
Abstract
Cryopreservation (CP) is a critical component in enabling on-demand access to biological material (macromolecules, cells, and tissues), yet CP has evolved little over the last several decades. Today's CP processes often yield a suboptimal “product,” which has slowed progression in areas such as cell therapy and stem cell research. Recent discoveries focusing on molecular control and buffering of cell stress responses to CP as well as the development of new devices that improve sample freezing and thawing are now providing a path to improve sample quality. Numerous reports now identify the problems associated with CP-induced delayed-onset cell death (CIDOCD), with a focus on the mitigation of cell stress responses through freeze media formulation to optimize survival during “cold chain” sample management. Importantly, these new approaches that manage cell stress response are now providing a basis for advancements in cell therapy development. Herein, we provide an overview of the molecular stress responses of cells to CP, including the impact of hypothermic and cell death continuums and the strategies for improving preservation outcome.
Introduction
B
While process optimization has advanced over the past decade, significant limits with current cryopreservation (CP) practices remain. They include (1) continued post-thaw cell death, (2) prolonged loss of function, (3) use of animal-based products in the storage media, (4) activation of molecular stress response pathways in “surviving” cells, and (5) downstream effects on epigenetic control of gene expression and protein levels.3–9 CP-induced, inheritable epigenetic events may change our understanding of the impact of low temperature on cell function well beyond that of the physical/chemical changes (i.e., osmolarity, membrane disruption), which constitute our historic understanding of the post-thaw survival by bringing into play the complex molecular and cell signaling pathways identified over the past two decades.
The preservation of biologics utilizes low temperatures either in the form of hypothermic storage or CP. Hypothermic storage is defined as the use of nonfreezing (>37°C–0°C) temperatures to maintain biologics in a reduced metabolic state. A temperature range between 4°C and 10°C is most commonly utilized. In contrast, CP extends the hypothermic window to ultralow temperatures (less than −80°C to −196°C). Long-term storage below −140°C is considered ideal as the nominal glass transition temperature (Tg) of water is achieved. The latter approach has demonstrated qualified but successful outcomes necessary to launch the fields of stem cell research, regenerative medicine, and tissue engineering. However, the clinical translation of these fields is placing new demands on current protocols as limitations of current CP methodologies are revealed. 10
To respond to the new demands for post-thaw cellular integrity, a molecular-based focus is emerging. The activation of cell stress pathways post-thaw (i.e., gene-regulated cell death) necessitates attention to molecular events that impact both CP and hypothermic storage. This article provides an overview of CP focusing on a molecular perspective while discussing strategies useful in mitigating the magnitude of post-thaw delayed-onset cell death.
Evolution of CP Strategies
Modern efforts toward the freeze preservation of mammalian cells (at or below −80°C) can be traced to the discovery that glycerol could serve as a preservative for avian spermatozoa 11 and human red blood cells. 12 The postulated primary role of glycerol, as a cryoprotective agent (CPA), was to prevent intracellular ice formation. These reports were significant to the evolving field of CP by identifying (1) the need for a CPA, (2) the process of exposing cells to penetrating CPA, (3) the protocol for freezing and thawing, and (4) the launch of a deliberative, scientific approach to cell preservation. In 1959, 13 the first use of dimethyl sulfoxide as a CPA was reported with the advantage of both enhanced permeability when contrasted with glycerol and cell survival for many cell types. Advances that followed over the next ∼40 years focused on changes in carrier media containing diverse CPAs along with postulated mechanisms of cryoinjury.1,14 In 1972, Mazur et al. 15 put forth the “Two-factor Hypothesis” that proposed a functional dependency between cooling rates and survival. Suboptimal slow cooling rates were predicted to be toxic due to solution effects caused by the long-term exposure to hyperosmotic conditions following freeze concentration of solutes.15,16 Higher (nonoptimal) cooling rates would result in the formation of lethal intracellular ice formation. Higher suboptimal cooling rates were proposed to cause incomplete cell dehydration thereby increasing the likelihood of intracellular ice formation.15,17,18 What evolved from these studies was the generalization that successful CP was primarily dependent on biophysical parameters that most mammalian cells would respond similarly to a standardized (optimal) preservation protocol. The adoption of a process that involved preincubation of cells with a CPA at ∼4°C followed by cooling at a rate of −1°C per minute to nonstandard subfreezing temperatures (−40°C to −80°C) and quenching/storage in liquid nitrogen provided the “optimal” CP protocol. This approach, however, did not take into account the activation of the yet to be discovered molecular stress response of the cell to CP as a result of the process and “hypothermic continuum.” 1 Rapid thawing was also shown to improve outcomes by limiting the time cell systems were exposed to stressing temperatures and solute levels following the storage stage. 19 While convenient, this protocol was not optimal and, in effect, treated the cell as a biologically passive participant in the process. In effect, these studies established the biophysical foundation on which CP research has rested for nearly half a century.
