately 3–4 minutes in comparison to the commonly used 10–20 minutes. When transferring samples into LN before or immediately after the crystallization point, the resulting motility was 0%. In this case, the cooling rate was likely very high, which can cause nonreversible damage to the spermatozoa during freezing, so the situation is similar to rapid cooling at 1 cm above LN. Therefore, we conclude that the cooling rate determining ice crystal formation from the crystallization point down to −90°C is an essential parameter that controls successful cryopreservation of fish sperm.
Effect of seeding and supercooling on sperm samples during cryopreservation
Finally, we define two methods of controlled seeding. When the raft with the samples was dipped into the LN for 1 second, the cooling rate increased. This increase most likely happened due to increased LN evaporation after contact with the raft and, consequently, this led to a situation that was similar to freezing at 1 cm above the LN, which caused 0% motility after thawing (even when supercooling was absent). The second method of seeding with cooled forceps also prevented samples from supercooling, whereas the cooling rate and sperm motility ranking were not affected.
The Styrofoam rafts used to freeze sperm in our experiment provide free heat exchange between the sample and the surroundings. The sample releases latent heat of ice formation into the surroundings immediately after ice crystallization is initiated, and the surroundings “adapt” to the sample temperature and are not forcing the cooling rate. This is demonstrated in Figure 3 (freezing curve after seeding “b” and 3 cm) when the two curves overlap each other. Therefore, we suppose that supercooling the sample during conventional freezing does not cause damage under such experimental conditions. However, damage could appear in a programmable freezer (which controls temperature inside of samples) without the use of seeding. In that case, after supercooling and spontaneous crystallization, the program will try to compensate for latent heat released from the sample by additional cooling delivered to the chamber. Thus, when the heat is released, the chamber will reach quite low temperatures. Due to the crystallization, much damage could occur, as water will have no time to exit the cell and will trigger immediate internal ice crystallization.15,20 However, Morris et al. 11 demonstrated the absence of intracellular ice formation during rapid cooling of spermatozoa and suggested that damage to spermatozoa is a result of an osmotic imbalance encountered during thawing. These mechanisms of damage most likely explain the extremely low motility in our experiments, when seeding by dipping a raft into LN was applied (seeding “a”).
Another possible damaging factor could be caused by the very small ice crystals that are formed at low temperatures during rapid cooling. Smaller crystals could be the least damaging. However, on thawing, small crystals melt at a temperature that is still very low. At this temperature, the melted water will still be able to crystallize, producing considerably larger crystals of ice by a re-crystallization process and, therefore, leading to greater damage. 21
Conclusions
We demonstrate that the cooling rate after the ice crystallization point and before −90°C has been reached is the most critical factor for successful cryopreservation of fish sperm, whereas supercooling during conventional freezing does not cause damage to the carp sperm. This result explains why the determination of cooling rate represents a crucial step in any freezing protocol development for fish sperm. We determined the endpoint for carp sperm cryopreservation during conventional freezing, which, in turn, results in 3× shortening of time, which is required for freezing of one sample. We assume that the present set of data could be used for the development of optimal cooling protocol for cryopreservation methods for gene banking of other aquatic species. The combination of using programmable freezers and seeding procedures should be addressed in future experiments.
Footnotes
Acknowledgments
The study was financially supported by the Ministry of Education, Youth and Sports of the Czech Republic—projects “CENAKVA” (No. CZ.1.05/2.1.00/01.0024), “CENAKVA II” (No. LO1205 under the NPU I program), COST (No. LD14119), and COST Office (Food and Agriculture COST Action FA1205: AQUAGAMETE), by the Grant Agency of the University of South Bohemia in Ceske Budejovice (No. 125/2016/Z), and by the Czech Science Foundation (GACR 16-02407Y). The authors greatly appreciate the useful comments of Catherine Labbe, Barry Fuller and valuable assistance provided by Jacky Cosson during article preparation. The abstract of the article and part of the results were used in the final thesis of Denisa Sochorová at the Faculty of Fisheries and Protection of Waters.
Author Disclosure Statement
No conflicting financial interests exist.