Hypothermic Continuum
CP research and method development rarely addresses hypothermia as either a significant element of the cold chain preservation process or as a potential zone of low-temperature damage. As described, CP typically focuses on the biophysical events related to the water-phase change and osmotic flux during the controlled rate cooling (CRC) interval with the net outcome of creating two artificially distinct specialized fields of thermal biology. Information emergent from hypothermia research rarely finds translation to CP. This dichotomy finds its roots in a lack of recognition that successful CP includes the transitioning of biologics that avoid intracellular freezing through a deepening state of hypothermia through the glass transition temperature (Tg, nominally −135°C). 1 That is, the intracellular environment experiences ultraprofound hypothermia but not freezing through the freeze–thaw round trip. The depth of hypothermia experienced by a cell can be defined as follows: mild = 32°C to 35°C, moderate = 27°C to 32°C, deep or profound = 10°C to 27°C, and ultraprofound = <10°C). 20
The list of “stress factors” experienced by a cell during intervals of progressively deepening hypothermia is extensive (Table 1). During hypothermic exposure, both a decrease in thermal kinetic energy and ATP availability result in the uncoupling of biochemical reactions within the cell5,21,22 and structural changes in the membrane (transition from liquid crystalline- to solid gel-like state). Changes in membrane-mediated transport that follow 23 yield pronounced ionic imbalances within the cell (i.e., increases in intracellular sodium and calcium concentrations coupled with decreased potassium and intracellular acidosis approaching a pH of 4).5,24–26 Other simultaneous intracellular events include the generation of free radicals, disruption of cytoskeleton, leakage of lysosomal hydrolases, and disruption of the mitochondrial transmembrane potential leading to apoptotic activation.3,5,23,27–29 Clearly, the importance of managing these stressors will be of paramount importance in improved biopreservation.
CP process
CP launches the “hypothermic continuum” in a period of active extracellular ice growth until an intracellular glassy state (vitrified) is reached. 1 CPA exposure relies on a diversity of membrane-permeable, penetrating, and/or nonpenetrating agents dissolved in a carrier media. 30 Cells are incubated in the cryoprotective cocktail (∼10–30 minutes at 4°C) followed by cooling at a nominal (optimal) cooling rate (a range from −1°C to −10°C/min is common for many mammalian cells). Ice nucleation or seeding is best accomplished at temperatures close to the equilibrium melting point of the cryoprotective medium (−2°C to −6°C) to provide for controlled growth of extracellular ice, which is necessary for water efflux from the cell so that supercooling followed by flash freezing of the intracellular environment is avoided. 31 Extracellular ice formation continues as temperature drops limiting the availability of freezable water, yielding freeze concentration of solutes that increases intracellular viscosity. Too rapid a cooling rate yields inadequate cellular dehydration and an increased probability of lethal intracellular ice formation. 18 Nonoptimal preservation protocols support increased cell rupture and necrosis over the first few hours post-thaw.32–34 Prolonged exposure to multimolar levels of CPAs occurs if cooling rates are too slow resulting in cell toxicity (solution effects).15,35 “Solution effect” toxicity has been linked with and is potentially responsible for delayed necrosis (peaking 6–12 hours post-thaw) and apoptosis (12–36 hours post-thaw).33–35
Two methods of CRC are common. One method relies on devices that provide for the controlled injection of liquid nitrogen to the sample to achieve active (programmed) CRC. These devices monitor a representative sample vial, straw, or bag and follow a pre-established program to achieve a desired cooling profile. Profiles are typically set to maintain a standard rate of cooling (e.g., −1°C/min) over a prescribed temperature range and include a seeding event through a thermal shock administered by a surge in cryogen to flatten the temperature rebound resulting from the latent heat of fusion of ice formation. A second passive method utilizes insulated containers (foam or alcohol baths) placed in a −80°C freezer. Passive CRC devices contain the sample surrounded by, but isolated from, an alcohol bath or a thermal-insulation material. When placed in a freezer (−80°C), a curvilinear rate (e.g., approximately −1°C/min) is achieved in the samples. In the passive CRC process, seeding is often accomplished via mechanical agitation to create a nucleation event at a prescribed time during the cooling period. After the nucleation point, sample cooling continues at the controlled rate until a predetermined temperature is achieved (−40°C to −80°C). Once this target temperature is reached, samples may be transferred to an ultralow-temperature environment, such as liquid nitrogen immersion, liquid nitrogen vapor phase, or mechanical storage of less than −135°C. CP storage at −80°C does not provide the same level of long-term stability of viable cells because storage is at temperatures above glass transition temperature (Tg) for water. 36 The importance of this threshold is related to loss of molecular motion (translational and rotational) and the absence of low-level metabolism below Tg and the theoretical loss of all biochemical activity.36,37 The transition through Tg also signifies an end to the hypothermic continuum as cells enter a vitrified (glassy) state surrounded by a highly viscous liquid to glass to extracellular ice boundary. When vitrified, cells are said to be in a state of “suspended animation.” Sample recovery from storage requires rapid thawing typically accomplished by placing the sample container in a 37°C–40°C stirred water bath or in a dry thawing system (SmartThaw, ThawStar, or other device) until most of the ice melts. Rapid thawing limits the time biologics spend in the damaging poststorage hypothermic conditions (between Tg and 0°C, nominally). Once the ice has dispersed, elution of the cryoprotectant cocktail with cell culture media in a single-step or a stepwise (for high CPA concentrations) dilution process is used. Stepwise elution minimizes the volume excursions of the cell, thereby preventing mechanical damage to the cell membrane and rupture.
CP-induced delayed-onset cell death
The success of many CP protocols is assessed immediately (1–2 hours.) post-thaw by dye exclusion assays. These assays fail to provide accurate representation of CP outcome as stress-dependent, cellular damage is manifested hours later. When examined 24–48 hours later, a significant cell loss (up to 70%) is observed due to CP-induced delayed-onset cell death (CIDOCD), an integrated, complex set of cell signaling pathways32,38 (Fig. 1). Dye exclusion assays provide information on structural integrity, which is often interpreted incorrectly as viability. For example, with stem cells, CIDOCD can account for not only a significant loss (>50%) of the total population39–41 but also result in compromised functionality, dysregulated differentiation, alterations in gene expression, and epigenetic changes (i.e., DNA methylation) following a “successful” CP event.6–9,34 As evidence emerged on cell death complexity following the discovery of post-thaw gene-regulated cell death (apoptosis and secondary necrosis), it was recognized that ultimate protocol optimization would require integrative studies that included the biophysical component of cell response along with an understanding of cell signaling cascades driven by a cell's molecular biology.2,3,38 Today, there are four categorical modes of cell death associated with CP that manifest post-thaw, including freeze rupture, necrosis, apoptosis, and necroptosis (sometimes referred to as 2° necrosis) (Fig. 2). Each of these events has been shown to play a critical role in cell survival following CP.32,33 Freeze rupture and necrosis are the most commonly (historically) recognized forms of cell death associated with CP.1,3,10,13,15 Apoptotic cell death was identified in the late 1990s/early 2000s to play a critical role in CP outcome being responsible for CIDOCD.32,33 More recently, necroptosis (a recently described necrotic-like form of programmed cell death) has been implicated,42–47 yet its role in CP failure requires further study. While each of these represents individual paths of cell death, studies have shown that there is a substantial degree of pathway cross talk following CP, further complicating the picture.32,34,38

Timing and Progression of Cell Death Following Cryopreservation. Representation of the progression of the temporal sample survival status during recovery from cryopreservation. Cell viability is typically seen as elevated immediately post-thaw and then progressively decreases during the initial 24 to 48 hours of recovery as apoptotic and necrotic events manifest yielding a “true” survival, which is much lower than initially observed. (Reproduced from Baust et al. 2 )
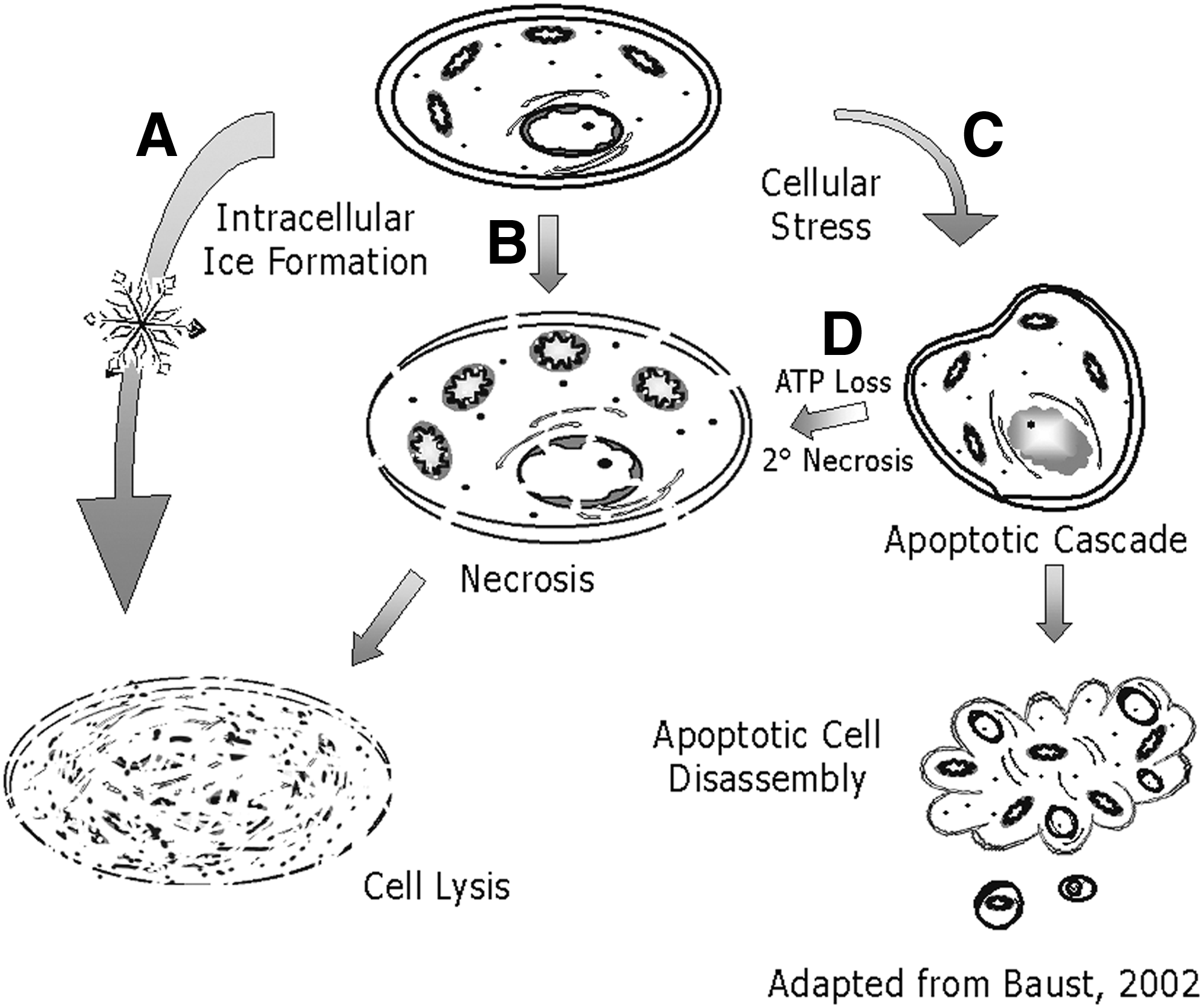
Cell Death Pathways Associated with Cryopreservation Failure. Diagrammatic representation of the various paths of death a cell may undergo as a result of cryopreservation stresses. Physical ice rupture
Stress-Dependent Cell Death Following CP
The involvement of multiple paths of cell death following CP has led to a number of improvements in preservation techniques. Since initial reports of the observation of apoptosis following CP and subsequent reports, which included a wide variety of cell types,48–56 efforts have been made to regulate cell death pathways with new CP solutions.30,41,57 The temporal aspect is significant to understand the impact of molecular-based cell death. The historical assumption that if a cell emerged from the CP process intact, it had “survived” and remained viable. However, it is now recognized that accumulated stresses during the preservation cold chain can initiate molecular-based cell death processes within hours to days following thawing, resulting in delayed cell death. 3 CP-Induced, Delayed-Onset Cell Death (CIDOCD) 32 refers to the phenomenon of delayed peaks in necrosis and apoptosis many hours following thawing. Accordingly, if an assessment of cell death is conducted too early (i.e., before the launch of cell death cascades over 6–48 hours), survival estimates may be biased.4,38,52
In addition, changes to cell function following the preservation process such as diminished functions, loss of surface markers, and molecular differences such as changes in gene expression have been observed.34,58–64 Unstudied but of concern is the potential for the preservation process to select for increased resistance to preservation stresses. The alteration of cell survival signaling pathways is a hallmark of cancer formation and progression. Changes in cell signaling related proteins (i.e., overexpression of Bcl-2 or modification of P53) have been shown experimentally following a freezing exposure.65–68 Hence, an important element of preservation progress will be protocols that limit functional changes in a cell's physiology.
Controlling Molecular-Based Death
We now recognize that a multitude of different pathways and mitigating agents may play a role in determining a cell's fate following CP. Typical CP solutions are prepared by supplementing a cell's basal culture media with serum proteins and dimethylsulfoxide (DMSO). While this formulation provides adequate physical protection and control of intracellular ice formation, it provides only low-level protection against many of the other stresses a cell encounters during the freeze–thaw process. The development of more intracellular-like solutions supplemented with CPAs compared to standard culture media-based CP solution30,41,57,64,69–71 yield improved survival. These advanced CP media are often referred to as complex cryopreservation media (CCM) as they balance ionic needs of cells at low temperatures, buffer pH, provide energy substrates, osmotic stabilization, and protection against free radical damage, among others. Typically, the use of CCM results in improved sample quality post-thaw, including increased viability, functionality, and regrowth, which are usually not evident until hours after thawing. These improvements by CCM have been shown to be a result of decreases in molecular-based cell death during the recovery period.2,30
More recently, advances in CP have been achieved through the incorporation of compounds into the CP media with the aim to control cell death through the targeting of molecular stress response pathways. This strategy is often referred to as Targeted Apoptotic Control (TAC). The use of chemical modulators (i.e., free radical scavengers, protease inhibitors, and ion chelators) has demonstrated the ability to improve CP and hypothermic storage outcomes.28,72–74 For instance, Mathew et al. 29 demonstrated the beneficial effect of the addition of vitamin E, a potent antioxidant, during hypothermic preservation. Other reports described the beneficial effect of EDTA addition and demonstrated implications of apoptotic involvement. 74 In the area of CP, early reports by Baust et al., 33 Yagi et al., 75 and others49,56,76 demonstrated the benefit of caspase inhibition in improving CP outcome. These early reports demonstrated the ability to control cell death and ultimately improve postpreservation outcomes in a number of different cell systems.
Recent studies of targeted molecular control of cell death have focused on apoptosis. Through the use of caspase inhibitors during CP, reports have demonstrated significant improvements in outcome through the blocking of apoptosis activation.33,49,56,75,76 Interestingly, studies have shown that the utilization of caspase inhibitors has a beneficial effect in multiple cell types through a decrease in both apoptosis and necrosis, further strengthening the linkage of the cell death continuum.38,54,77 Other studies show that the poststorage addition of caspase inhibitors has beneficial effects on outcome.56,76 Another area of focus centers on apoptotic control through the management of Rho-associated kinase (ROCK) through the use of ROCK inhibitor.78,79 Specifically, research has shown beneficial effects of ROCK inhibitor on stem cell CP when incorporated during and postpreservation, including increased survival and decreased spontaneous differentiation. 80 Interestingly, the utilization of ROCK inhibitor before single-cell dissociation of pluripotent stem cells, a necessary step for CP, has been shown to block the dissociation-based apoptotic activation in these cells and thus improves preservation outcomes. Importantly, these studies demonstrate that the entire preservation process (the cold chain) is critical for cell quality and survival. To this end, a combinatorial approach of targeting stress pathways both during and after CP may result in improved outcomes for biologics.
Next Steps in Improving CP
A next step in our ability to modulate the post-thaw cell stress response will be at the level of transcriptional and translational events.1,2,4,6–8,81 Typically, improved outcomes result from targeted approaches that act through changes to cellular function at the genomic, proteomic, and epigenetic level. Newer molecular tools such as RNAi, 82 gene manipulation, and epigenetic control may demonstrate the potential for alterations at the level of DNA and RNA as novel approaches for improved CP outcomes. Similarly, the ability to manipulate genes at the level of DNA transcription through gene insertion or deletion has the potential to improve CP outcome. Through these techniques, it may be possible to control levels of both prodeath and prosurvival proteins in an effort to increase survival through a targeted approach.
There is a growing body of scientific literature about epigenetic alterations observed after freezing. Exposure of cells and gametes to DMSO has shown to alter global DNA methylation patterns in mouse embryoid bodies. 6 Preservation also decreases DNA methylation levels in bovine oocytes. 7 Freezing has been associated with DNA fragmentation and DNA methylation in sperm.7–9 Epigenetic changes that persist have serious implications for preserved gametes used in assisted reproductive technologies (ART). There is the potential for ART to result in imprinting-related diseases, and it is not clear the contribution made by CP. Epigenetic alterations are also a concern for gene-edited cells as they may produce instabilities and changes in cell karyotype. Alterations in the epigenome of cells or gametes that have been preserved need further study. Reports now show a connection between thermal tolerance and epigenetic changes, 81 demonstrating the potential for epigenetic manipulations as a modulator to improve CP outcome. Epigenetics could potentially allow for the targeted control of gene expression in a reversible manner that may not alter a cell's DNA sequence. As such, it will be important to understand the nature of the alterations and specific factors that lead to those alterations (exposure to DMSO or hypertonic solutions, the freezing event, etc.), implications for those epigenetic alterations, and strategies to minimize them.
Last, while our understanding of the roles that organelles such as the cell membrane, nucleus, and mitochondria play in the cell death cascade continues to evolve, research has revealed that other organelles may contribute to a cell's fate as well. Specifically, the lysosomes, Golgi apparatus, and endoplasmic reticulum (ER) have been shown to play a role in cellular stress monitoring with the ability to trigger a molecular-based response if irregularities are detected. 83 The ER in particular has become a major area of research in recent years as reports have revealed its role in cellular homeostasis and its subsequent ability to trigger an apoptotic response in the event of profound cellular distress.84–86 Specifically, the mechanism through which the ER conducts these functions is known as the unfolded protein response.87–91 As our understanding of the role and impact these areas have on CP outcome continues to evolve, they too may provide additional targets for modulation to improve post-thaw outcome (survival, function, and genomic stability).
Summary
As cell-based research and development continue to advance, it has become clear that CP protocols have not kept pace with current and emerging needs. While today's standard of practice for CP still focuses primarily on the control of osmotic flux, ice formation, and associated stressors, numerous reports have emerged demonstrating that the combined targeting of physical, biochemical, and molecular responses to the freez–thaw process can greatly improve cellular outcome. The impact of this combined approach not only extends well beyond influencing cell death but also has a long-term impact on cellular functionality. The results of these practices are, of course, only realized on sample thawing. Given the complexity of events that occur during the seemingly simple and routine process of freezing, it is important to recognize and understand the impact of sample preparation, cooling, storage temperature, storage time, and warming on overall sample quality. The ability to successfully cryopreserve samples depends on the integration of the molecular strategy to obtain the highest quality cell product post-thaw. This strategic shift includes (1) structural methodologies that are reasonably effective (optimized) in preventing ice-related damage, (2) the use of CCM-based CP media, and (3) integration of modulation strategies to reduce CIDOCD. While this new approach is well supported, there remains a continuing need to decipher the cell's responsiveness to the stress events associated with CP to overcome the significant cell death after thawing.2,32 Particularly important is the integrated study of cell response to hypothermic and subfreezing temperatures as they are experienced by cells during both freezing and thawing processes associated with CP. Through a more complete understanding of low-temperature stress response and how associated changes alter a cell's typical, normothermic baseline of function, significant improvements in CP strategies can be achieved in turn yielding improved biologics for downstream utilization.
Footnotes
Author Disclosure Statement
J.M.B., R.V.B. and K.K.S. are employees of CPSI Biotech. No other conflicting financial interests exists.